Simple Summary
The involvement of the oviductal epithelial cells and progesterone hormone on the sperm fertilizing ability was investigated here by using an in vitro fertilization (IVF) swine model. By using different techniques, we demonstrated that oviductal epithelial cells (OECs) pre-treated with progesterone (P4) at 100 ng/mL represent the ideal environment for sperm capacitation, inducing a positive effect on the sperm acquisition of the fertilizing ability. Moreover, treatment of OECs with P4 modifies OVGP1, SPP1 and DMBT1 gene transcription but does not interfere with important cytoskeleton structures or cell doubling time, thereby allowing the correct development and progression of important biochemical pathways and signal transduction processes.
Abstract
Mammalian spermatozoa are infertile immediately after ejaculation and need to undergo a functional modification, called capacitation, in order to acquire their fertilizing ability. Since oviductal epithelial cells (SOECs) and progesterone (P4) are two major modulators of capacitation, here we investigated their impact on sperm functionality by using an IVF swine model. To that, we treated SOECs with P4 at 10, 100, and 1000 ng/mL before the coincubation with spermatozoa, thus finding that P4 at 100 ng/mL does not interfere with the cytoskeleton dynamics nor the cells’ doubling time, but it promotes the sperm capacitation by increasing the number of spermatozoa per polyspermic oocyte (p < 0.05). Moreover, we found that SOECs pre-treatment with P4 100 ng/mL is able to promote an increase in the sperm fertilizing ability, without needing the hormone addition at the time of fertilization. Our results are probably due to the downregulation in the expression of OVGP1, SPP1 and DMBT1 genes, confirming an increase in the dynamism of our system compared to the classic IVF protocols. The results obtained are intended to contribute to the development of more physiological and efficient IVF systems.
1. Introduction
Human infertility is estimated to affect approximately 48 million couples and 186 million individuals in the world [1,2], thus raising serious concerns in the last several decades. Currently, the efficacy of the assisted reproductive technology (ART) procedures is considered still considered to be far from optimal if compared with natural conception, in which the process of fertilization is subjected to the dynamics of the in vivo environment rather than to the static artificial conditions.
In vivo, the process of fertilization in mammals occurs within the oviduct, a complex structure that only a few spermatozoa are able to reach and where they strongly bind to the epithelial cells, forming a functional sperm reservoir [3,4]. During this storage, the interactions between the oviductal epithelial cells (OECs) and spermatozoa are thought to play an important role in sperm selection [5], sperm viability maintenance [6], and premature capacitation prevention [7]. After an indefinite period of time and around the time of ovulation, spermatozoa are released from the OECs to continue swimming towards the fertilization site, where they reach the egg [5]. The dynamic context created by the oviduct environment is characterized by the involvement of several molecules. Among these, progesterone (P4) plays an important role in spermatozoa. P4 is a steroid hormone produced in several organs and tissues, such as the placenta [8], the adrenal cortex, the ovary and the testis [9,10]. P4 secreted from the ovary arrives to the oviduct through a local countercurrent transfer [11], where it is able to act on the sperm cells. Among the multiple and wide-studied functions of P4 on spermatozoa it is worth to increasing the regulation of sperm hyperactivation [12] by increasing the intracellular concentration of calcium [Ca2+] mediated by CatSper channels (pH-dependent Ca2+ channel of the sperm flagellum) [13], the sperm chemotaxis towards the mature oocyte [14,15], the induction of acrosome reaction (AR) [16,17], and the promotion of capacitation in vitro [18,19]. Moreover, in the in vitro swine model, P4 has been observed to increase the sperm-binding capacity in to SOECs previously treated with E2 and P4, simulating estrus [20]. However, some important information is still missing regarding the effects caused by P4 directly on the oviductal cells that interact with the male gametes and that could also exert an indirect function on sperm cells.
The oviduct plays a key role as well also in the genetic and epigenetic reprogramming derived from the embryo-maternal interactions that take place during the early embryo development [21]. The significant increase in the prevalence of disorders generated to the embryo after ART procedures is worthy of note. For instance, the phenomenon of large offspring syndrome (LOS) in farm animals is characterized by macrosomia, macroglossia, omphalocele and abnormal organ growth [22], with a phenotypic similarity to the Beckwith-Wiedemann syndrome (BWS) in humans [22,23,24]. Many reports also showed the correlation between ART and Angelman syndrome (AS), a rare neurogenetic human syndrome characterized by severe mental retardation, the absence of speech, ataxia, seizures, an abnormal EEG pattern and hyperactivity [25,26]. The increasing prevalence of these pathologies presents an urgent need to improve ART protocols, progressing as much as possible towards the replication of the physiological conditions.
Several researchers around the globe are dedicated to the study of different approaches to improve IVF protocols, for instance by integrating oviductal fluid and uterine fluid in IVF [27,28] and embryo culture media [28], and with the introduction of 3-D oviductal epithelial cell culture systems to mimic the oviduct [29]. At present, the studies available to date are not focused on developing new strategies to increase the spermatozoa fertilizing ability.
Here, we propose the implementation of the sperm fertilizing ability using an IVF system that includes the somatic component (a monolayer of swine OECs, SOECs) and the addition of different P4 concentrations (10, 100, 1000 ng/mL [30,31,32]). We used a multiple-combinations strategy: on one hand, spermatozoa were capacitated in the presence of SOECs previously treated with P4; on the other hand, spermatozoa were capacitated both in the presence of SOECs and P4 at different concentrations. Thus, the final aim of this study was to evaluate the effects caused by P4 addition to both the somatic component and the gametes in an effort to approximate the IVF protocols to a more physiological condition.
2. Materials and Methods
2.1. Chemicals
Unless otherwise stated, all the chemicals were purchased from Sigma Aldrich (St. Louis, MO, USA).
2.2. SOECs Collection and Incubation
Swine oviducts at peri-ovulatory stages were collected from a local slaughterhouse and dissected. SOECs were gathered by scratching the whole oviduct (ampulla and isthmus tracts) following an established protocol [30,31]. Cells were pooled and then rinsed with a washing medium (TCM 199 supplemented with FBS 10%, penicillin/streptomycin 2% and amphotericin B 1%). SOECs were then cultured in 12-well plates in a medium containing TCM 199 supplemented with FBS 10% and penicillin/streptomycin 2% [29]. SOEC were treated with P4 at different concentrations depending on the experiment (10, 100, 100 ng/mL) and 17-β-estradiol (E2, 4 ng/mL), maintaining a control sample and incubated in a humidified atmosphere with 5% CO2 at 38.5 °C (Hera Cell Thermo Fisher Scientific, Monza, MB, Italy). The medium was renewed every 48 h.
2.3. Cell Population Doubling Time
To calculate the doubling time of SOECs, cells were cultured at a concentration of 1 × 106 cells/mL, following the protocol already described and treated with the different P4 concentrations. After 48 h and seven days, cells were trypsinized and counted. The SOECs doubling time was calculated (Roth V. 2006 Doubling Time Computing, Available from: http://www.doubling-time.com/compute.php accessed on 1 March 2021) as:
where:
DT = doubling time
FC = final concentration
IC = initial concentration
2.4. Phalloidin Staining
To assess the effect of progesterone treatment on actin polymerization, SOECs were cultured in 3.5 cm Petri dishes (38.5 °C, 5% CO2 and humidified atmosphere) and treated with the three different P4 concentrations. Once they reached 50% and 100% of confluence, SOECs were fixed overnight in paraformaldehyde 4%, then washed three times with PBS for 5 min each prior to the incubation with TRITC-conjugated phalloidin 488 (50 μg/mL in PBS, dilution 1:100) for 1 h. After three washes with PBS for 5 min, cell samples were mounted with Vectashield mounting medium (Vector H-1000) and subjected to microscopy analysis.
