Simple Summary
Avian pathogenic Escherichia coli (APEC) is the pathogen responsible for colibacillosis in poultry. Transcriptional regulator ygeK has been shown to decrease APEC’s flagellar formation ability, bacterial motility ability, serum sensitivity, and adhesion ability. However, we did not study the effects of ygeK on biofilm formation and environmental stress resistance in APEC. In this study, we investigated ygeK in APEC biofilm formation and bacterial resistance to different environmental stresses. We also analyzed the multi-level regulation of ygeK in APEC and investigated associations between differentially expressed proteins and key ygeK targets. This work provides a basis for further analysis of APEC pathogenesis mechanisms.
Abstract
Avian pathogenic Escherichia coli (APEC) is one of the most common pathogens in poultry and a potential gene source of human extraintestinal pathogenic E. coli (ExPEC), leading to serious economic losses in the poultry industry and public health concerns. Exploring the pathogenic mechanisms underpinning APEC and the identification of new targets for disease prevention and treatment are warranted. YgeK is a transcriptional regulator in APEC and is localized to the type III secretion system 2 of E. coli. In our previous work, the transcription factor ygeK significantly affected APEC flagella formation, bacterial motility, serum sensitivity, adhesion, and virulence. To further explore ygeK functions, we evaluated its influence on APEC biofilm formation and resistance to environmental stress. Our results showed that ygeK inactivation decreased biofilm formation and reduced bacterial resistance to environmental stresses, including acid and oxidative stress. In addition, the multi-level regulation of ygeK in APEC was analyzed using proteomics, and associations between differentially expressed proteins and the key targets of ygeK were investigated. Overall, we identified ygeK’s new function in APEC. These have led us to better understand the transcriptional regulatory ygeK and provide new clues about the pathogenicity of APEC.
1. Introduction
Avian pathogenic Escherichia coli (APEC) is a type of extraintestinal pathogenic E. coli (ExPEC) that causes colibacillosis in poultry, leading to significant economic losses to the poultry industry [1]. Due to its diverse serotypes and complex virulence factors, no effective vaccines are available against APEC [2]. In addition, APEC is becoming increasingly resistant to widespread antibiotic use; even the polymyxin resistance gene mcr-1 has begun to appear in clinical APEC isolates, with life-threatening risks on a global scale [3,4,5]. Gene cluster comparisons of animal and human mcr-resistant strains suggest that bacteria carrying mcr genes are potentially zoonotic. Therefore, exploring pathogenic mechanisms in APEC is important and will promote the development of effective vaccines or new drug targets to control this infection.
Biofilms are multicellular bacterial aggregates bound by a polymeric matrix consisting of complex mixtures of extracellular polysaccharides, proteins, and DNA [6,7]. These structures allow bacteria to tolerate a variety of environmental pressures [7,8]. Many bacterial species form complex and diverse biofilms. Researchers have concentrated on exploring the formation mechanism of biofilms and looking for inhibitory strategies for biofilm formation [9,10]. In APEC, several transcription regulators regulate bacterial biofilm formation, e.g., deletion of the two-component system basSR inhibits in vitro APEC biofilm formation and decreases bacterial virulence and colonization in vivo [11]. PhoP is a transcriptional regulator in the two-component phoP/phoQ regulatory system; it up-regulates APEC biofilm formation and is associated with changes in bacterial drug resistance and cell-membrane-related properties [12]. McbR increases APEC biofilm formation by up-regulating transcription of the biofilm-associated genes, bcsA, fliC, wcaF, and fimA, and also affects oxidative responses by regulating transcription of the yciGFE operon [13]. Oxidative stress is one of the stress environments that bacteria encounter when they infect the host. In APEC, some genes have been reported to be associated with environmental stress (such as acid, alkali, and oxidative stress). Understanding bacterial resistance to environmental stress will help further explore bacterial survival mechanisms.
