1. Introduction
Raw starch, the main energy source of monogastric animals, widely exists in energy feed (corn, wheat, barely, etc.) and can be abruptly stressful in the digestion process of weaning piglets [
1,
2,
3] because weaning stress has been associated with impaired intestinal digestive function [
4,
5]. Extrusion has been the most common pre-digestion process for raw starch in feed production [
6]. The highly ordered structure of starch is destroyed and the gelatinization degree is increased after extrusion [
7], which increases the efficiency of enzymatic starch hydrolysis by α-amylase in the small intestine, and further improves the digestibility of starch in weaning piglets [
8].
Starch digestion occurs primarily in the piglets’ small intestine, where the pancreatic α-amylase hydrolyzes starch to maltose and larger oligosaccharides, which are further degraded to glucose by brush border membrane oligosaccharidases before absorption [
9,
10]. Previous research has shown that amylase supplementation in vivo could complement the deficiency of endogenous amylase, improving the digestibility of starch in cereal and enhancing the subsequent growth performance of weaning piglets [
11,
12,
13].
Although the various functions of extrusion processes in vitro or amylase supplementation in vivo have received extensive attention in weaning piglets [
1,
2,
3,
4,
5,
6,
7,
8,
9,
10,
11,
12,
13], there are few studies on the application of amylase in vitro during the extrusion of raw corn starch due to the high temperature of the process. However, the optimal reaction temperature of thermostable α-amylase is as high as 105 °C, which implicates the feasibility of in vitro degradation of corn starch via enzymatic hydrolysis during the extrusion process [
6,
7,
8]. Therefore, the present experiment was designed to investigate the effects of extruded corn with added thermostable α-amylase under different moisture conditions on the growth performance, digestive capability, and intestinal microbiome of weaning pigs, which provided new insights into the applications of extruded corn with added enzymes for the pig industry.
2. Materials and Methods
All animal procedures associated with this study were approved by the Animal Care and Use committee, Sichuan Agricultural University (Ethic Approval Code: SICAUAC202106-1; Chengdu, China). The primary experiment was conducted at the Animal Experiment Center of Tongwei Co., Ltd., Chengdu, China.
The conditioner (MUTZ1610) and extruder (EXT155G) were provided by Jiangsu Speed Machinery Co., Ltd., Changzhou, China and Beijing Modern Yanggong Technology Development Co., Ltd., Beijing, China, respectively. In addition, the α-amylase (provided per gram of α-amylase units) was provided by Sainuo biological technology Co., Ltd., Baiyin, China.
2.1. Experimental Design and Animal Management
A total of 48 twenty-four-d-old pigs (Duroc × Landrace × Yorkshire, weaned at 22 ± 1 d and fed the basal diet for a 3-d adaptation period) with a body weight (BW) of 6.76 ± 0.15 kg were used in a 14-d experiment. At the beginning of the experiment, pigs were assigned to four treatments with six replicates (one male and one female per pen) on the basis of their initial BW and sex in a randomized complete block design. The four dietary treatment groups included: (1) NL (adding 7.5% water before corn extrusion); (2) NH (adding 15.0% water before corn extrusion); (3) PL (adding 7.5% water and 4 kg/t α-amylase before corn extrusion); and (4) PH (adding 15% water and 4 kg/t α-amylase before corn extrusion).
The corn meal was pretreated before conditioning and extruded according to the different treatments. Briefly, NL added water with 7.5% the mass of corn meal, NH added water with 15% the mass of corn meal, PL added water with 7.5% the mass of corn meal and supplemented with 4 kg/t α-amylase, and PH added water with 15% the mass of corn meal and supplemented with 4 kg/t α-amylase; following this, the pre-treated corn meal was prepared under the same conditioning at 90 °C, extruding at 105 °C, cooling at room temperature, and screening at 2.0 mm. The basal diet (
Table 1) was formulated based on the NRC (2012) recommendations [
14].