2.5. Tubulin Immunocytochemistry
SOECs were cultured with a cover glass, treated, and fixed as described above. After overnight fixation and washing, samples were permeabilized with 5% Triton X-100 in PBS for 30 min. SOECs were washed three times in PBS for 5 min and then incubated with BSA 5% + Normal Goat Serum (NGS) 5% in PBS for 30 min to block unspecific binding of the antibodies. Cells were then incubated overnight at 4 °C with monoclonal anti-mouse-α-tubulin antibody (SIGMA 025k4809) diluted 1:500 in PBS, containing 1% BSA. After washing, the bound antibody was detected using anti-mouse cy3 TRICT-conjugated diluted 1:500 in PBS containing 1% BSA incubated for 1 h at room temperature and followed by washing three times at 5 min intervals with 1% BSA—05% Tween20—PBS. Finally, the cover glasses were stained with DAPI 1:1000 and mounted in Vectashield mounting medium (Vector H-1000). SOECs samples incubated with PBS without the primary antibody were used as negative controls.
2.6. Confocal Analysis of SOECr
The acquisition was realized with a Nikon A1r laser confocal scanning microscope equipped with a Plan Apo λ 100X Oil objective and a Galvano detector with a pinhole size of 69 μm and a pixel size 0.04 µm. We used an averaged 2 mode in channels series as follows:
Channel 1: DAPI: λexc = 404 nm; λem = 450/50 nm
Channel 2: TRITC: λexc = 561.5 nm; λem = 595/50 nm
2.7. Preparation and Incubation of Sperm Samples
The preparation of boar semen samples was carried out following an already standardized protocol [33]. In brief, sperm samples were purchased from GENEETIC SRL, Modena, Italy. Before each experiment, sperm motility was visually estimated and only samples with a sperm motility >80% were considered for further analyses. To achieve capacitation, spermatozoa were incubated with confluent SOECs (after eight days of culture) in a capacitating medium TCM199 supplemented with 13.9 mM glucose, 1.25 mM sodium pyruvate, 2.25 mM calcium lactate and 1 mM of caffeine [34] at a final concentration of 1 × 107 cells/mL and up to 1.5 h, at 38.5 °C in 5% CO2 humidified atmosphere (Heraeus, Hera Cell). The concentration of P4 100 ng/mL was added to the capacitation medium, preserving a control sample without the hormone.
2.8. In Vitro Fertilization
To obtain mature oocytes necessary for the IVF, ovaries from gilts were collected at a local slaughterhouse and transported to the laboratory within 1 h of slaughter maintaining a temperature of 25 °C. After washing the ovaries in a normal saline solution and bialcohol, the follicles of 4/5 mm diameter were selected on the basis of their translucent appearance, good vascularization and compactness of their granulosa layer and cumulus mass [35]. COCs (cumulus-oocyte complex) were collected by aspirating healthy selected follicles. The maturation process to the MII stage was obtained in vitro by culturing the COCs in four-well dishes containing α-MEM medium added with 10% FBS, 1% Penicillin/Streptomycin, 1% Ultraglutamine, hCG (5 UI/mL) and PMSG (5 UI/mL) for 44 h at 38.5 °C in a humidified atmosphere with 5% CO2 (Heraeus, Hera Cell) [35].
At the end of the maturation, the oocytes were denuded in DPBS with hyaluronidase on a warmed stage at 38.5 °C under a stereomicroscope. Only oocytes presenting the first polar body (MII stage) under the stereomicroscope were utilized for the IVF assay, using the same groups described above.
The day of the fertilization, swine spermatozoa were washed and incubated as previously described. Then, the capacitated spermatozoa at a final concentration of 1 × 106 cells/mL were incubated with the oocytes in the presence of SOECs in a fertilization medium (capacitation medium supplemented with 10% FBS [35]). After 3 h, the potential fertilized oocytes were removed from the Petri dish, transferred into fresh medium and maintained in culture for at least 12 h. Subsequentially, the oocytes were fixed in Glutaraldehyde 0.5%, stained with Hoechst 33342 and observed with a fluorescence microscope.
As the experimental design (Figure 1) shows, IVF experiments were carried out in two steps: first, we perform the experiments using the combination of SOECs, spermatozoa and the three different P4 concentrations (10,100, 1000 ng/mL) to identify the most effective concentration (100 ng/mL) (Figure 1A), and then we performed the IVF using four different combinations, including SOECs not treated with P4 and sperm capacitation in the presence of P4, SOECs treated with P4 and sperm capacitation either with or without P4, and control conditions (Figure 1B). For both sets of experiments, the control group corresponding to spermatozoa capacitated in the presence of SOECs without pre-treatment with P4 nor P4 addition during capacitation.
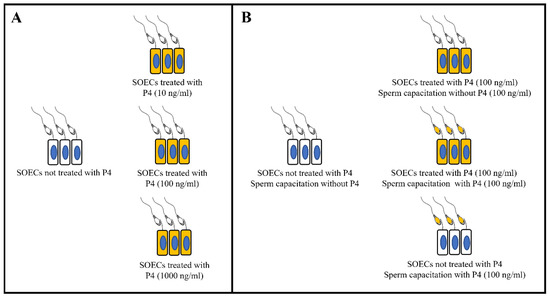
Figure 1.
Experimental design for IVF. (A) Experimental groups to identify the most effective P4 concentration. (B) Experimental groups using the most effective P4 concentration and sperm capacitation with and without P4 (100 ng/mL) in the presence of SOECs previously treated or not with P4.
IVF outcomes have been expressed as the fertilization rate (% of penetrated oocytes), the incidence of polyspermy (% of polyspermic oocytes), and the number of penetrating spermatozoa/polyspermic oocytes according to already published works [36,37]. We performed eight independent experiments, reaching a total number of 291 oocytes.
2.9. Gene Expression Analysis
Total RNA was extracted using the Total RNA Purification Kit (Cat. 17259, NORGEN Biotek Corp. Thorold, ON, Canada) according to the manufacturer’s instructions. Next, total RNA was evaluated and quantified using a Thermo Scientific NanoDrop 2000c UV-Vis spectrophotometer at 260 nm and stored at −80 °C until use in reverse transcription (RT). For the reverse transcription, 1 µg of total RNA from each sample was reverse transcribed into complementary DNA using an RT reaction with a Random Hexamers primer, (dNTP) mix and Tetro Reverse Transcriptase (Bioline, Luckenwalde, Germany) in a 20 µL reaction volume mixture according to the manufacturer’s instructions. cDNAs were diluted three times (60 µL final volume) and stored at −20 °C until used as template in an RT-qPCR mixture. Quantitative real-time PCR (qPCR) was carried out using primer sequences for GAPDH, OVGP1, DMBT1 and SPP1 genes that codify for glyceraldehyde 3-phosphate dehydrogenase and oviduct specific glycoprotein, deleted in malignant brain tu mors 1 and osteopontin proteins, respectively. The primers for GAPDH and OVGP1 genes had been previously published [38]. The primers for DMBT1 and SPP1 genes were designed by using the Primer BLAST Tool (https://www.ncbi.nlm.nih.gov/tools/primer-blast/ 1 February 2021) and according to Thornton and Basu [39]. The primer sequences and their annealing temperatures are listed in Table 1. The specificity of the qPCR product was verified using the melting curve analysis program. The RT-qPCR reaction consisted of 13 µL master mix containing SensiFAST TM SYBR Lo-ROX kit (Bioline), primers and 2 µL of the diluted cDNA in a total volume of 15 µL, according to the manufacturer’s instructions. The cycling program was the two-step cycling protocol for 40 cycles (10 s at 95 °C for denaturation and 30 s at 60 °C for annealing/extension) with a 7500 Fast Real-time PCR System (Thermo Fisher Scientific, Monza, MB, Italy) followed by melt-profile analysis (7500 Software v2.3). For each qPCR analysis, each sample was performed in triplicate, and values were normalized to GAPDH endogenous reference gene (housekeeping gene). The relative gene expression was calculated by the comparative Ct (ΔΔCt) method and converted to the relative expression ratio (2−ΔΔCt).