YgeK was identified as a regulator of gene expression in enterohemorrhagic E. coli [14,15]. We previously showed that ygeK inactivation in AE81 reduced several bacterial functions, including flagella formation, motility, bactericidal activity, and adhesion [16]. Studies reported that flagella are more than a locomotive organelle for E. coli, they are also critical for biofilm formation [17]. However, we did not study the effects of ygeK on biofilm formation in APEC. In this study, we investigated ygeK in APEC biofilm formation and bacterial resistance to different environmental stresses. We also analyzed the multi-level regulation of ygeK in APEC and investigated associations between differentially expressed proteins and key ygeK targets. This work provides a basis for further analysis of APEC pathogenesis mechanisms.
2. Materials and Methods
2.1. Wild-Type, Mutant, and Complement Strains
The wild-type strain, AE81, was isolated from the lung of a dead, septicemic chicken with suspected colibacillosis in Anhui, China [18]. The mutant, AE81ΔygeK, and complemented strain, AE81ΔygeK-pCmygeK, were constructed in our previous study [16]. Where necessary, we supplemented chloramphenicol (30 µg/mL) to lysogeny broth solid medium.
2.2. Crystal Violet (CV) Staining of Biofilms
AE81, AE81ΔygeK, and AE81ΔygeK-pCmygeK cultures were diluted to OD600 = 0.03 and incubated at 28 °C for 72 h. Stationary phase cultures were washed three times in sterile phosphate buffer saline (PBS) and air-dried. Then, 100% methanol was added to immobilize biofilm-forming cells for 5 min and a 0.1% (w/v) CV solution was added to stain cells for 15 min. Excess stain was rinsed away with distilled water and cells were re-air-dried. The remaining CV in the growth tube was dissolved in 33% glacial acetic acid (Sanggong, Shanghai, China) and the solution read at 492 nm on a Micro Elisa microplate reader (Thermo Scientific, Pittsburgh, PA, USA) [19]. Experiments were performed three times.
2.3. Scanning Electron Microscopy (SEM)
We incubated 1 mL bacterial suspensions (OD600 = 1.0) on sterile glass coverslips (diameter = 10 mm) in 12-well plates at 37 °C for 24 h. The next day, the coverslip was washed three times in PBS and glutaraldehyde (2.5%) was added to the wells. After incubation at 4 °C for 10 h, wells were washed three times in PBS and treated with 1% osmic acid for 5 h. Cells were then dehydrated in 30, 50, 60, 70, 80, 90 and 100% ethyl alcohol. Cells were then observed using field SEM (Hitachi S-4800, Chiyotaku, Japan).
2.4. Hydrogen Peroxide (H2O2) Stress Assays
Overnight AE81, AE81ΔygeK, and AE81ΔygeK-pCmygeK cultures were diluted to OD600 = 0.03 in fresh lysogeny broth (LB) broth. Then, 10 µL culture aliquots were spotted onto LB agar plates containing 0.8 mM H2O2 and allowed to air dry [13]. The plates were incubated at 37 °C over night, after which colonies were photographed the next day. Experiments were performed three times.
2.5. Acid Resistance Assays
AE81, AE81ΔygeK, and AE81ΔygeK-pCmygeK cultures were grown to logarithmic phase (OD600 = 1.0) then centrifuged and resuspended in PBS. The pH of LB medium was adjusted with Tris-HCl (100 mmol/L, pH 10.0) to different acid pH levels (pH = 1, pH = 2, pH = 3). Bacteria were cultured in LB at different pH’s at 37 °C for 30 min and then aliquots were spotted onto LB agar plates and incubated overnight. The next day, colonies were counted. All experiments were performed in triplicate.