All weaning pigs were fed the diets three times per day at 08:00, 14:00, and 20:00 h, provided with water ad libitum, and housed in a temperature- (27 ± 1 °C) and relative-humidity-controlled (55% to 65%) room. All weaning pigs were weighed at the beginning and the end of the experiment after 12 h of fasting, and the average daily gain (ADG), average daily feed intake (ADFI), and feed conversion ratio (F/G) were calculated with feed intake (FI) per pen daily throughout the experiment.
2.2. Sampling
The dry basal diet samples and fresh fecal samples collected with a partial collection method per pen after immediate defecation from days 11 to 14 during the trial were collected for chemical analysis. Next, 10 mL of a 10% dilute sulfuric acid solution was added to each 100 g of plastic-bagged fecal sample for corrosion prevention and fixation of excreta nitrogen, which was stored at 20 °C. On day 14, one average BW weaning pig per pen was selected and euthanized with sodium pentobarbital according to a previous study [
15], and gut sections were collected from the unfolded abdomen immediately. The complete small intestine was removed, cleaned, and cut into the duodenum, jejunum, and ileum in line with our previous study [
16]. Subsequently, approximately 1-cm, 2-cm, and 3-cm segments of proximal duodenum, jejunum, and ileum, respectively, were obtained using the following three steps: immediate isolation, wash with 0.9% physiological saline, and preservation with 10% formaldehyde-phosphate buffer for histological analysis. A 10-cm section of proximal jejunum was gently emptied with gloved hands, carefully flushed with saline, softly placed on an icy surface, and the jejunal mucosa was gently scraped and stored at −80 °C for RNA extraction and activities of digestion-related and absorption-related enzymes. Additionally, for determining gut microbiome and metabolites, the digesta from the middle cecum (10 cm) and middle colon (10 cm) were collected and stored at −80 °C.
2.3. Diarrhea
During the experiment from d 1 to d 14, the same person observed and recorded the occurrence of diarrhea on two piglets per pen every morning and evening, and calculated an accumulative diarrhea score per treatment and day to calculate the diarrhea rate according to a previous study [
17]. The diarrhea rate was calculated as follows: diarrhea rate (%) = D/(T × 14 d) × 100, in which D = total number of diarrheal pigs per pen and T = number of pigs per pen.
2.4. Moisture and Maltooligosaccharides Content in Extruded Corn
The extruded enzyme-added corn samples were analyzed for moisture content (method 930.15; AOAC, 1995) [
18]. Moreover, the extruded enzyme-added corn was analyzed using high-performance anion exchange chromatography with pulsed amperometric detection (HPEAC-PAD, Thermo Scientific, Waltham, MA, USA) to detect the maltooligosaccharides content [Glucose (G1), maltose (G2), and maltotetraose through maltoheptaose (G4–G7)] according to the previous study by Ding et al. (2020) [
19].
2.5. Apparent Digestibility of Nutrients and Enzyme Activities
The fresh feces from days 11 to 14 of each pen were mixed thoroughly with cleaned plastic bags, dried at 65 °C for 72 h, and ground with a small-scale mill to pass through a standard 40-mesh screen. Chromium trioxide (Cr
2O
3), as a digestibility indicator, was applied to measure the apparent total tract digestibility (ATTD) of nutrients, with an adaptation period of 4 days. The Cr
2O
3 in the diet and fecal samples was analyzed with a Chinese National Standard Method (GB/T 13088-2006) [
20]. After the Cr
2O
3 analysis, all feed and fecal samples were analyzed for dry matter (method 930.15; AOAC, 1995), ash (method 923.03; AOAC, 1995), crude fat (method 920.39; AOAC, 1995), crude protein (method 990.03; AOAC, 1995), calcium (method 927.02; AOAC, 1995), and phosphorus (method 995.11; AOAC, 1995) [
18]. A specific adiabatic oxygen bomb calorimetry (Parr Instrument Co., Moline, IL, USA) was used for the determination of gross energy. The ATTD was calculated as follows: ATTD (%) = {1 − [(A
1 × F
2)/(A
2 × F
1)]} × 100, in which A
1 = dietary Cr
2O
3 content (% DM), A
2 = fecal Cr
2O
3 content (% DM), F
1 = dietary nutrient content (% DM), and F
2 = fecal nutrient content (% DM).