Table 1.
Primer sequences and corresponding annealing temperatures for RT-qPCR analysis.
2.10. Statistical Analysis
For statistical analysis, GraphPad Prism 6 Software (La Jolla, CA, USA) was used. Data were checked for normal distribution with a D’Agostino and Pearson normality test prior to perform the comparison with parametric or non-parametric tests, as required. In all cases the differences among groups were considered statistically significant when p < 0.05. To assess the effect of different treatments on IVF we carried out eight independent experiments using samples from different animals. An a priori power analysis was done to establish the number of oocytes with G*Power 3.1.9.7 software to get a final power of our analysis ≥95%. The results were expressed as the difference between the CTRL and treated samples, expressed as a percentage on the CTRL (ΔIVF%) [40]. To analyze the IVF data, Tukey’s and Dunnett’s multiple comparison tests were used.
To evaluate the effect of different treatments on gene expression, we carried out three independent experiments. The results of a D’Agostino and a Pearson normality test indicated a non-Gaussian distribution of the data and the controls are characterized by a variance = 0 (by definition the CRTL value is 1). Thus, we used the Kruskal Wallis test, and the data are expressed as median +/− range.
3. Results and Discussion
Fertilization is a complex process involving several essential components and an ensemble of connected processes. Among them, spermatozoa acquisition of the fertilizing ability through the process of capacitation is a key one, allowing them to interact with the mature oocytes within the oviduct. All these events are coordinated by the neuroendocrine axis through the dynamic changes induced by some steroid hormones such as P4, which modulates the oviductal environment and prepares the spermatozoa to fertilize the egg. The aim of the present work was to evaluate the action on the sperm fertilizing ability of two major modulators of capacitation, the oviductal epithelial cells and progesterone hormone, in an IVF model.
To achieve our goal, we performed two different sets of experiments:
- The identification of the optimal P4 concentration among three P4 concentrations (10, 100, 1000 ng/mL) [30,31] on morpho-functional parameters:
- Cell doubling time
- Actin organization
- Tubulin organization
The effects, in terms of sperm fertilizing ability in IVF assay, of SOECs exposure to P4 were calculated prior to their use as the environment for sperm capacitation.
- Once the most favourable P4 concentration (100 ng/mL) was identified, we performed the comparison with further experimental conditions of interest (as shown in Figure 1B) to evaluate the sperm fertilizing ability, both with the SOECs treatment with or without P4 (100 ng/mL) and the addition or not of P4 (100 ng/mL) during capacitation.
3.1. P4 Supplementation Does Not Affect the Cells Growing Rate
We used different biological parameters to study the effects of P4 treatment on cell physiology. As with all the steroids, P4 is a lipophilic hormone, thus hampering the monitoring of its real bioavailable concentration regardless of the initial concentration of P4 with which SOECs are treated.
To explore whether the treatment of SOECs with P4 had effects on the cells’ growth and division, we evaluated the cells doubling time (DB) [41]. The cell doubling time is an important parameter used for the description of the cell development dynamics [42] and corresponds to the time it takes for a cell population to double in size [43]. It depends on different parameters: the time required for cell adhesion to the substrate; the cell adaptation to the artificial environment they are exposed to; and the time necessary for cell division [43]. In a cell culture, after the initial growth suppression a fast increase in the growth rate can be observed, followed by a constant exponential growth phase during the rest of the cell cycle [44]. Thus, with an exponential growth curve, the DB is an accurate measure of population cell growth [43]. We have calculated the DB of SOECs at two time points, 48 h and seven days. As shown in Figure 2, the doubling time was not affected by the presence of P4 at any of the three different concentrations (p > 0.05). Since this parameter is related with the ability of a cell culture to grow and adapt to the environmental conditions, these findings demonstrate that P4 does not exert a negative or toxic effect for the SOECs culture, independently on the concentration (within the range 10–1000 ng/mL).
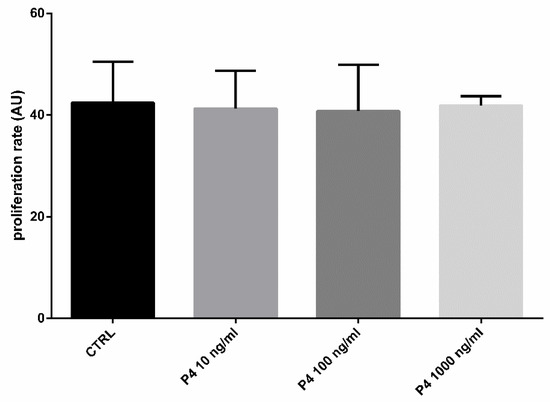
Figure 2.
Effect of the different P4 concentrations on SOECs doubling time. The histograms show a normal proliferation rate of the samples treated with different concentrations of P4 (10, 100 and 1000 ng/mL), similar to the control (CTRL) group (p > 0.05). Three independent experiments were performed.
3.2. P4 Supplementation Does Not Modify the SOECs Cytoskeleton
The cell cytoskeleton is composed of actin filaments, intermediate filaments and α- and β-tubulins that polymerize into microtubules. Due to the great importance of the cytoskeleton for the biological functions of cells such us migration, morphogenesis, cytokinesis, endocytosis, phagocytosis and response to extracellular stimuli [45], here we examined the effects of the SOECs pre-treatment with P4 directly on the actin filaments and tubulin patterns. Interestingly, either the F-actin organization and tubulin patterns remains unaffected by P4 treatment, as displayed respectively in Figure 3.
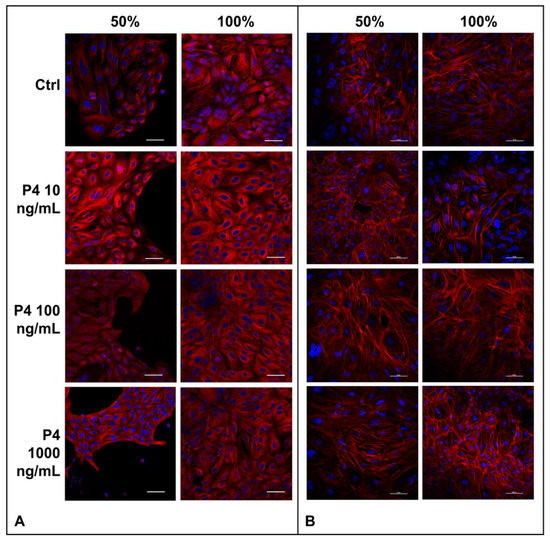
Figure 3.
(A) Microtubules organization of SOECs. Immunofluorescence analysis of α-tubulin of SOECs after P4 treatment at different confluence time points (50% e 100%) and control condition (CTRL). Monoclonal anti-mouse-α-tubulin antibody and secondary antibody anti-mouse cy3 TRICT-conjugated (red) were used to detect α-tubulin; DAPI was used to stain the nuclei (blue). Three independent experiments were performed. (B) Actin filaments of SOECs. Phalloidin staining was used to study the F-actin of SOECs after P4 treatments at different confluence time points (50% e 100%) and control condition (CTRL). DAPI (blue, nuclei), TRITC-conjugated phalloidin (red, F-actin). Three independent experiments were performed.
Actins are a family of globular multi-functional proteins that polymerize to form filamentous structures. The most important physiological function of actin filaments is to produce force to be applied, for instance in cell migration and morphogenesis [46,47].
