
Biohydrogenation Pathway of α-Linolenic Acid in Rumen of Dairy Cow In Vitro

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Animal Management
2.2. C18 Fatty Acid Analysis
2.3. Statistical Analysis
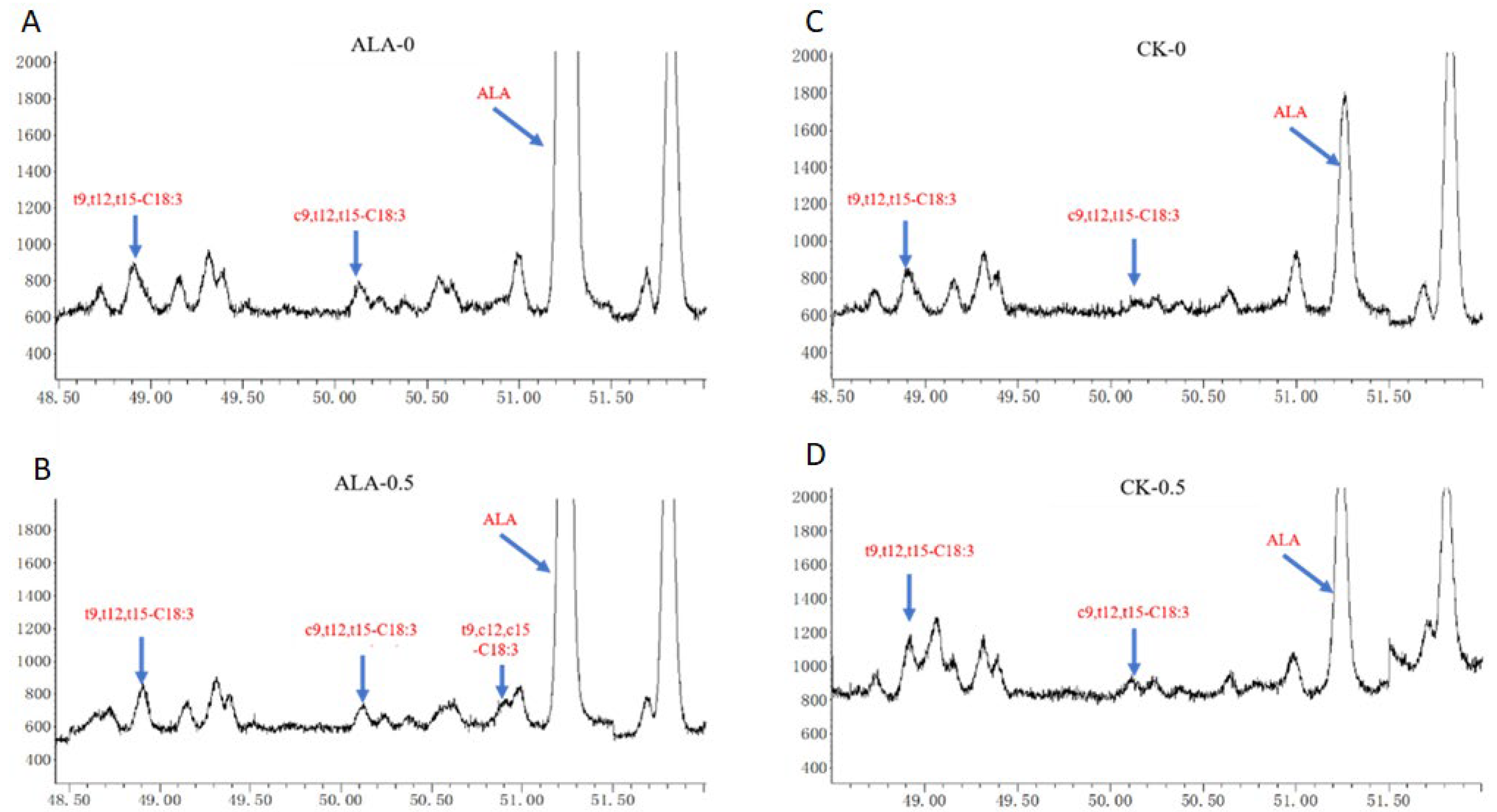
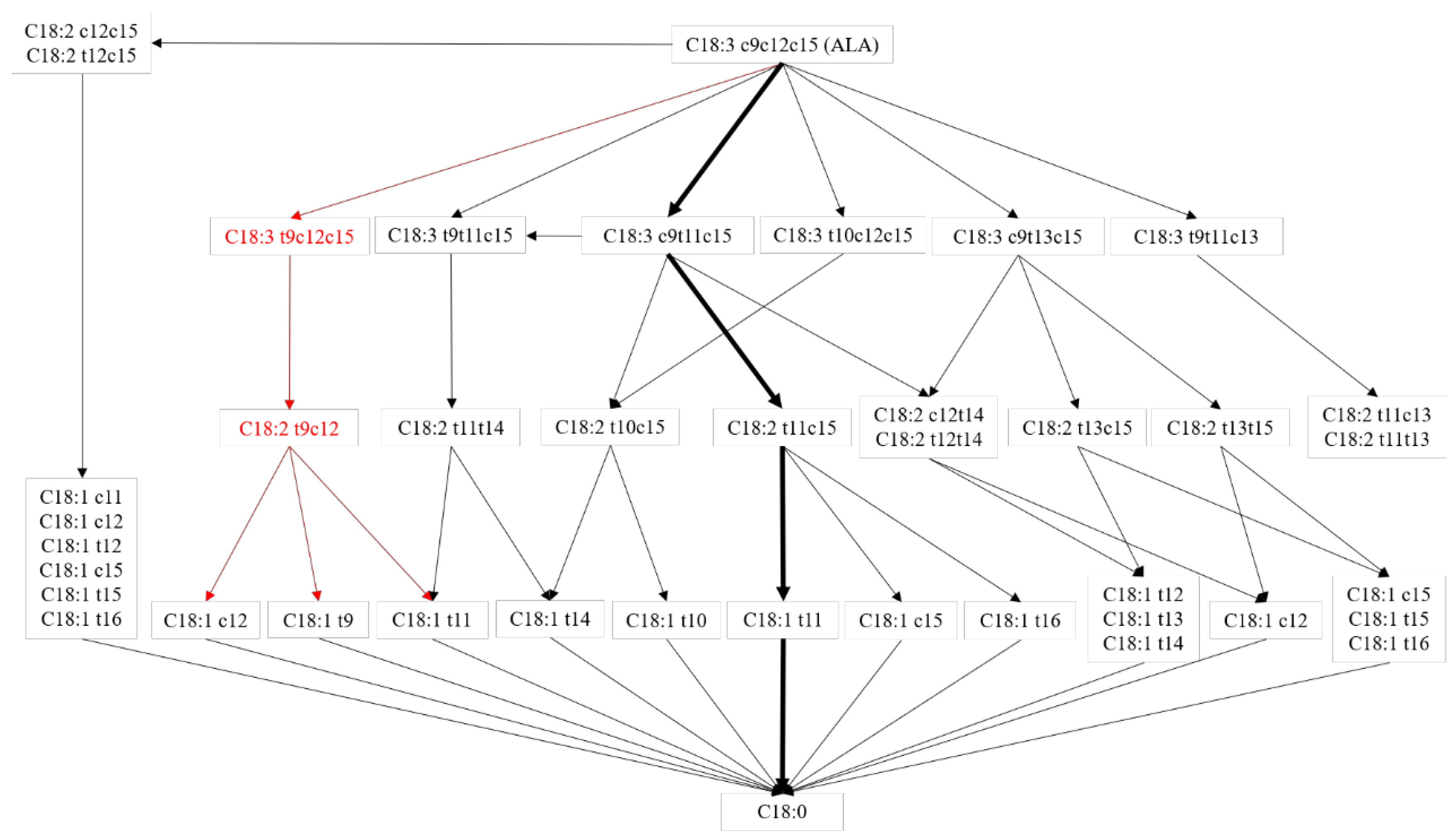
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, G.X.; Zhang, Y.D.; Xu, Q.B.; Zheng, N.; Zhao, S.G.; Liu, K.Z.; Qu, X.Y.; Yu, J.; Wang, J.Q. DHA content in milk and biohydrogenation pathway in rumen: A review. PeerJ 2020, 8, e10230. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, A.; Bernard, L.; Meynadier, A.; Malpuech-Brugère, C. Production of trans and conjugated fatty acids in dairy ruminants and their putative effects on human health: A review. Biochimie 2017, 141, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, M.; Ramos-Morales, E.; Wallace, R.J. The role of microbes in rumen lipolysis and biohydrogenation and their manipulation. Animal 2010, 4, 1008–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewanckele, L.; Toral, P.G.; Vlaeminck, B.; Fievez, V. Invited review: Role of rumen biohydrogenation intermediates and rumen microbes in diet-induced milk fat depression: An update. J. Dairy Sci. 2020, 103, 7655–7681. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Yang, Y.; Nie, S.; Xie, M.; Chen, F.; Luo, P.G. Formation of trans fatty acids during the frying of chicken fillet in corn oil. Int. J. Food Sci. Nutr. 2014, 65, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.Q.; Zhang, Y.D.; Wang, F.E.; Zheng, N.; Wang, J.Q. Simultaneous determination of C18 fatty acids in milk by GC-MS. Separations 2021, 8, 118. [Google Scholar] [CrossRef]