The jejunal sample was per-treated before measuring the activity of digestion and absorption enzymes. Briefly, the jejunal mucosa sample was weighed with pre-cooled saline according to a mass volume ratio of 1:9 (g/mL), and the mixture was homogenized with an utrasonic homogenizer at 4000× g for 10 min at 4 °C to collect the supernatant solution. Subsequently, supernatant protein concentration, as well as the activities of trypsin, lipase, α-amylase, β-amylase, lactase, maltase, sucrase, γ-glutamyl transferase, Na+-K+-ATPase, alkaline phosphatase, and creatine kinase were analyzed using commercial kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) according to the manufacturer’s instructions.
2.6. Intestinal Morphology
The jejunum morphology was measured as described previously [
21]. Briefly, the jejunum preserved in the fixed solution was embedded in paraffin, made into 5-μm slices, flattened with warm water, and baked at 60 °C for 2 h, followed by staining with hematoxylin and eosin, dehydration with alcohol, and sealing with resin glue. An Olympus CK 40 microscope (Olympus Optical Company, Shenzhen, China) was applied to measure villus height and crypt depth. A minimum of 10 complete villi height, and the corresponding depth of 10 crypts, from each jejunal segment were measured.
2.7. Real-Time Quantitative PCR
Jejunum mucosal samples (approximately 0.1 g) were homogenized in 1 mL RNAiso Plus reagent (TaKaRa, Dalian, China), and total RNA was extracted and stored at −80 °C according to the instructions. The concentration and quality of total RNA were verified with a nucleic acid protein instrument that measured the corresponding optical density (OD)260:OD280 ratio, a qualified RNA sample of which should range from 1.8 to 2.0. Meanwhile, the synthesis of cDNA per sample was obtained using a PrimeScript™ reverse transcription reagent kit (TaKaRa, Dalian, China) following the manufacturer’s instructions.
Specific primers for the Sodium-glucose co transporter 1 (
SGLT1), Glucose transporter 2 (
GLUT2), Peptide transporter 1 (
PePT1), Zonula occludens 1 (
ZO-1), Occludin (
OCLN), Claudin 1 (
CLDN-1), and Mucin 1 (
MUC1) were designed and purchased from Invitrogen (Shanghai, China), which are listed in
Table 2. The real-time PCR reactions were performed on a CFX96 Real-Time PCR Detection System (Bio-Rad Laboratories, Inc., Hercules, CA, USA), using SYBR Green PCR reagents (TaKaRa, Dalian, China). After each real-time quantitative PCR test, a melting curve analysis was generated to check and verify the specificity and purity of all PCR products, and β-actin was chosen as the reference gene to normalize cDNA loading. After verifying that the primers amplified with an efficiency of approximately 100%, the results were analyzed using the 2
−ΔΔCt method [
22].
2.8. Gut Microbiome
The QIAamp DNA stool Mini Kit (Qiagen GmbH, Hilden, Germany) was used for the extraction of total genomic DNA from fresh fecal samples. The concentration (purity) and integrity of the extracted genomic DNA were measured using a NanoDrop ND-1000 Spectrophotometer (Nano-Drop Technologies Inc., Wilmington, DE, USA) and electrophoresis on 1% (
w/v) agarose gels, respectively. Extracted fecal DNA samples were sent to Majorbio Bio-pharm Technology Co., Ltd. (Shanghai, China) to perform amplicon pyrosequencing on the Illumina MiSeq platforms (Illumina Inc., San Diego, CA, USA). The V3–V4 hypervariable region of the 16S rRNA gene was amplified by PCR with primers 338F (5′-ACTCCTACGGGAGGCAGCAG-3′) and 806R (5′-GGACTACHVGGTWTCTAAT-3′). The relative abundance of each operational taxonomic units (OTU) was examined at different taxonomic levels. Diversity within communities (Alpha diversity) calculations and taxonomic community assessments were performed by Mothur 1.30.2 (University of Michigan, Ann Arbor, MI, USA) and Qiime 1.9.1 (National Microbiology Data Center, Beijing, China). Principal coordinates analysis (PCoA) plots were produced using unweighted UniFrac metrics [
23]. The linear discriminant analysis (LDA) effect size (LEfSe) method was performed to elucidate the difference among treatments.