The other important protein examined here, tubulin, is a protein superfamily of globular proteins. α- and β-tubulins polymerize into microtubules, a major component of the eukaryotic cytoskeleton with an essential function in cell biology, such as mitosis, DNA segregation and cell division [48]. Tubulin and actin are involved in the dynamical arrangement of the cell cytoskeleton in response to intracellular (Ca2+, ATP and other second messengers) and extracellular (hormones, growth factors, ECM proteins) stimuli, and are able to control each other [49].
Abnormalities of these structures are associated with many pathological disorders. For instance, the altered expression of tubulin isotypes, alterations in tubulin post-translational modifications and changes in the expression of microtubule-associated proteins are associated with cancer [45,46,50]. Furthermore, the cytoskeleton alterations are involved in neurological [51], neurodevelopmental and neurodegenerative disorders [52]. For example, mutations in the DCX and LIS1 genes that encode for microtubule-associated proteins are related to migration defects leading to the lack of development of brain folds and grooves [52].
Thus, from the results derived from the present work showing no alterations of these structures, we could affirm that P4 supplementation does not interfere with the development and progression of important biochemical pathways, thus not carrying a dysregulation of the signal transduction.
3.3. Sperm Capacitation on SOECs Previously Supplemented with P4 100 ng/mL Significantly Increases the IVF Outcomes
Based on the great importance of the oviduct and the neuroendocrine female axis in the fertilization process, we designed an IVF protocol using oviductal epithelial cells pre-treated with well-studied concentrations of P4 for the capacitation of the sperm cells [30,31,32].
Mammalian spermatozoa are not able to fertilize immediately after ejaculation, but the ability to fertilize the egg is only acquired after a series of changes that modify some of their biochemical, chemical, or physicochemical features within a process known as capacitation, which takes place during the transit of spermatozoa through the female genital tract [53,54]. This maternal environment operates a sperm selection as well, resulting in a very low percentage of sperm reaching the fertilization site. Thus, after swimming up the uterus and crossing the utero-tubal junction, the sperm arrive to the isthmus (the distal part of the oviduct), and release some messenger biomolecules that modulate the gene expression in epithelial cells before the physical contact [55].
Sperm cells bind preferentially to ciliated epithelial cells [56] through specific carbohydrate residues or the lectin-like proteins, thus forming the sperm reservoir [53,57]. Only motile, non-capacitated, acrosome-intact and normal chromatin structure spermatozoa are able to bind to the oviductal epithelium [58], extending their lifespan and delaying capacitation [59]. The mechanisms of sperm detachment are still partially unknown, but it is well-accepted that ovulation-associated signals induce the sperm release for the transit upper the oviduct tract (the ampulla) where fertilization occurs [5,60]. Numerous factors have been demonstrated to be involved in the release of spermatozoa, including ovarian steroid hormones [30,31,32,61], increased calcium levels [62], and anandamide [62], among others.
In our experiments, we first attempted to identify the most favorable P4 concentration to use as a supplement during SOECs growth prior to IVF. As it is evident from the IVF outcomes analysis (see Figure 4), the best results were obtained when performing capacitation on SOECs previously treated with P4 100 ng/mL, this concentration thus being considered to be the most effective one. It is possible to use the IVF assay to evaluate the sperm fertilizing ability in boar [37,63] due to the fact that the fertilization rates, the number of polyspermic oocytes and number of spermatozoa/polyspermic oocytes are related to the capacitation status and fertility of the semen [64,65]. Here, we used the IVF as a functionality test for spermatozoa to evaluate the capacitation status of sperm cells, instead that for producing embryos. In swine, in vitro maturated oocytes are partially unable to avoid the polyspermic fertilization [37,63], thus in specific conditions and in terms of sperm/oocytes ratio and length of coincubation it is possible to obtain polyspermic oocytes as a marker of their fertilizing ability. Multiple hypotheses arise that should be investigated to explain these results as the interaction between extracellular vesicles (EVs) and spermatozoa [66,67]. In a recent study, Asaadi and co-workers isolated EVs from oviductal fluid showing their presence and importance for fertilization and the embryo development in the oviductal environment [68]. However, further experiments should be done to shed some light into this interaction.
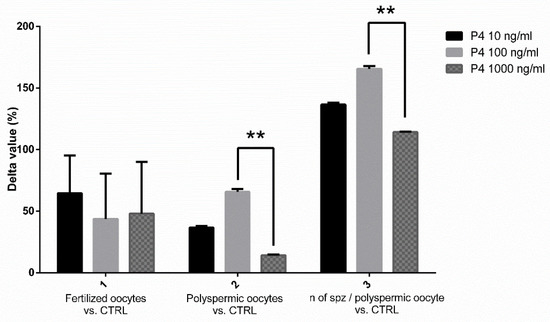
Figure 4.
Graphical representation of the effects of P4 at different concentrations. Results are ex-pressed as the difference in percentage (Δ IVF%) of the fertilization rates, the number of polyspermic oocytes and the number of spermatozoa per polyspermic oocyte, comparing the groups of spermatozoa capacitated in the presence of SOEC treated with P4 (at 10,100 and 1000 ng/mL) to the control group without P4. The best IVF results were obtained when SOECs were treated with P4 at 100 ng/mL, due to the higher values obtained with this concentration in terms of difference of percentage in the number of polyspermic oocytes and in the number of spermatozoa/polyspermic oocyte. The data are presented as mean of four independent experiments. Data were analyzed using a Tukey’s test ** p < 0.01.
3.4. Pre-Treatment of SOECs with P4 but Not Sperm Capacitation Supplementation Improves IVF Outcomes
Once the most effective P4 concentration was identified, we performed a second set of IVF experiments to compare the IVF outcomes obtained when pre-treating the SOECs with P4 and the performance of the system when the hormone is added directly during sperm capacitation in the presence of SOECs (see the experimental design of Figure 1B).
The new data (illustrated in Figure 5) confirmed the results previously obtained: spermatozoa capacitated with SOEC pre-treated with P4 represented the most favorable condition with respect to the groups where P4 (100 ng/mL) was added during capacitation. From the data displayed in Figure 5, it emerges that the pre-treatment of SOECs showed the best performance due to the greater values obtained in the percentage of polyspermic oocytes that allow the inference of the sperm’s fertilizing ability [37,63,64,65].
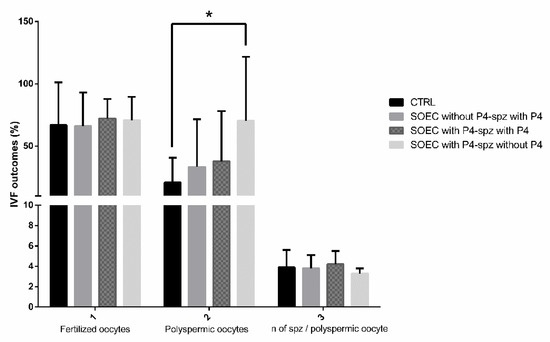
Figure 5.
IVF outcomes: comparison between pre-treatment of SOECs with P4 and addition of P4 during sperm capacitation. The graph shows the effects of co-incubation of sperm during capacitation with P4 (100 ng/mL) and SOECs pre-treated with P4 (100 ng/mL) on IVF outcomes. By analyzing the graph, it arises that the best IVF outcomes are obtained when SOECs are pre-treated with P4 at 100 ng/mL and sperm capacitation is performed without the addition of P4. The data are presented as mean of four independent experiments. Data were analyzed using a Dunnett’s test. * p < 0.05 versus control.