- Menke, K.H.; Raab, L.; Salewski, A.; Steingass, H.; Fritz, D.; Schneider, W. The estimation of the digestibility and metabolizable energy content of ruminant feeding stuffs from the gas production when they are incubated with rumen liquor in vitro. J. Agric. Sci. 1979, 93, 217. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.X.; Guo, L.Y.; Chang, X.F.; Liu, K.Z.; Tang, W.H.; Zheng, N.; Zhao, S.G.; Zhang, Y.D.; Wang, J.Q. Effect of whole or ground flaxseed supplementation on fatty acid profile, fermentation, and bacterial composition in rumen of dairy cows. Front. Microbiol. 2021, 12, 760528. [Google Scholar] [CrossRef] [PubMed]
- Baldin, M.; Rico, D.E.; Green, M.H.; Harvatine, K.J. Technical note: An in vivo method to determine kinetics of unsaturated fatty acid biohydrogenation in the rumen. J. Dairy Sci. 2018, 101, 5669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pi, Y.; Ma, L.; Pierce, K.M.; Wang, H.R.; Xu, J.C.; Bu, D.P. Rubber seed oil and flaxseed oil supplementation alter digestion, ruminal fermentation and rumen fatty acid profile of dairy cows. Animal 2019, 13, 2811–2820. [Google Scholar] [CrossRef] [PubMed]
- Costa, D.F.A.; Quigley, S.P.; Isherwood, P.; McLennan, S.R.; Sun, X.Q.; Gibbs, S.J.; Poppi, D.P. The inclusion of low quantities of lipids in the diet of ruminants fed low quality forages has little effect on rumen function. Anim. Feed Sci. Tech. 2017, 234, 20–28. [Google Scholar] [CrossRef]