2.9. Microbial Metabolites Analysis
Approximately 0.7 g of digesta samples from the cecum and colon were used to determine the concentration of volatile fatty acids (VFA) [
15]. Briefly, the supernatants of digesta samples were centrifuged at 500×
g for 10 min after adding 1:1 distilled water, then 2 mL of the supernatant was transferred to a sterile tube and centrifuged at 12,000×
g for 10 min, after which 1 mL of the supernatant was transferred to a new sterile tube to which 0.2 mL of 25% metaphosphoric acid was added. This was left at room temperature for 30 min and then centrifuged at 12,000×
g for 10 min. Next, 500 μL of the supernatant was transferred to another sterile tube, to which 500 μL of methanol was added and the mixture was centrifuged at 12,000×
g for 10 min, with the resulting supernatant being stored at −20 °C until testing. A gas chromatographic system (VARIAN CP-3800, Varian, Palo Alto, CA, USA) was applied for the separation and quantification of the volatile fatty acids (acetic acid, propionic acid and butyric acid, isobutyric acid, valeric acid, and isovaleric acid).
2.10. Statistical Analysis
Moisture and maltooligosaccharides content in extruded corn data (n = 2) were analyzed by a t-test using the statistical program of SAS (SAS Inst. Inc., Cary, NC, USA). Moreover, the other data (n = 6) were analyzed by a two-way ANOVA using the Generalized Linear Models procedure of SAS 9.0 software (SAS, Raleigh, NC, USA), which included the effects of water content, enzyme addition, and the interaction between water content and enzyme addition. The results were presented as means and SEM. Duncan’s multiple-range test was used to determine statistical differences among treatment. For significance determination, the data results were significant with p < 0.05, and 0.05 < p < 0.1 was considered as a tendency.
4. Discussion
Previous studies have shown that the moisture content of corn meal during conditioning could affect the maltooligosaccharides content in extruded corn [
24,
25]. The present study found that enzyme treatment increased the content of small molecular sugars, especially the oligosaccharides composed of 6–7 glucose molecules, which was consistent with the previous study [
26,
27] that reported that the addition of amylase during the conditioning and extruding process could increase the content of maltooligosaccharides.
The moisture content of raw materials, as a medium of α-amylase action, has a synergistic effect on enzymatic hydrolysis, which plays a key role in the extrusion with added enzymes [
28]. A low moisture content of raw materials could be detrimental to the enzymatic reaction, and an excessive moisture content of raw materials made against the extrusion effect and subsequent processing [
29,
30]. In the present study, it was found that enzyme treatment significantly increased animal feed intake and further improved the daily weight gain of animals. However, the increase of moisture content from 7.5% to 15% had no significant effect on animal growth performance, and there was no interaction effect between the moisture and enzyme treatments.
During the post-weaning period, piglets often suffer from social (maternal and littermate separation), environmental (transportation), and dietary (abrupt change in diets) stressors [
31,
32]. Weaning stress was associated with destroyed intestinal morphology [
33], impaired intestinal barrier, lower activities of epithelial brush border enzyme, decreased digestibility of nutrients, and a subsequently reduced performance [
34,
35,
36]. The present study found that the enzyme treatment improved the intestinal morphology of weaning piglets. We supposed that positive results observed in the study herein may be owing to the following reasons: the low feed intake during the period of weaning resulted in a decrease in villus height and an increase in crypt depth [
37]; and extrusions with added enzymes significantly increased the feed intakes of weaning pigs and improved the digestibility of corn starch in the first two weeks, as it was reported that the high feed intakes after weaning could improve the morphology of the small intestine in weaning piglets [
33].