3.5. P4 Induces a Downregulation of the Expression of OVGP1, SPP1 and DMBT1 Genes
Once the most effective P4 concentration for IVF protocol was identified, we evaluated the effect of P4 (100 ng/mL) treatment on gene expression of SOECs, using as a positive control SOECs treated with 17-β-estradiol (E2, 4 ng/mL). We performed and validated a RT-PCR robust system in which the mRNA expression of the three following genes was evaluated: oviductin (OVGP1), osteopontin (SPP1), or deleted in malignant brain tumors 1 (DMBT1).
These three genes are critical in sperm capacitation and fertilization in vitro, since previous studies have shown that their increased expression has been associated with the increase of the rate of monospermic fertilization in swine, through different mechanisms, thus resulting in an increase of normal fertilization in vitro [69]. Other specific effects of each gene on reproduction are reported below.
The OVGP1 gene encodes for the oviduct specific glycoprotein (OVGP1, also called oviductin) [70], which is produced by the non-ciliated epithelial cells of the oviduct and secreted into the lumen [70,71]. OVGP1 has multiple important roles deeply related to fertilization [71,72] conferring by association a higher resistance to the zona pellucida (ZP) before fertilization [69,71]. Moreover, some positive effects have been conferred to the protein during the process of sperm capacitation in terms of viability, motility and fertilization ability, [72], sperm-egg binding and early embryo development [73].
The SPP1 gene codifies for osteopontin (OPN), a phosphoprotein that was first found in the mineralized matrix of bovine bones [69,74] and subsequentially identified in various tissues, including male and female reproductive tracts [74,75]. OPN is involved in immune regulation, matrix communication, cell adhesion, cell migration and bone homeostasis [76]. In the reproductive field, it may play a role in sperm-egg interaction. Indeed, OPN may be able to bind the integrins of the oocytes by its GRGDS sequence, reducing the likelihood of sperm binding [69,74].
Finally, the DMBT1 gene encodes for deleted in malignant brain tumors 1 protein (DMBT1). DMBT1 is a multifunctional glycoprotein present in epithelial cells, mucosal surfaces and the skin [77]. It has been identified as playing several roles in the innate immune defense system against pathogens, homeostasis, inflammation and tumor suppression [77,78,79]. In reproductive organs, DMBT1 has been detected in the oviduct of sus scrofa and human females, and is related to the ability of sperm binding and probably involved in the homeostasis of the epithelium covering the female tract [80,81,82].
Previous studies have shown that steroid hormones might affect the gene expression of the oviductal epithelium both in vitro and ex vivo [38,83,84]. Our results show a consistent downregulation of OVGP1, SPP1 and DMBT1 genes in SOECs pre-treated with P4, as well as a significant higher expression of these genes in SOECs treated with E2 (Figure 6) in comparison to the control. These data confirm that OVGP1 and DMBT1 gene expression is estrogen-dependent [69]. Moreover, our findings demonstrated that P4 administration downregulated the expression of the SPP1 gene, while estradiol also increased the expression of this gene (Figure 6B). Previous works reported a regulation of SPP1 gene expression in a species-specific manner. For instance, in bovine species the gene expression was not correlated to the estrous cycle phases [75], while in other small mammals as the mouse the expression was highest in estrus and lowest in diestrus, indicating that it may be regulated by E2 [85].
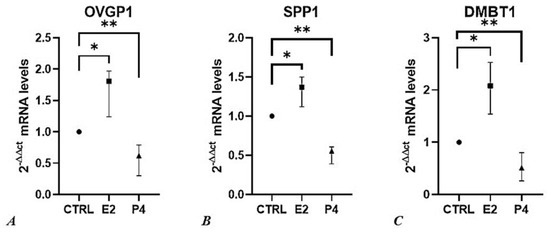
Figure 6.
The effect of progesterone and estradiol on relative gene expression (2−ΔΔCt) of (A) OVGP1, (B) SPP1, and (C) DMBT1. Results are represented as median value with 95 % confidence interval. Data were analyzed using the Kruskal-Wallis test. For each qPCR analysis, each sample was performed in triplicate, and values were normalized to the GAPDH endogenous reference gene. P4, Progesterone; E2, Estradiol; ** = p < 0.001, * = p < 0.01.
The higher percentage of polyspermic oocytes observed as a consequence of the administration of P4 100 ng/mL (Figure 5) is fully concordant with the decrease in mRNA expression of OVGP1, SPP1 and DMBT1, in the same experimental setting.
In line with these results, a previous study demonstrated that increased levels of serum progesterone induce an evident inhibitory effect on OVGP1 mRNA expression in oviductal mucosal tissue from fertile women [86].
Quantitative RT-PCR data show changes in the bioactivity of SOECs treated with P4 with respect to the control group. This confirmed an increase in the dynamism of our system compared to the classic IVF protocols, while partly explaining the increased sperm fertilizing ability. Further proteomic analyzes are needed to evaluate the real presence of the proteins of interest.
4. Conclusions
In conclusion, despite its limitations, our study shed light into the functional link between sperm capacitation and two important modulators of sperm physiology, SOEC and P4. It clearly demonstrates that the incubation of male gametes in the presence of SOECs pre-treated with P4 (100 ng/mL) enhances their fertilizing ability.
These findings could be very important for two reasons. First, they suggest the existence of new mechanisms that can drive capacitation, with the implication in the understanding of the physiology and the pathology of that process. They could then set the basis for the design of innovative approaches to promote capacitation in a dynamic in vitro environment, not only for human ARTs but also for farm animals’ embryo production.
Extending our understanding on the involvement of the main modulators of sperm capacitation in the oviductal environment as with the oviductal epithelial cells and progesterone remains essential in order to design new strategies not only for human ARTs but also for farm animals’ embryo production.
