4.1. Carcass Yields
It was observed that the animals harvested in the summer season (291.5 ± 11.65 kg) had a lower undressed weight than the animals harvested in the winter season (324.4 ± 5.55 kg;
Table 1). The differences in undressed carcass weight were reflected in the lower dressed carcass yields (warm and cold) observed for the summer-harvested animals (warm = 168.9 ± 5.79 kg; cold = 164.5 ± 5.53 kg) compared to the winter-harvested animals (warm = 193.1 ± 3.95 kg; cold = 188.3 ± 4.03 kg). Information on the carcass weight of the Plains zebra is limited to two studies [
21,
22]. Both these studies reported lower carcass yields compared to the current study, which may potentially be attributed to the dry season the animals in these two studies were harvested in. The lower carcass yields reported for the summer-harvested animals in the current study could be due to seasonal differences in the diet, harvest location and forage behaviour, altering the plain of nutrition and as a result the body composition. In the Western Cape Province, the rainy season is in the winter between May and August and the dry summer months between November and February. During the wet season, the availability of forage increases in biomass and consists of leafy grass with high nutrient concentrations. In contrast, the biomass during the dry season is reduced and the Plains zebras are dependent on fibrous grass species with low nutritional quality [
23]. Additionally, the Plains zebras harvested in the winter season, in this study, had access to a supplement lick during the dry months to restore depleted minerals. Furthermore, the Plains zebra also tends to consume more forage in the rainy season, than in the dry season, due to natural grazing being more abundant, leading to more grass bites per step taken [
24]. This results in an increase of fat deposition in the rainy season and as observed in this study, the undressed and dressed carcass weights were heavier for the Plains zebras harvested in the winter season. Visually, it was noted in this study that the winter-harvested Plains zebra carcasses were characterised by a higher degree of subcutaneous fat deposition, when compared to that of the summer-harvested Plains zebras. However, it needs to be kept in mind that the difference in level of subcutaneous fat deposition may potentially also be ascribed to the mixed age composition of the summer-harvested group (i.e., sub-adults and adults), whereas the winter-harvested group consisted of only adult animals
Historically, meat-producing horses were slaughtered at the end of their working career, and only recently this practice was changed with foals being slaughtered for consumption [
25]. The slaughter of meat-producing horses is thus based on age which have a beneficial influence on carcass characteristics, with season not considered as these horses are provided with artificial feed year-round irrespective of the location or season [
26]. In this study, to investigate the effect of age on undressed, warm and cold carcass weights, and dressing percentage for Plains zebra, an exponential regression analysis was carried out. A low coefficient of determination (R
2 ≤ 0.24) was reported for all the carcass factors, indicating that less than 24% of the variation in carcass yields can be ascribed to the influence of animal age. The low R
2 value potentially indicate that the summer-harvested animals already achieved maximum growth and thus were in the plateau phase on a typical sigmoidal curve at the point of harvest. Mentis [
20] found that free-roaming Plains zebra reach their mature body weight at the age of 36 months. The youngest animal in this study was 24 months, and the advanced growth may potentially be ascribed to the fact that these animals were part of the Quagga project, i.e., would have received additional feed when required and thus not have been subjected to the challenges of seasonal forage availability and quality that free-roaming Plains zebras experience. Thus, the differences found between the two groups in this study are most likely due to the effect of the harvesting season on subcutaneous fat deposition and, as a result, body composition.
Little information is available on the meat production potential of Plains zebra, therefore findings in this study will be compared to related information about commercial horse meat breeds and/or donkey breeds. With a focus on specialized horse breeds for meat production, most of the breeds had higher slaughter and carcass weights than the Plains zebra. Heavier weights were reported for the Burguete horse breed slaughtered at 16 months (411.3 kg and 275.5 kg, respectively) [
27] and 24 months (395 kg and 258.9 kg, respectively), the Hispano-Bretón (HB) breed at 24 months (406 kg and 275.5 kg, respectively) [
28], and Sanfratellano breed at 18 months (411 kg and 243.75 kg) [
29]. The Haflinger breed also slaughtered at 18 months had a comparable slaughter weight (349.83 kg) and carcass weight (207.83 kg) to the Plains zebras in this study [
29]. In this Plains zebra study, it was noted that there is little correlation between the carcass yields and age; similarly, no significant differences were found between age and carcass weights in slaughter horses between the ages of three and 21 years. The average warm carcass weights of these slaughtered horses were 586.4 kg for geldings, 663.5 kg for mares and 671.5 kg for stallions [
30]. The Plains zebra is heavier than Martina Franca donkeys (101–181 kg; 8–15 months of age) [
31,
32,
33] and domesticated donkeys found in tropical regions in Africa (such as Botswana), Central America and Asia (<150 kg) [
34]. Interestingly the undressed and cold carcass weight of the Plains zebra harvested in the summer season are comparable to Martina Franca × Ragusana male donkeys (285 kg and 154 kg, respectively) whilst those harvested in the winter season to male mules (363 kg and 212 kg, respectively) slaughtered at 16 years of age [
35]. Compared to the meat-producing horse and donkey breeds, the Plains zebras in this study are intermediate in terms of undressed-, warm- and cold carcass weights.