In the present study, it was found that enzyme treatment significantly improved the ATTD of nutrients in weaning piglets, while moisture content had no significant effect. This was consistent with growth performance, indicating that the pre-digestion effect of α-amylase on corn starch during extrusion was the main reason to improve the nutrients’ digestibility. The enzyme activity in the digestive tract was considered to be a key factor affecting intestinal health and nutrient digestibility [
38]. In addition, many previous studies have shown that the exogenous addition of amylase or improvement of starch gelatinization can promote the activity of digestive enzymes in weaning piglets [
39,
40]. The present study found that the enzymatic hydrolysis of starch in the process of extrusion in vitro can significantly improve the jejunal trypsin and α-amylase, lipase, sucrase, maltase, γ-glutamine transferase, and alkaline phosphatase activities. Recent studies showed that
SGLT1,
GLUT2, and
PePT1 were extensively located at the intestinal mucosa, where they serve as the main transporters, respectively, for glucose, glucose, and amino acids [
41]. We found that enzyme treatment significantly increased the relative mRNA expressions of the
SGLT-1 and
GLUT-2 genes in the jejunum of weaning piglets, which was consistent with the results reported by other studies, where starch that was hydrolyzed in vitro by α-amylase could significantly increase the mRNA expressions of
SGLT-1 and
GLUT-2 genes in IPEC-J2 cell [
42].
The integrity of the intestinal barrier played an important role in the proper functioning of the epithelial cells, which could reduce the increment of pathogenic bacteria [
43]. However, weaning stress contributed to impaired intestinal barrier function and enhanced intestinal permeability [
36]. The intestinal tight junction protein family (
CLDN,
OCLN,
ZO, etc.) determined endothelial and epithelial paracellular barrier functions, which prevented the paracellular diffusion of intestinal bacteria and other antigens across the epithelium [
44,
45]. The present study found that the enzyme treatment significantly increased the relative mRNA expressions of the
ZO-1 and