2.6. iTRAQ-Based Quantitative Proteomic Analysis
The sequenced strains were the original, AE81, and the deletion strain, AE81ΔygeK. Isobaric tags for relative and absolute quantitation (iTRAQ) proteomic services were provided by Shenzhen Huada Gene Co., Ltd. (Shenzhen, China). iTRAQ data were quantified by IQuant [20] software independently developed by BGI, which integrates the Mascot Percolator algorithm [21]. A 1% false-positive rate (FDR) filtering (PSM-level FDR ≤ 0.01) step at the peptide-spectrum match level was performed to obtain spectrum and peptide lists for identification, then proteins were assembled using peptides, and protein groups were generated. Proteins were filtered again using picked protein FDR [22] with FDR at 1% at the protein level (protein-level FDR ≤ 0.01) to control the false positive rate. The Kyoto Encyclopedia of Genes and Genomes Pathway (KEGG-PATH) of differentially expressed proteins was to compare the identified proteins with the Kyoto Encyclopedia of Genes and Genomes (KEGG) database to derive corresponding classification pathway results.
2.7. Statistical Analysis
SPSS (v19.0) software was used to analyze data. Between AE81 and AE81ΔygeK groups, or AE81ΔygeK and AE81ΔygeK-pCmygeK groups, paired t-tests were used for statistical comparisons. A p-value ≤ 0.05 was used to indicate statistical significance.
3. Results
3.1. YgeK Inactivation Decreases Biofilm Formation in AE81 Strains
The influence of ygeK inactivation on biofilm formation was evaluated in vitro. AE81 and AE81ΔygeK-pCmygeK strains formed intact biofilms on glass tubes and liquid surfaces, but the AE81ΔygeK strain generated almost none (Figure 1A). Biofilms were stained in 0.1% crystal violet, dissolved in 33% glacial acetic acid, and read at 492 nm (Figure 1B).
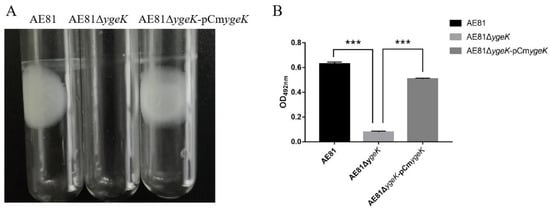
Figure 1.
(A) Biofilms formed on tubes by AE81, AE81ΔygeK, and AE81ΔygeK-pCmygeK strains. (B) Measurement of biofilm mass. Values are average of three independent experiments. Error bars indicate standard deviation (*** p < 0.001).
3.2. Biofilm Observations using SEM
AE81 biofilm morphology under SEM was compact; the outer layer had a thick extracellular matrix and surface bacteria were aggregated to form a dense regional surface layer. YgeK deletion in AE81 resulted in thin membranous structures and large bacterial shapes in the biofilm, whereas AE81ΔygeK-pCmygeK biofilms were more compact than AE81ΔygeK (Figure 2). Inactivation of ygeK resulted in reduced ability of APEC to form biofilms.

Figure 2.
Biofilm structures were observed by scanning electron microscopy (×7000 magnification).
3.3. YgeK Influences APEC Resistance to H2O2
In co-cultured bacteria with H2O2 at different dilutions (10−1, 10−2, 10−3, 10−4, and 10−5), AE81, AE81ΔygeK, and AE81ΔygeK-pCmygeK strains were observed. However, at 10−5, only the AE81 strain was observed. As shown (Figure 3), AE81ΔygeK colony numbers were reduced when compared with AE81 and AE81ΔygeK-pCmygeK, suggesting inactivation of ygeK led to APEC’s response to H2O2 stress.
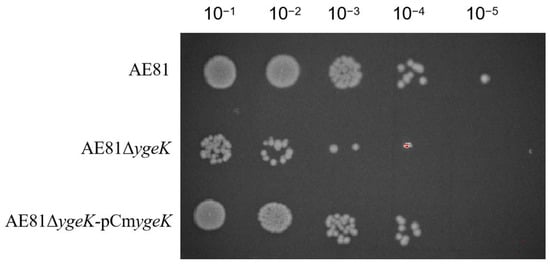
Figure 3.
Hydrogen peroxide sensitivity in AE81, AE81ΔygeK, and AE81ΔygeK-pCmygeKI strains.
3.4. YgeK Influences APEC Acid Resistance
Acid resistance assays are often performed to evaluate a pathogen’s resistance to acid stress environments [23,24]. As shown (Figure 4), ygeK inactivation reduced acid resistance significantly in medium at different pH values (pH 1.0, 2.0, and 3.0).