- Collomb, M.; Sollberger, H.; Bütikofer, U.; Sieber, R.; Stoll, W.; Schaeren, W. Impact of a basal diet of hay and fodder beet supplemented with rapeseed, linseed and sunflower seed on the fatty acid composition of milk fat. Int. Dairy J. 2004, 14, 549–559. [Google Scholar] [CrossRef]
- Harfoot, C.G. Lipid metabolism in the rumen. Prog Lipid Res. 1978, 17, 21–54. [Google Scholar] [CrossRef]
- Kishino, S.; Ogawa, J.; Yokozeki, K.; Shimizu, S. Metabolic diversity in biohydrogenation of polyunsaturated fatty acids by lactic acid bacteria involving conjugated fatty acid production. Appl. Microbiol. Biotechnol. 2009, 84, 87–97. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, G.; Guo, L.; Chen, M.; Wu, X.; Tang, W.; Zheng, N.; Zhao, S.; Zhang, Y.; Wang, J. Biohydrogenation Pathway of α-Linolenic Acid in Rumen of Dairy Cow In Vitro. Animals 2022, 12, 502. https://doi.org/10.3390/ani12040502
Huang G, Guo L, Chen M, Wu X, Tang W, Zheng N, Zhao S, Zhang Y, Wang J. Biohydrogenation Pathway of α-Linolenic Acid in Rumen of Dairy Cow In Vitro. Animals. 2022; 12(4):502. https://doi.org/10.3390/ani12040502
Chicago/Turabian StyleHuang, Guoxin, Liya Guo, Meiqing Chen, Xufang Wu, Wenhao Tang, Nan Zheng, Shengguo Zhao, Yangdong Zhang, and Jiaqi Wang. 2022. "Biohydrogenation Pathway of α-Linolenic Acid in Rumen of Dairy Cow In Vitro" Animals 12, no. 4: 502. https://doi.org/10.3390/ani12040502