OCLN genes in the jejunum of weaning piglets, a possible reason for which might owned to the improvement of small molecular oligosaccharides by α-amylase action during the conditioning and extruding process, which may contain some oligosaccharides with intestinal regulation function, such as Isomaltooligosaccharides. It was found that the addition of Isomaltooligosaccharide could increase the relative mRNA expression of the
OCLN gene in intestinal epithelial cells, increasing the intestinal barrier function of weaning piglets [
46].
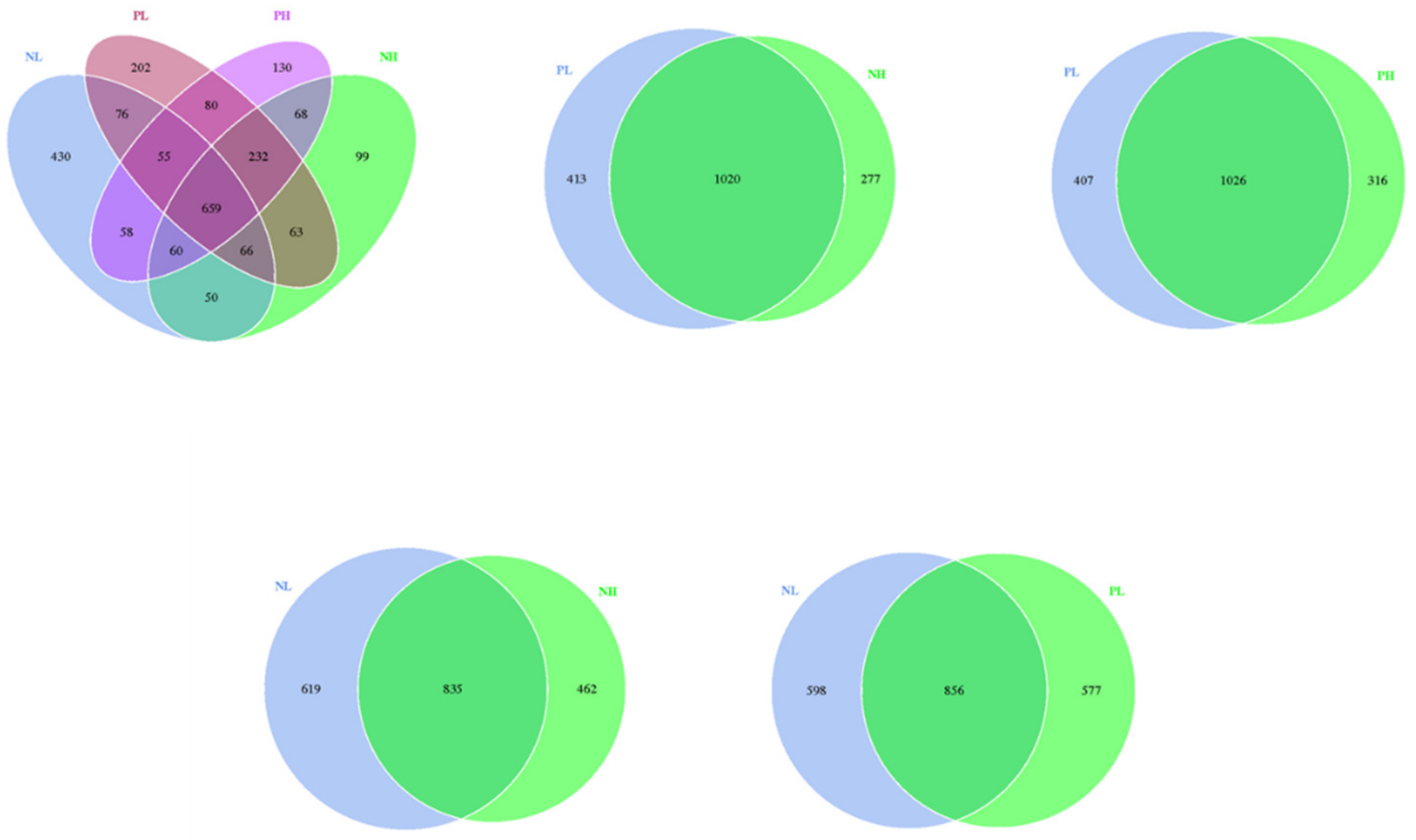
The intestine microorganisms are associated with digestive and absorptive functions, as well as the subsequent gut health [
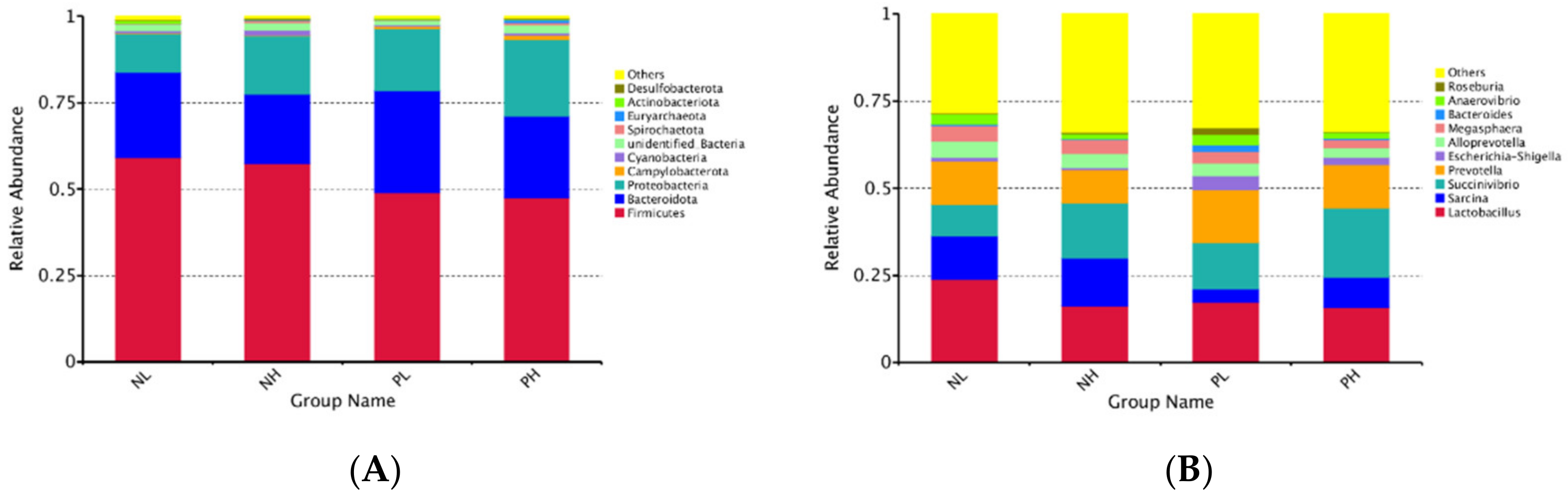
47]. In our study, 16S rRNA sequencing was used to investigate the gut microbiome responses to moisture contents (adding 7.5 vs. 15% water) and enzyme additions (adding 4 kg/t α-amylase or not) to extruded corn in weaning piglets. Our results showed that moisture content and enzyme treatment reduced the number of OTUs of the fecal microbiome, but there was no significant difference in the results of the alpha diversity analysis of the fecal microbiome. At the phylum level, the abundance of Firmicutes tended to decrease in the enzyme treatment group, suggesting that the enzyme treatment may reduce the polysaccharide fermentation of hindgut flora, and further reducing the production of volatile fatty acids (VFAs).
In the present study, piglets fed NL diets increased
Aeriscardovia and
Christensenlace-r-7-group. The previous study found that the
Christensenlace-r-7-group was significantly negatively correlated with metabolic diseases such as body index and inflammation [
48].
Aeriscardovia could synthesize a variety of enzymes, amino acids, and vitamins by itself and metabolize acetic acid, lactic acid, and so on [
49]. The dominant bacteria in the NH group were
Holdemanella and
Desulfovibrio.
Holdemanella was a harmful bacterium associated with intestinal inflammation [
50], and studies have shown that
Desulfovibrio belong to desulfobacterate, and its role might be related to the occurrence of some inflammatory intestinal diseases and colon cancer [
51]. The dominant bacteria in the PL group were
Staphylococcus and