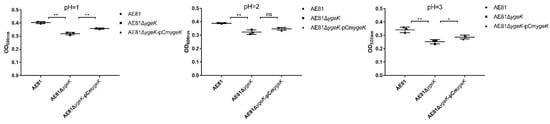
Figure 4.
AE81, AE81ΔygeK, and AE81ΔygeK-pCmygeK resistance to different pH values (pH = 1.0, 2.0, 3.0). Values are average of three independent experiments (* p < 0.05, ** p < 0.01, ns: no significance).
3.5. Screening Differentially Expressed Proteins in AE81 and AE81ΔygeK Strains
In total, 91 significant differentially expressed proteins were identified using fold change > 1.2 and Q-value < 0.05 criteria, of which 26 proteins were significantly up-regulated and 65 significantly down-regulated (Figure 5). The most up-regulated protein was YhaK, which belonged to the Pirin family. The AE81ΔygeK/AE81 ratio was 10. The most down-regulated protein was AdiC which was an arginine: agmatine antiporter (Table S1).
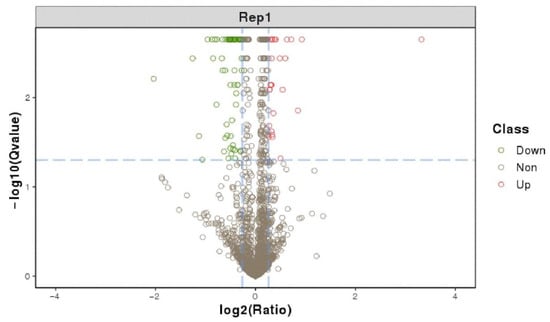
Figure 5.
Differentially expressed protein volcano graph. The X-axis is the protein difference multiple (take log2) and the Y-axis is the corresponding −log10 (Q-value).
3.6. KEGG Pathway Annotation Analysis of Differentially Expressed Proteins
KEGG pathway annotation analyses showed that differentially expressed proteins were mainly enriched in microbial metabolism in diverse environments, two-component systems, flagellar assembly, bacterial chemotaxis, glycolysis/gluconeogenesis, and pyrimidine metabolism, amino sugar and nucleotide sugar metabolism, and fructose and mannose metabolism (Figure 6). Differentially expressed proteins were most prevalent in microbial metabolism in diverse environments, two-component systems, bacterial chemotaxis, and flagellar assembly (Figure 7).
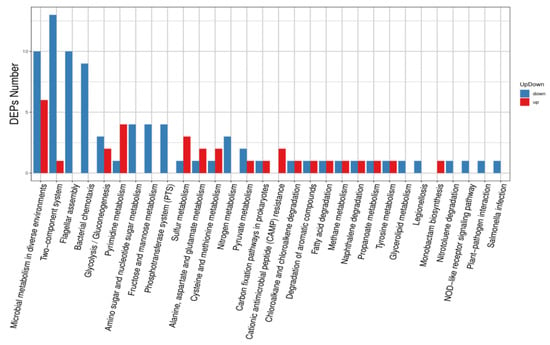
Figure 6.
Up-regulated and down-regulated differentially expressed protein pathway classification.
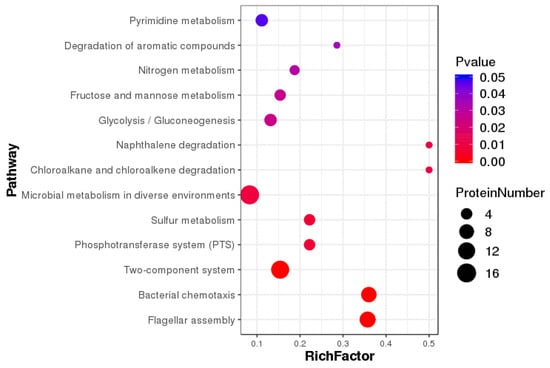
Figure 7.
Enrichment pathways show significant differentially expressed proteins.