Dressing percentage is an important parameter when the meat production potential of an animal or species is investigated [
36]. In game animals, the dressing percentage is the proportion of the dressed weight relative to the undressed carcass weight. Game animals cannot be fasted before harvesting and therefore may have a higher gut fill than domestic livestock species that are typically fasted in lairage for 24 h before slaughter. Comparable results for the dressing percentage for both the winter (59.5 ± 0.55%) and summer seasons (58.1 ± 0.68%) were found in this study. The difference observed is attributed to the higher carcass yields observed and the higher subcutaneous fat level noted in the winter harvesting group. A lower dressing percentage of 56% for Plains zebra was reported [
21], however, the sample size of the study was very small (
n = 2) and animals were harvested in very dry, semi-arid conditions; these conditions may have contributed to the low carcass yields and dressing percentages reported.
Meat-producing horses typically yield high dressing percentages, when compared to other domestic livestock species. The dressing percentage of the Plains zebras in this study were higher than that reported for the HB × Galician Mountain crossbreed (52.8%) and the Galician Mountain breed (50.3%) [
37], and are comparable to that of the Haflinger (59.6%) and Sanfratellano horse breeds (59.3%) [
29]. However, horse breeds such as the Burguete [
27,
28], HB [
28], Italian Heavy Draft horse breed [
38] and horses slaughtered in Poland [
39] had higher dressing percentages (>63%) than the Plains zebras harvested in this study. The studies reporting high dressing percentages can potentially be ascribed to a low gut fill as a result of the animals subjected to pre-slaughter fasting, which subsequently resulted in higher dressing percentages. Another factor that can possibly influenced the dressing percentage reported for Plains zebra in this study, is the degree of blood loss before weighing; potentially resulting in a higher calculated dressing percentage. The dressing percentage of the Plains zebras harvested in both seasons compared favourably to that of Martina Franca donkeys (49.2–57.5%) found in various studies [
32,
33,
34]. The warm and cold dressing percentage of the Plains zebra also compared favourably to Martina Franca × Ragusana male donkey foals reared intensively (55.2% and 54.3%, respectively) and extensively (53.1% and 51.3%, respectively) [
40].
When comparing the dressing yield of the Plains zebra with other game species commonly found in South Africa, the dressing percentage compared favourably with impala (58.0%) [
41], springbok (56%) [
42], blesbok (50.6–53.7%) [
42,
43] and large-bodied game species such as the greater kudu (58.3%) [
44], gemsbok (54%) [
21], eland (50.8–51%) [
42,
45], black wildebeest (53.19%) [
46], African savannah buffalo (48–53%) [
47] and male giraffe (51.6–59.2%) [
48]. However, higher dressing percentages for male springbok (64.9%), blesbok (62.9%) and impala (63.4%) have been reported [
49]. The differences in dressing percentages can be attributed to seasonal and location differences influencing the body composition of these species [
50].
The dressing percentages of game animals are also comparable with domestic livestock species such as cattle, sheep, and goats. The dressing percentage of the Plains zebra compares favourably to that of Nguni (52.1%), Bonsmara (56.9%) and Aberdeen Angus (53.7%) cattle [
51]. The Plains zebra values in this regard also compared favourably to sheep breeds such as the South African Mutton Merino (SAMM; 41.5%), Dormer (44.2%) [
52], and fat-tailed indigenous Damara (59.9%) and also to goats such as Boer goats and indigenous goats (55.7%) [
53].
4.2. Offal Yields
Game animals produce by-products that are edible and non-edible. Edible by-products include tripe, liver, heart, lungs, kidneys and spleen; and external offal which include the head, tail and feet/lower legs [
54]. Edible offal is marketed by informal suppliers and forms part of the traditional diet in South Africa [
54] with a per capita consumption of 4.7 kg and 4.8 kg for the year of 2009 and 2013, respectively [
1]. Information on the quantity of edible offal of game species is limited and needs to be investigated since edible offal is a low-cost protein and nutrient-rich food source that has the potential to contribute to food security [
54,
55]. Non-edible offal, in game species, such as the skin and horns (as well as the head after removal of the cheek and other meat/muscles) are primary products collected during trophy hunting and are of high economic value in the game industry [
54,
56]. The Plains zebra is a hindgut fermenter and lacks horns, unlike some game species, and should therefore rather be compared with other equine species in terms of head and GIT weights. Information on offal yields in horses and donkeys, however, is limited.