Cag-873.
Staphylococcus is a gram-positive cocci, which might have a negative effect on intestinal health [
52].
Cag-873 was a genus of Bacteroidetes, which was related to hindgut energy homeostasis and had the ability to degrade cellulose, hemicellulose, and lignin [
53]. In addition, the dominant bacteria in the PH group were
Mitsuokella and
Romboutsia, where
Mitsuokella mediates the positive regulation of cell-growth-related genes through the metabolism of acetic acid, and may inhibit the growth of intestinal
Salmonella [
54].
The change of gut microbiome composition contributes to the variation in microbial metabolites. The VFAs are not only positive in providing energy for intestinal epithelial cells, but also promote the formation of cells, improving intestinal morphology [
55]. Among VFAs, acetic acid, propionic acid, and butyric acid are the most important VFAs, and the change of their composition and proportion can, in turn, affect the change of the gut microbiome. Butyric acid was the energy substrate of colonic epithelial cells and can inhibit colonic cell apoptosis [
56]. Acetic acid had the highest concentration among VFAs, accounting for about 60%, followed by propionic acid and butyric acid; in addition, acetic acid is a potential precursor of cholesterol and fatty acids, which can be used as a substrate for the de novo synthesis of fatty acids [
57]. While propionic acid could participate in gluconeogenesis, and valeric acid and isobutyric acid are the products of bacterial fermentation proteins, marking the catabolism of intestinal proteins [
58]. Meanwhile, isobutyric acid could enhance intestinal permeability, which may have a negative impact on intestinal health [
59]. The present study found that the enzyme treatment significantly reduced the contents of isobutyric acid in the cecal digesta, as well as acetic acid and isobutyric acid in the colonic digesta of weaning piglets, and tended to reduce the total VFAs in the colon digesta; a possible reason these results might be related to the higher digestibility of starch and protein, subsequently lowering the fermentation substrate of hindgut flora when supplemented with α-amylase.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}