4. Discussion
APEC is reportedly a potential zoonotic pathogen that transfers virulence and resistance genes to human ExPEC; therefore, developing effective vaccines or new drug targets is imperative [25]. In our previous study, ygeK significantly affected APEC flagella formation, bacterial motility, serum sensitivity, adhesion, and virulence via different functional pathways [16]. Here, we report that inactivation of the transcriptional activator ygeK decreased biofilm formation and reduced bacterial resistance to acid and oxidative stresses.
Biofilm formation is a protected growth mode that enables pathogen survival in hostile environments [26,27]. Several transcriptional regulators are involved in APEC biofilm formation, including basSR, phoP, mcbR, and cpxA [11,12,13,28]. In bacteria, common regulators, extracellular polymeric production, and biofilm heterogeneity are all central responses and contributors to oxidative stress [29]. For example, Salmonella biofilms cultured in vitro are also tolerant to H2O2 [30]. In APEC, inactivation of ibeA decreased biofilm formation and ibeA can confer increased H2O2 resistance to APEC [31,32]. In our study, ygeK inactivation decreased biofilm formation and reduced oxidative responses in APEC. Inactivation of mcbR increased APEC’s biofilm formation but decreased APEC’s resistance to H2O2 stress [13]. Deletion of waaL increased biofilm formation and reduced resistance to oxidative and alkali environmental stress [33]. Therefore, we hypothesize that these genes affecting biofilm formation and H2O2 resistance are involved in different pathways. H2O2 readily crosses bacterial membranes and enters the cytoplasm where it forms hydroxyl radicals that damage DNA, proteins, and lipid membranes [31]. In APEC, oxidative responses exert important roles during pathogenic processes in hosts. We provide a greater understanding of how APEC is recalcitrant to oxidative stress during chronic infection.
Acid stress is also a typical environmental stress [34]. Microorganisms have developed sophisticated physiological and molecular mechanisms to survive under acid stresses such as the sophisticated acid-resistant systems of E. coli. One acid-resistant system, composed of Glutaminease YbaS and the Glu-GABA antiporter GadC, helps E. coli survive in extremely acid environments [35,36,37]. There are also other factors influencing acid stress. For example, deletion of yfcO decreased survival under acidic stress conditions [38]. The mutation of ybjX decreased resistance to environmental stress (alkaline and acid) [23]. Without ygeK, the acid resistance of APEC decreased, with our proteomics data showing that GadC protein expression was down-regulated, indicating ygeK may affect APEC mediated acid resistance via GadC. This result highlights how ygeK has a role in developing resistance to acid stress.
In this study, we reported that deletion of ygeK decreased biofilm formation ability and reduced resistance to environmental stress. When combined with our previous findings, we hypothesize that ygeK plays a vital function in APEC processes. In addition, we analyzed ygeK function in APEC using proteomics, which not only enriches the APEC network but also unravels APEC pathogenic mechanisms for the generation of alternative control strategies for APEC virulence. In the future, substrate interaction with ygeK should be investigated to further investigate the mechanism.
5. Conclusions
This study proved that transcriptional regulator ygeK influences APEC biofilm formation, resistance to H2O2, and acid resistance. In addition, we provide a greater understanding of how APEC is recalcitrant to oxidative stress during chronic infection, highlight how ygeK has a role in developing resistance to acid stress, and elucidate its regulatory network. These results define the critical role of ygeK in APEC, guiding the search for new drug targets and vaccines.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ani12091160/s1, Table S1: The differentially expressed proteins between AE81 and AE81ΔygeK.
Author Contributions
Conceptualization, M.X. and D.F.; methodology, M.X. and D.F.; software, J.H.; validation, M.X. and D.F. and J.H.; formal analysis, J.H.; investigation, Y.S.; resources, K.Q.; data curation, X.S.; writing—original draft preparation, M.X.; writing—review and editing, J.T. and X.S.; visualization, M.X. and D.F.; supervision, J.T. and Y.S.; project administration, Y.S.; funding acquisition, M.X. and K.Q. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the National Natural Science Foundation of China (31772707) and Jinling Institute of Technology Scientific research start-up fund for high-end talents (jit-b-202154), and “The APC was funded by a Jinling Institute of Technology Scientific research start-up fund for high-end talents (jit-b-202154).