In this study, the highest contributing external offal component to the undressed carcass weight in Plains zebras harvested in both locations, was the skin followed by the head (
Table 3). Presently, low quantities of Plains zebra meat enter the formal market value chain in South Africa due to Plains zebras being primarily hunted by recreational hunters for their skins with the meat seen as a secondary product. However, large numbers of zebras are culled by professional teams for the utilisation of their skin and meat. The meat is typically exported to the European Union as Plains zebras are excluded from the FMD control regulation [
57]. It is important for the industry to note the contribution of the skin and head (as well as the legs under the knee joint with the hooves) to the carcass weight as these body parts remain attached to the carcass during transportation to the processing plants. This enables the marketers to achieve a top price for the skin since the skin on the head is also flayed with that of the body and lower legs. Another reason for keeping the skin on the carcass during transportation and chilling, is to reduce moisture loss since only a few game species have a subcutaneous fat layer to minimise this loss. However, in this study, the head and skin were removed prior to transportation and chilling and the head of the Plains zebras from both locations was weighed without the skin as the skin was flayed according to industry standards. A lower skin weight and contribution to the undressed weight for animals harvested in the winter season (22.288 ± 0.533 kg; 6.9 ± 0.19%) were found than for animals harvested in the summer season (24.046 ± 0.927 kg; 8.3 ± 0.18%). The lower skin weight of animals harvested in the winter than in the summer can potentially be attributed to different skinning teams that were used for each season. A professional skinning team were used for the winter harvest, and a non-professional skinning team were used for the summer harvest, which may have resulted in variations in the subcutaneous fat left on the skin, thus influencing the contribution of the skin to the undressed weight.
The head weight and its contribution to the undressed weight were found to be very similar for the winter (12.606 ± 0.322 kg; 3.9 ± 0.05%) and summer-harvested animals (12.046 ± 0.344 kg; 4.2 ± 0.13%). A weak relationship between animal age and head and skin weight, as evident in the low R
2 value obtained with the exponential regression analysis, is indicative that the animals were mature at the point of harvest, which supports the conclusion for the carcass yields. These findings agree with the observations in male horses slaughtered in México where the skin contributed 7.4% and the head 4.5% to the live weight (277.8 kg) [
58]. The slaughter weight of these horses fell in the range of the undressed weight of the Plains zebras harvested in the summer season (
Table 1). Catalan crossbreed donkeys with a live weight of more than 151 kg were found to have a similar head and lower skin contribution to the empty body weight (live weight−gastrointestinal content) than the Plains zebras in the current study [
59]. The lower skin contribution observed can be postulated to be due to the skin weight not including the skin on the head and legs, as the study did not provide a detailed methodology [
59].
When compared to smaller game species, the proportional weight of the skin of the Plains zebra from both seasons was higher than found for male impala (3.9%) and male fallow deer (6.6%) with an undressed weight of 49.9 kg and 47.4 kg, respectively [
41,
60]. The skin weight and contribution can also be compared to large-bodied game species which will have heavier skins and body weights such as eland bulls (14.3 kg and 6.8%) with an undressed weight of 208.6 kg [
61] and blue wildebeest bulls (13.6–17.9 kg and 7.9–8.4%) weighing 168.8–208.2 kg [
62].
In terms of the internal offal, animals harvested in the winter season (70.760 ± 3.113 kg; 21.8 ± 0.89%) produced heavier or similar yields when compared to animals harvested in the summer season (66.132 ± 3.782 kg; 22.5 ± 0.50%). The GIT had the highest internal weight and contribution to the undressed weights of animals harvested in the winter (60.150 ± 3.300 kg; 18.6 ± 0.97%) and summer season (56.050 ± 3.547 kg; 19.1 ± 0.54%). The lower GIT weight reported for the summer-harvested animals can potentially be attributed to a lower gut fill, since it has been shown by the exponential regression that the effect of age had a weak effect on the GIT proportion (R
2-value = 0.1153;
Table 4). It has been found that in the rainy (winter) season the Plains zebra consumes more forage (by counting the grass bites per step taken) because of the natural grazing being more abundant [
24].