Author Contributions
C.C., A.T. and N.B. conceived the study; C.C., A.T., M.R.-S. and N.B. prepared the original draft; L.V. realized the confocal microscopy experiments, image acquisition and figures; C.C., A.T. and G.C., realized immunocytochemistry and IVF experiments; F.M. and A.C. realized and analyzed gene expression experiments; N.B. and B.B. supervised and funded the research. C.C., F.M., A.T., M.R.-S., A.C., G.C., S.M., L.V., I.T., N.B. and B.B. were involved in interpretation of data and in approval of the final version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mascarenhas, M.N.; Flaxman, S.R.; Boerma, T.; Vanderpoel, S.; Stevens, G.A. National, Regional, and Global Trends in Infertility Prevalence Since 1990: A Systematic Analysis of 277 Health Surveys. PLoS Med. 2012, 9, e1001356. [Google Scholar] [CrossRef] [PubMed]
- Boivin, J.; Bunting, L.; Collins, J.A.; Nygren, K.G. International estimates of infertility prevalence and treatment-seeking: Potential need and demand for infertility medical care. Hum. Reprod. 2007, 22, 1506–1512. [Google Scholar] [CrossRef] [PubMed]
- Ardon, F.; Markello, R.D.; Hu, L.; Deutsch, Z.I.; Tung, C.-K.; Wu, M.; Suarez, S.S. Dynamics of Bovine Sperm Interaction with Epithelium Differ Between Oviductal Isthmus and Ampulla. Biol. Reprod. 2016, 95, 90. [Google Scholar] [CrossRef] [PubMed]
- Suarez, S.S. Mammalian sperm interactions with the female reproductive tract. Cell Tissue Res. 2016, 363, 185–194. [Google Scholar] [CrossRef]
- Tienthai, P. The porcine sperm reservoir in relation to the function of hyaluronan. J. Reprod. Dev. 2015, 61, 245–250. [Google Scholar] [CrossRef][Green Version]
- Ellington, J.E.; Samper, J.C.; Jones, A.E.; Oliver, S.A.; Burnett, K.M.; Wright, R.W. In vitro interactions of cryopreserved stallion spermatozoa and oviduct (uterine tube) epithelial cells or their secretory products. Anim. Reprod. Sci. 1999, 56, 51–65. [Google Scholar] [CrossRef]
- Yeste, M.; Lloyd, R.E.; Badia, E.; Briz, M.; Bonet, S.; Holt, W.V. Direct contact between boar spermatozoa and porcine oviductal epithelial cell (OEC) cultures is needed for optimal sperm survival in vitro. Anim. Reprod. Sci. 2009, 113, 263–278. [Google Scholar] [CrossRef]
- Tuckey, R.C. Progesterone synthesis by the human placenta. Placenta 2005, 26, 273–281. [Google Scholar] [CrossRef]
- Miller, W.L.; Bose, H.S. Early steps in steroidogenesis: Intracellular cholesterol trafficking. J. Lipid Res. 2011, 52, 2111–2135. [Google Scholar] [CrossRef]
- Taraborrelli, S. Physiology, production and action of progesterone. Acta Obstet. Gynecol. Scand. 2015, 94, 8–16. [Google Scholar] [CrossRef]
- Einer-Jensen, N.; Hunter, R.H.F. Counter-current transfer in reproductive biology. Reproduction 2005, 129, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Fujinoki, M.; Takei, G.L.; Kon, H. Non-genomic regulation and disruption of spermatozoal in vitro hyperactivation by oviductal hormones. J. Physiol. Sci. 2016, 66, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Lishko, P.V.; Botchkina, I.L.; Kirichok, Y. Progesterone activates the principal Ca2+ channel of human sperm. Nature 2011, 471, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Teves, M.E.; Barbano, F.; Guidobaldi, H.A.; Sanchez, R.; Miska, W.; Giojalas, L.C. Progesterone at the picomolar range is a chemoattractant for mammalian spermatozoa. Fertil. Steril. 2006, 86, 745–749. [Google Scholar] [CrossRef]
- Guidobaldi, H.A.; Teves, M.E.; Uñates, D.R.; Anastasía, A.; Giojalas, L.C. Progesterone from the Cumulus Cells Is the Sperm Chemoattractant Secreted by the Rabbit Oocyte Cumulus Complex. PLoS ONE 2008, 3, e3040. [Google Scholar] [CrossRef] [PubMed]
- Bronson, R.A.; Peresleni, T.; Golightly, M. Progesterone promotes the acrosome reaction in capacitated human spermatozoa as judged by flow cytometry and CD46 staining. Mol. Hum. Reprod. 1999, 5, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Baldi, E.; Luconi, M.; Muratori, M.; Marchiani, S.; Tamburrino, L.; Forti, G. Nongenomic activation of spermatozoa by steroid hormones: Facts and fictions. Mol. Cell. Endocrinol. 2009, 308, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Foresta, C.; Rossato, M.; Mioni, R.; Zorzi, M. Progesterone induces capacitation in human spermatozoa. Andrologia 2009, 24, 33–35. [Google Scholar] [CrossRef]
- López-Torres, A.S.; Chirinos, M. Modulation of Human Sperm Capacitation by Progesterone, Estradiol, and Luteinizing Hormone. Reprod. Sci. 2017, 24, 193–201. [Google Scholar] [CrossRef]
- Chen, S.; Einspanier, R.; Schoen, J. In vitro mimicking of estrous cycle stages in porcine oviduct epithelium cells: Estradiol and progesterone regulate differentiation, gene expression, and cellular function. Biol. Reprod. 2013, 89, 54. [Google Scholar] [CrossRef]
- Pérez-Cerezales, S.; Ramos-Ibeas, P.; Acuna, O.S.; Avilés, M.; Coy, P.; Rizos, D.; Gutieŕrez-Adán, A. The oviduct: From sperm selection to the epigenetic landscape of the embryo. Biol. Reprod. 2018, 98, 262–276. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hagen, D.E.; Ji, T.; Bakhtiarizadeh, M.R.; Frederic, W.M.; Traxler, E.M.; Kalish, J.M.; Rivera, R.M. Altered microRNA expression profiles in large offspring syndrome and Beckwith-Wiedemann syndrome. Epigenetics 2019, 14, 850–876. [Google Scholar] [CrossRef] [PubMed]
- Grace, K.S.; Sinclair, K.D. Assisted reproductive technology, epigenetics, and long-term health: A developmental time bomb still ticking. Semin. Reprod. Med. 2009, 27, 409–416. [Google Scholar] [CrossRef] [PubMed]
- La Rovere, M.; Franzago, M.; Stuppia, L. Epigenetics and Neurological Disorders in ART. Int. J. Mol. Sci. 2019, 20, 4169. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, M.; Katalinic, A.; Groß, S.; Sutcliffe, A.; Varon, R.; Horsthemke, B. Increased prevalence of imprinting defects in patients with Angelman syndrome born to subfertile couples. J. Med. Genet. 2005, 42, 289–291. [Google Scholar] [CrossRef] [PubMed]
- Eroglu, A.; Layman, L.C. Role of ART in Imprinting Disorders. Semin. Reprod. Med. 2007, 83, 255–262. [Google Scholar] [CrossRef][Green Version]
- Soriano-Úbeda, C.; Avilés-López, K.; García-Vázquez, F.A.; Romero-Aguirregomezcorta, J.; Matás, C. Epididymal and ejaculated sperm functionality is regulated differently by periovulatory oviductal fluid in pigs. Andrology 2021, 9, 426–439. [Google Scholar] [CrossRef]
- París-Oller, E.; Navarro-Serna, S.; Soriano-Úbeda, C.; Lopes, J.S.; Matás, C.; Ruiz, S.; Latorre, R.; López-Albors, O.; Romar, R.; Cánovas, S.; et al. Reproductive fluids, used for the in vitro production of pig embryos, result in healthy offspring and avoid aberrant placental expression of PEG3 and LUM. J. Anim. Sci. Biotechnol. 2021, 12, 32. [Google Scholar] [CrossRef]
- Ferraz, M.A.M.M.; Henning, H.H.W.; Costa, P.F.; Malda, J.; Melchels, F.P.; Wubbolts, R.; Stout, T.A.E.; Vos, P.L.A.M.; Gadella, B.M. Improved bovine embryo production in an oviduct-on-a-chip system: Prevention of poly-spermic fertilization and parthenogenic activation. Lab Chip 2017, 17, 905–916. [Google Scholar] [CrossRef]