Institutional Review Board Statement
Not applicable. This study did not involve humans or animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
iTRAQ-based proteomic sequencing data were deposited into the iProX integrated proteome resources with Project ID IPX0004369000.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kathayat, D.; Lokesh, D.; Ranjit, S.; Rajashekara, G. Avian Pathogenic Escherichia coli (APEC): An Overview of Virulence and Pathogenesis Factors, Zoonotic Potential, and Control Strategies. Pathogens 2021, 10, 467. [Google Scholar] [CrossRef] [PubMed]
- Mellata, M. Human and avian extraintestinal pathogenic Escherichia coli: Infections, zoonotic risks, and antibiotic resistance trends. Foodborne Pathog. Dis. 2013, 10, 916–932. [Google Scholar] [CrossRef]
- Perreten, V.; Strauss, C.; Collaud, A.; Gerber, D. Colistin Resistance Gene mcr-1 in Avian-Pathogenic Escherichia coli in South Africa. Antimicrob. Agents Chemother. 2016, 60, 4414–4415. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, N.L.; Pimenta, R.L.; de Melo, D.A.; Nolan, L.K.; de Souza, M.M.S.; Logue, C.M. mcr-1 Identified in Fecal Escherichia coli and Avian Pathogenic E. coli (APEC) from Brazil. Front. Microbiol. 2021, 12, 659613. [Google Scholar] [CrossRef] [PubMed]
- Lima Barbieri, N.; Nielsen, D.W.; Wannemuehler, Y.; Cavender, T.; Hussein, A.; Yan, S.G.; Nolan, L.K.; Logue, C.M. mcr-1 identified in Avian Pathogenic Escherichia coli (APEC). PLoS ONE 2017, 12, e0172997. [Google Scholar] [CrossRef]
- Monds, R.D.; O’Toole, G.A. The developmental model of microbial biofilms: Ten years of a paradigm up for review. Trends Microbiol. 2009, 17, 73–87. [Google Scholar] [CrossRef]
- Hobley, L.; Harkins, C.; MacPhee, C.E.; Stanley-Wall, N.R. Giving structure to the biofilm matrix: An overview of individual strategies and emerging common themes. FEMS Microbiol. Rev. 2015, 39, 649–669. [Google Scholar] [CrossRef]
- Gebreyohannes, G.; Nyerere, A.; Bii, C.; Sbhatu, D.B. Challenges of intervention, treatment, and antibiotic resistance of biofilm-forming microorganisms. Heliyon 2019, 5, e02192. [Google Scholar] [CrossRef]
- Nahar, S.; Jeong, H.L.; Kim, Y.; Ha, A.J.; Roy, P.K.; Park, S.H.; Ashrafudoulla, M.; Mizan, M.F.R.; Ha, S.D. Inhibitory effects of Flavourzyme on biofilm formation, quorum sensing, and virulence genes of foodborne pathogens Salmonella Typhimurium and Escherichia coli. Food Res. Int. 2021, 147, 110461. [Google Scholar] [CrossRef]
- Peng, L.Y.; Yuan, M.; Cui, Z.Q.; Wu, Z.M.; Yu, Z.J.; Song, K.; Tang, B.; Fu, B.D. Rutin inhibits quorum sensing, biofilm formation and virulence genes in avian pathogenic Escherichia coli. Microb. Pathog. 2018, 119, 54–59. [Google Scholar] [CrossRef]
- Yu, L.; Wang, H.; Han, X.; Li, W.; Xue, M.; Qi, K.; Chen, X.; Ni, J.; Deng, R.; Shang, F.; et al. The two-component system, BasSR, is involved in the regulation of biofilm and virulence in avian pathogenic Escherichia coli. Avian Pathol. 2020, 49, 532–546. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Li, Q.; Xue, M.; Wang, Z.; Tu, J.; Song, X.; Shao, Y.; Han, X.; Xue, T.; Liu, H.; et al. The role of the phoP transcriptional regulator on biofilm formation of avian pathogenic Escherichia coli. Avian Pathol 2019, 48, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Li, W.; Qi, K.; Wang, S.; Chen, X.; Ni, J.; Deng, R.; Shang, F.; Xue, T. McbR is involved in biofilm formation and H2O2 stress response in avian pathogenic Escherichia coli X40. Poult. Sci. 2019, 98, 4094–4103. [Google Scholar] [CrossRef] [PubMed]