The liver represents a valuable source of vitamin A, vitamin B
1, and nicotinic acid. The liver and kidneys contain higher levels of iron, copper and zinc than skeletal muscle, [
63], making it a valuable low-cost protein source especially for the rural communities of South Africa [
55]. However, recently offal is rather processed into niche-market products that are seen as delicacies in upmarket restaurants [
55]. The liver weight and contribution to total carcass yield in the winter season (3.300 ± 0.130 kg; 1.0 ± 0.04%) were comparable with that found for the summer season (3.320 ± 0.109 kg; 1.2 ± 0.5%). The kidney weights in the winter season (0.740 ± 0.030 kg; 0.3 ±0.01% of carcass weight) were also comparable with that found for the summer season (0.648 ± 0.032 kg; 0.2 ± 0.03%). The contribution of the liver weight to the empty body weight (live weight−gastrointestinal content) in Catalan crossbreed donkeys with a live weight of >151 kg was found to be similar to the Plains zebras when calculated in the same manner [
59].
The liver weight as a proportion to carcass weight is comparable to that of impala (1.3%; [
41]), blesbok (1.3%; [
49]) and blue wildebeest (1.0%; [
62]). When compared to livestock species such as the South Africa Mutton Merino (SAMM; 0.99 kg) and Dormer lambs (1.13 kg) [
52], the Plains zebra has heavier livers. However, the contribution to the undressed weight was similar to SAMM (1.9%) and Dormer (2.0%) sheep breeds [
52]. The proportional weight of the kidneys relative to the live weight are comparable with impala (0.3%; [
41]), fallow deer (0.3%; [
60]), blesbok (0.3%; [
49]) and blue wildebeest (0.2%; [
62]).
4.3. Muscle Yields
The same primal meat cuts used to section beef carcasses are also used for the sectioning of game meat species and meat-horse breeds. The game meat trade however tends to sell muscles whole rather than as meat cuts that are frequently made up out of two or more muscles. In this study, seven muscles including the LTL (loin), SM (topside), BF (silverside), ST (eye of the round), PM (fillet) and two additional representative shoulder muscles, the SS and IS, were removed from the carcass and weighed to determine each muscle’s contribution to the overall meat yield in Plains zebra. To the best of our knowledge, no information is available on the contribution of the respective muscles to the meat yield in Plains zebra.
Recorded weight for all seven muscles for the winter-harvested animals were numerically heavier when compared to the weights recorded for the summer-harvested group (
Table 5). Notable numerical differences between the winter and summer harvest can be observed for the LTL (7.455 ± 0.829 kg and 5.410 ± 0.298 kg, respectively), SM (3.178 ± 0.093 kg and 2.642 ± 0.120 kg, respectively) and BF (5.271 ± 0.188 kg and 4.215 ± 0.142 kg, respectively) muscles. The difference in weight between the two seasons can potentially be attributed to the lower carcass yields observed for the summer-harvested group (
Table 1), due to the real muscle yield being dependent on the weight at slaughter and consequently the carcass weight. The remainder of the seven muscles had almost numerically similar weights and proportions when compared between seasons. Age did not have a strong influence on carcass or organ yields observed for the summer-harvested animals, and therefore an exponential regression analysis was carried for each muscle weight and its calculated proportion to the cold carcass weight. The analysis generated high R
2 values for the LTL (0.6501), BF (0.7466) and ST (0.6431), however, it is more of value to consider the proportional contribution of each muscle to the cold carcass weight. The R
2-values showed a weak increase with the cold carcass weight with 29.1% (highest R
2-value) of the variation in the LTL and 0.4% (lowest R
2-value) of the variation in the ST explained by the exponential relationship with the cold carcass weight. The results indicates that the proportional contribution of the muscles is relatively similar irrespective of the lower or higher carcass weights observed in this study.