- Lamy, J.; Corbin, E.; Blache, M.C.; Garanina, A.S.; Uzbekov, R.; Mermillod, P.; Saint-Dizier, M. Steroid hormones regulate sperm–oviduct interactions in the bovine. Reproduction 2017, 154, 497–508. [Google Scholar] [CrossRef]
- Ramal-Sanchez, M.; Bernabo, N.; Tsikis, G.; Blache, M.C.; Labas, V.; Druart, X.; Mermillod, P.; Saint-Dizier, M. Progesterone induces sperm release from oviductal epithelial cells by modifying sperm proteomics, lipidomics and membrane fluidity. Mol. Cell. Endocrinol. 2020, 504, 110723. [Google Scholar] [CrossRef] [PubMed]
- Romero-Aguirregomezcorta, J.; Cronin, S.; Donnellan, E.; Fair, S. Progesterone induces the release of bull spermatozoa from oviductal epithelial cells. Reprod. Fertil. Dev. 2019, 31, 1463–1472. [Google Scholar] [CrossRef] [PubMed]
- Maccarrone, M.; Barboni, B.; Paradisi, A.; Bernabò, N.; Gasperi, V.; Pistilli, M.G.; Fezza, F.; Lucidi, P.; Mattioli, M. Characterization of the endocannabinoid system in boar spermatozoa and implications for sperm capacitation and acrosome reaction. J. Cell Sci. 2005, 118, 4393–4404. [Google Scholar] [CrossRef] [PubMed]
- Bernabò, N.; Machado-simoes, J.; Valbonetti, L.; Ramal-sanchez, M.; Capacchietti, G.; Fontana, A.; Zappacosta, R.; Palestini, P.; Botto, L.; Marchisio, M.; et al. Graphene Oxide increases mammalian spermatozoa fertilizing ability by extracting cholesterol from their membranes and promoting capacitation. Sci. Rep. 2019, 9, 8155. [Google Scholar] [CrossRef]
- Bernabò, N.; Fontana, A.; Sanchez, M.R.; Valbonetti, L.; Capacchietti, G.; Zappacosta, R.; Greco, L.; Marchisio, M.; Lanuti, P.; Ercolino, E.; et al. Graphene oxide affects in vitro fertilization outcome by interacting with sperm membrane in an animal model. Carbon N. Y. 2018, 129, 428–437. [Google Scholar] [CrossRef]
- Bernabò, N.; Tettamanti, E.; Pistilli, M.G.; Nardinocchi, D.; Berardinelli, P.; Mattioli, M.; Barboni, B. Effects of 50 Hz extremely low frequency magnetic field on the morphology and function of boar spermatozoa capacitated in vitro. Theriogenology 2007, 67, 801–815. [Google Scholar] [CrossRef]
- Abeydeera, L.R.; Day, B.N. Fertilization and subsequent development in vitro of pig oocytes inseminated in a modified Tris-buffered medium with frozen-thawed ejaculated spermatozoa. Biol. Reprod. 1997, 57, 729–734. [Google Scholar] [CrossRef]
- Chen, S.; Einspanier, R.; Schoen, J. Long-term culture of primary porcine oviduct epithelial cells: Validation of a comprehensive invitro model for reproductive science. Theriogenology 2013, 80, 862–869. [Google Scholar] [CrossRef]
- Thornton, B.; Basu, C. Real-time PCR (qPCR) primer design using free online software. Biochem. Mol. Biol. Educ. 2011, 39, 145–154. [Google Scholar] [CrossRef]
- Bernabò, N.; Valbonetti, L.; Raspa, M.; Fontana, A.; Palestini, P.; Botto, L.; Paoletti, R.; Fray, M.; Allen, S.; Machado-Simoes, J.; et al. Graphene Oxide Improves in vitro Fertilization in Mice with No Impact on Embryo Development and Preserves the Membrane Microdomains Architecture. Front. Bioeng. Biotechnol. 2020, 8, 629. [Google Scholar] [CrossRef]
- Oraiopoulou, M.E.; Tzamali, E.; Tzedakis, G.; Vakis, A.; Papamatheakis, J.; Sakkalis, V. In Vitro/In silico study on the role of doubling time heterogeneity among primary glioblastoma cell lines. Biomed Res. Int. 2017, 2017, 8569328. [Google Scholar] [CrossRef] [PubMed]
- Korzyńska, A.; Zychowicz, M. A method of estimation of the cell doubling time on basis of the cell culture monitoring data. Biocybern. Biomed. Eng. 2008, 28, 75–82. [Google Scholar]
- Lindström, H.J.G.; Friedman, R. Inferring time-dependent population growth rates in cell cultures undergoing adaptation. BMC Bioinform. 2020, 21, 583. [Google Scholar] [CrossRef] [PubMed]
- Tzur, A.; Kafri, R.; LeBleu, V.S.; Lahav, G.; Kirschner, M.W. Cell Growth and Size Homeostasis in Proliferating Animal Cells. Science 2009, 325, 167. [Google Scholar] [CrossRef]
- Sun, B.; Fang, Y.; Li, Z.; Chen, Z.; Xiang, J. Role of cellular cytoskeleton in epithelial-mesenchymal transition process during cancer progression. Biomed. Rep. 2015, 3, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Tojkander, S.; Gateva, G.; Lappalainen, P. Actin stress fibers-Assembly, dynamics and biological roles. J. Cell Sci. 2012, 125, 1855–1864. [Google Scholar] [CrossRef] [PubMed]
- Svitkina, T. The Actin Cytoskeleton and Actin-Based Motility. Cold Spring Harb. Perspect. Biol. 2018, 10, a018267. [Google Scholar] [CrossRef]
- Goodson, H.V.; Jonasson, E.M. Microtubules and Microtubule-Associated Proteins. Cold Spring Harb. Perspect. Biol. 2018, 10, a022608. [Google Scholar] [CrossRef]
- Etienne-Manneville, S. Actin and Microtubules in Cell Motility: Which One is in Control? Traffic 2004, 5, 470–477. [Google Scholar] [CrossRef]
- Parker, A.L.; Kavallaris, M.; McCarroll, J.A. Microtubules and their role in cellular stress in cancer. Front. Oncol. 2014, 4, 1–19. [Google Scholar] [CrossRef]
- Muñoz-Lasso, D.C.; Romá-Mateo, C.; Pallardó, F.V.; Gonzalez-Cabo, P. Much More Than a Scaffold: Cytoskeletal Proteins in Neurological Disorders. Cells 2020, 9, 358. [Google Scholar] [CrossRef] [PubMed]
- Magliocca, V.; Petrini, S.; Franchin, T.; Borghi, R.; Niceforo, A.; Abbaszadeh, Z.; Bertini, E.; Compagnucci, C. Identifying the dynamics of actin and tubulin polymerization in iPSCs and in iPSC-derived neurons. Oncotarget 2017, 8, 111096–111109. [Google Scholar] [CrossRef] [PubMed]
- Suarez, S.S.; Pacey, A.A. Sperm transport in the female reproductive tract. Hum. Reprod. Update 2006, 12, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.J. Review: The epic journey of sperm through the female reproductive tract. Animal 2018, 12, s110–s120. [Google Scholar] [CrossRef] [PubMed]
- Reshi, Q.U.A.; Viil, J.; Ord, J.; Lättekivi, F.; Godakumara, K.; Hasan, M.M.; Nõmm, M.; Jääger, K.; Velthut-Meikas, A.; Jaakma, Ü.; et al. Spermatozoa induce transcriptomic alterations in bovine oviductal epithelial cells prior to initial contact. J. Cell Commun. Signal. 2020, 14, 439–451. [Google Scholar] [CrossRef]
- Mahé, C.; Zlotkowska, A.M.; Reynaud, K.; Tsikis, G.; Mermillod, P.; Druart, X.; Schoen, J.; Saint-Dizier, M. Sperm migration, selection, survival, and fertilizing ability in the mammalian oviduct. Biol. Reprod. 2021, 105, 317–331. [Google Scholar] [CrossRef]
- Chang, H.; Suarez, S.S. Unexpected flagellar movement patterns and epithelial binding behavior of mouse sperm in the oviduct. Biol. Reprod. 2012, 86, 140. [Google Scholar] [CrossRef]
- Gualtieri, R.; Talevi, R. In vitro-cultured bovine oviductal cells bind acrosome-intact sperm and retain this ability upon sperm release. Biol. Reprod. 2000, 62, 1754–1762. [Google Scholar] [CrossRef]
- Fazeli, A.; Elliott, R.M.A.; Duncan, A.E.; Moore, A.; Watson, P.F.; Holt, W.V. In Vitro Maintenance of Boar Sperm Viability by a Soluble Fraction Obtained from Oviductal Apical Plasma Membrance Prepations. Available online: https://pubmed.ncbi.nlm.nih.gov/12683921/ (accessed on 23 November 2021).