- Hartleib, S.; Prager, R.; Hedenstrom, I.; Lofdahl, S.; Tschape, H. Prevalence of the new, SPI1-like, pathogenicity island ETT2 among Escherichia coli. Int. J. Med. Microbiol. 2003, 292, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Makino, K.; Ohnishi, M.; Kurokawa, K.; Ishii, K.; Yokoyama, K.; Han, C.G.; Ohtsubo, E.; Nakayama, K.; Murata, T.; et al. Complete genome sequence of enterohemorrhagic Escherichia coli O157:H7 and genomic comparison with a laboratory strain K-12. DNA Res. 2001, 8, 11–22. [Google Scholar] [CrossRef]
- Tu, J.; Fu, D.; Gu, Y.; Shao, Y.; Song, X.; Xue, M.; Qi, K. Transcription Regulator YgeK Affects the Virulence of Avian Pathogenic Escherichia coli. Animals 2021, 11, 3018. [Google Scholar] [CrossRef]
- Guttenplan, S.B.; Kearns, D.B. Regulation of flagellar motility during biofilm formation. FEMS Microbiol. Rev. 2013, 37, 849–871. [Google Scholar] [CrossRef]
- Xue, M.; Xiao, Y.; Fu, D.; Raheem, M.A.; Shao, Y.; Song, X.; Tu, J.; Xue, T.; Qi, K. Transcriptional Regulator YqeI, Locating at ETT2 Locus, Affects the Pathogenicity of Avian Pathogenic Escherichia coli. Animals 2020, 10, 1658. [Google Scholar] [CrossRef]
- Li, Q.; Yin, L.; Xue, M.; Wang, Z.; Song, X.; Shao, Y.; Liu, H.; Tu, J.; Qi, K. The transcriptional regulator PhoP mediates the tolC molecular mechanism on APEC biofilm formation and pathogenicity. Avian Pathol. 2020, 49, 211–220. [Google Scholar] [CrossRef]
- Wen, B.; Zhou, R.; Feng, Q.; Wang, Q.; Wang, J.; Liu, S. IQuant: An automated pipeline for quantitative proteomics based upon isobaric tags. Proteomics 2014, 14, 2280–2285. [Google Scholar] [CrossRef]
- Brosch, M.; Yu, L.; Hubbard, T.; Choudhary, J. Accurate and sensitive peptide identification with Mascot Percolator. J. Proteome Res. 2009, 8, 3176–3181. [Google Scholar] [CrossRef] [PubMed]
- Savitski, M.M.; Wilhelm, M.; Hahne, H.; Kuster, B.; Bantscheff, M. A Scalable Approach for Protein False Discovery Rate Estimation in Large Proteomic Data Sets. Mol. Cell. Proteom. 2015, 14, 2394–2404. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Qiu, M.; Jiang, H.; Xue, M.; Hu, J.; Liu, H.; Zhou, X.; Tu, J.; Qi, K. ybjX mutation regulated avian pathogenic Escherichia coli pathogenicity though stress-resistance pathway. Avian Pathol. 2020, 49, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Bao, Y.; Meng, Q.; Xia, Y.; Zhao, Y.; Wang, Y.; Tang, F.; ZhuGe, X.; Yu, S.; Han, X.; et al. IbeR facilitates stress-resistance, invasion and pathogenicity of avian pathogenic Escherichia coli. PLoS ONE 2015, 10, e0119698. [Google Scholar] [CrossRef]
- Logue, C.M.; Wannemuehler, Y.; Nicholson, B.A.; Doetkott, C.; Barbieri, N.L.; Nolan, L.K. Comparative Analysis of Phylogenetic Assignment of Human and Avian ExPEC and Fecal Commensal Escherichia coli Using the (Previous and Revised) Clermont Phylogenetic Typing Methods and its Impact on Avian Pathogenic Escherichia coli (APEC) Classification. Front. Microbiol. 2017, 8, 283. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef]
- Andrews, J.S.; Rolfe, S.A.; Huang, W.E.; Scholes, J.D.; Banwart, S.A. Biofilm formation in environmental bacteria is influenced by different macromolecules depending on genus and species. Environ. Microbiol. 2010, 12, 2496–2507. [Google Scholar] [CrossRef]
- Matter, L.B.; Ares, M.A.; Abundes-Gallegos, J.; Cedillo, M.L.; Yanez, J.A.; Martinez-Laguna, Y.; De la Cruz, M.A.; Giron, J.A. The CpxRA stress response system regulates virulence features of avian pathogenic Escherichia coli. Environ. Microbiol. 2018, 20, 3363–3377. [Google Scholar] [CrossRef]
- Gambino, M.; Cappitelli, F. Mini-review: Biofilm responses to oxidative stress. Biofouling 2016, 32, 167–178. [Google Scholar] [CrossRef]
- Hahn, M.M.; Gonzalez, J.F.; Gunn, J.S. Salmonella Biofilms Tolerate Hydrogen Peroxide by a Combination of Extracellular Polymeric Substance Barrier Function and Catalase Enzymes. Front. Cell. Infect. Microbiol. 2021, 11, 683081. [Google Scholar] [CrossRef]
- Flechard, M.; Cortes, M.A.; Reperant, M.; Germon, P. New role for the ibeA gene in H2O2 stress resistance of Escherichia coli. J. Bacteriol. 2012, 194, 4550–4560. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, S.; Niu, C.; Shi, Z.; Xia, Y.; Yaqoob, M.; Dai, J.; Lu, C. Effects of ibeA deletion on virulence and biofilm formation of avian pathogenic Escherichia coli. Infect. Immun. 2011, 79, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Han, X.; Wang, S.; Meng, Q.; Zhang, Y.; Ding, C.; Yu, S. The waaL gene is involved in lipopolysaccharide synthesis and plays a role on the bacterial pathogenesis of avian pathogenic Escherichia coli. Vet. Microbiol. 2014, 172, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Guan, N.; Liu, L. Microbial response to acid stress: Mechanisms and applications. Appl. Microbiol. Biotechnol. 2020, 104, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhao, Z.; Tong, W.; Ding, Y.; Liu, B.; Shi, Y.; Wang, J.; Sun, S.; Liu, M.; Wang, Y.; et al. An acid-tolerance response system protecting exponentially growing Escherichia coli. Nat. Commun. 2020, 11, 1496. [Google Scholar] [CrossRef]
- Brameyer, S.; Hoyer, E.; Bibinger, S.; Burdack, K.; Lassak, J.; Jung, K. Molecular design of a signaling system influences noise in protein abundance under acid stress in different gamma-Proteobacteria. J. Bacteriol. 2020, 202, e00121-20. [Google Scholar] [CrossRef]
- Lu, P.; Ma, D.; Chen, Y.; Guo, Y.; Chen, G.Q.; Deng, H.; Shi, Y. L-glutamine provides acid resistance for Escherichia coli through enzymatic release of ammonia. Cell Res. 2013, 23, 635–644. [Google Scholar] [CrossRef]
- Li, Y.; Wang, H.; Ren, J.; Chen, L.; Zhuge, X.; Hu, L.; Li, D.; Tang, F.; Dai, J. The YfcO fimbriae gene enhances adherence and colonization abilities of avian pathogenic Escherichia coli in vivo and in vitro. Microb. Pathog. 2016, 100, 56–61. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).