- Coy, P.; García-Vázquez, F.A.; Visconti, P.E.; Avilés, M. Roles of the oviduct in mammalian fertilization. Reproduction 2012, 144, 649–660. [Google Scholar] [CrossRef]
- Hunter, R.H.F.; Petersen, H.H.; Greve, T. Ovarian follicular fluid, progesterone and Ca2+ ion influences on sperm release from the Fallopian tube reservoir. Mol. Reprod. Dev. 1999, 54, 283–291. [Google Scholar] [CrossRef]
- Gervasi, M.G.; Osycka-Salut, C.; Sanchez, T.; Alonso, C.A.I.; Llados, C.; Castellano, L.; Franchi, A.M.; Villalõn, M.; Perez-Martinez, S. Sperm Release From the Oviductal Epithelium Depends on Ca(2+) Influx Upon Activation of CB1 and TRPV1 by Anandamide. J. Cell. Biochem. 2016, 117, 320–333. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Saito, Y.; Kagawa, N.; Yang, X. In vitro fertilization and polyspermy in the pig: Factors affecting fertilization rates and cytoskeletal reorganization of the oocyte. Microsc. Res. Tech. 2003, 61, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Abeydeera, L.R.; Funahashi, H.; Kim, N.H.; Day, B.N. Chlortetracycline fluorescence patterns and in vitro fertilisation of frozen-thawed boar spermatozoa incubated under various bicarbonate concentrations. Zygote 1997, 5, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Hunter, R.H.F.; Nichol, R. Capacitation potential of the fallopian tube: A study involving surgical insemination and the subsequent incidence of polyspermy. Gamete Res. 1988, 21, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Almiñana, C.; Tsikis, G.; Labas, V.; Uzbekov, R.; Silveira, J.C.; Bauersachs, S.; Mermillod, P. Deciphering the oviductal extracellular vesicles content across the estrous cycle: Implications for the gametes-oviduct interactions and the environment of the potential embryo. BMC Genom. 2018, 19, 622. [Google Scholar] [CrossRef] [PubMed]
- Franchi, A.; Moreno-Irusta, A.; Domínguez, E.M.; Adre, A.J.; Giojalas, L.C. Extracellular vesicles from oviductal isthmus and ampulla stimulate the induced acrosome reaction and signaling events associated with capacitation in bovine spermatozoa. J. Cell. Biochem. 2019, 121, 2877–2888. [Google Scholar] [CrossRef] [PubMed]
- Asaadi, A.; Dolatabad, N.A.; Atashi, H.; Raes, A.; van Damme, P.; Hoelker, M.; Hendrix, A.; Pascottini, O.B.; Van Soom, A.; Kafi, M.; et al. Extracellular vesicles from follicular and ampullary fluid isolated by density gradient ultracentrifugation improve bovine embryo development and quality. Int. J. Mol. Sci. 2021, 22, 578. [Google Scholar] [CrossRef]
- Coy, P.; Yanagimachi, R. The Common and Species-Specific Roles of Oviductal Proteins in Mammalian Fertilization and Embryo Development. Bioscience 2015, 65, 973–984. [Google Scholar] [CrossRef]
- Choudhary, S.; Janjanam, J.; Kumar, S.; Kaushik, J.K.; Mohanty, A.K. Structural and functional characterization of buffalo oviduct-specific glycoprotein (OVGP1) expressed during estrous cycle. Biosci. Rep. 2019, 39, BSR20191501. [Google Scholar] [CrossRef]
- Algarra, B.; Han, L.; Soriano-Úbeda, C.; Avilés, M.; Coy, P.; Jovine, L.; Jiménez-Movilla, M. The C-terminal region of OVGP1 remodels the zona pellucida and modifies fertility parameters. Sci. Rep. 2016, 6, 32556. [Google Scholar] [CrossRef]
- Lamy, J.; Nogues, P.; Combes-Soia, L.; Tsikis, G.; Labas, V.; Mermillod, P.; Druart, X.; Saint-Dizier, M. Identification by proteomics of oviductal sperm-interacting proteins. Reproduction 2018, 155, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhao, Y.; Yang, X.; Kan, F.W.K. Recombinant hamster oviductin is biologically active and exerts positive effects on sperm functions and sperm-oocyte binding. PLoS ONE 2015, 10, e0123003. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, R.F.; Wolinetz, C.D.; Killian, G.J. Influence of arginine-glycine-aspartic acid (RGD), integrins (αV and α5) and osteopontin on bovine sperm-egg binding, and fertilization in vitro. Theriogenology 2007, 67, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Gabler, C.; Chapman, D.A.; Killian, G.J. Expression and presence of osteopontin and integrins in the bovine oviduct during the oestrous cycle. Reproduction 2003, 126, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.X.; Denhardt, D.T. Osteopontin: Role in immune regulation and stress responses. Cytokine Growth Factor Rev. 2008, 19, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Madsen, J.; Sorensen, G.L.; Nielsen, O.; Tornøe, I.; Thim, L.; Fenger, C.; Mollenhauer, J.; Holmskov, U. A Variant Form of the Human Deleted in Malignant Brain Tumor 1 (DMBT1) Gene Shows Increased Expression in Inflammatory Bowel Diseases and Interacts with Dimeric Trefoil Factor 3 (TFF3). PLoS ONE 2013, 8, e64441. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Kim, B.C.; Yeo, H.Y.; Kim, K.H.; Yoo, B.C.; Park, J.W.; Chang, H.J. Deleted in malignant brain tumor 1 is a novel prognostic marker in colorectal cancer. Oncol. Rep. 2018, 39, 2279–2287. [Google Scholar] [CrossRef] [PubMed]
- Müller, H.; Nagel, C.; Weiss, C.; Mollenhauer, J.; Poeschl, J. Deleted in malignant brain tumors 1 (DMBT1) elicits increased VEGF and decreased IL-6 production in type II lung epithelial cells. BMC Pulm. Med. 2015, 15, 32. [Google Scholar] [CrossRef]
- Neville, M.C.; Zhang, P. Lactoferrin secretion into milk: Comparison between ruminant, murine, and human milk. J. Anim. Sci. 2000, 78, 26. [Google Scholar] [CrossRef]
- Ambruosi, B.; Accogli, G.; Douet, C.; Canepa, S.; Pascal, G.; Monget, P.; Moros, C.; Holmskov, U.; Mollenhauer, J.; Robbe-Masselot, C.; et al. Deleted in malignant brain tumor 1 is secreted in the oviduct and involved in the mechanism of fertilization in equine and porcine species. Reproduction 2013, 146, 119–133. [Google Scholar] [CrossRef]
- Roldán, M.L.; Marini, P.E. First evidence of the interaction between deleted in malignant brain tumor 1 and galectin-3 in the mammalian oviduct. Histochem. Cell Biol. 2014, 141, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Saint-Dizier, M.; Schoen, J.; Chen, S.; Banliat, C.; Mermillod, P. Composing the early embryonic microenvironment: Physiology and regulation of oviductal secretions. Int. J. Mol. Sci. 2020, 21, 223. [Google Scholar] [CrossRef] [PubMed]
- Valero, A.; Roldán, M.L.; Ruiz, M.F.; Teijeiro, J.M.; Marquez, S.B.; Marini, P.E. Deleted in Malignant Brain Tumor 1 (DMBT1) Expression Pattern in Normal Cervix and at Different Stages of Squamous Intraepithelial Lesions. Open Biomark. J. 2018, 8, 1–8. [Google Scholar] [CrossRef]
- Liu, Q.; Xie, Q.Z.; Zhou, Y.; Yang, J. Osteopontin is expressed in the oviduct and promotes fertilization and preimplantation embryo development of mouse. Zygote 2014, 760. [Google Scholar] [CrossRef] [PubMed]
- Briton-Jones, C.; Hung Lok, I.; Mo Yuen, P.; Tak Yu Chiu, T.; Ping Cheung, L.; Haines, C. Regulation of human oviductin mRNA expression in vivo. Fertil. Steril. 2001, 75, 942–946. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).