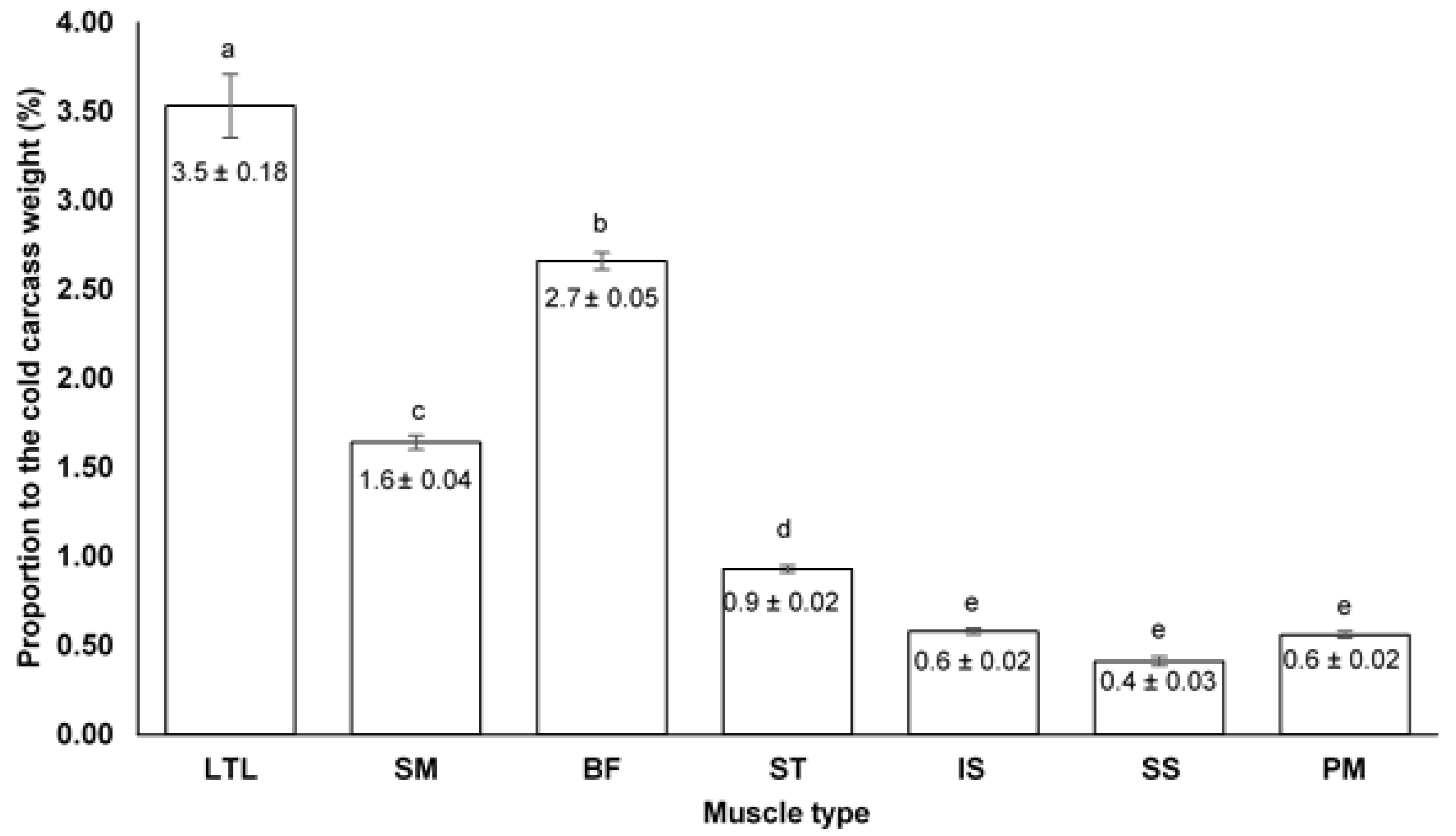
The mean proportion (%) of each individual muscle to the mean cold carcass weight of the 20 Plains zebra stallions were pooled and presented in
Figure 1. Significant differences were found between the hindquarter muscles; the LTL (3.5%), SM (1.6%), BF (2.7%) and ST (0.9%) which also significantly differed from the smaller sized muscles, the IS (0.6%), SS (0.4%) and PM (0.6%). The differences found between muscles in terms of contribution (and weight) can be attributed to their anatomical location and function. The LTL was found to be the highest contributing muscle which is expected since the LTL is an epaxial muscle with many bundle fibres extending over the vertebral column from the forequarter to the hindquarter [
64]. The BF was the second-highest contributing muscle followed by the SM and then the ST. These muscles are the proximal muscles on the pelvic limb and are larger than the muscles on the shoulder joint due to their locomotive and hip extension functions [
64,
65]. The PM muscle is a small muscle involved in the flexion of the hip and is one of the most tender muscles on a carcass due to its low functional activity resulting in small fibre diameters and less connective tissue. The PM muscle was found to have a similar contribution as the two stabilizing shoulder muscles; the IS and the SS which are respectively involved in the flexion and extension of the shoulder [
64].
The muscle yields of the Plains zebra can be compared to game species and various horse breeds. However, data on individual muscle yields in various horse breeds is limited, since yields are represented as primal meat cuts consisting out of two or more muscles and therefore cannot be compared. As most meat-producing horse breeds are larger at slaughter, one can speculate that muscles represented in primal meat cuts in the Plains zebra will be smaller than the corresponding muscles in horses, and bigger than the corresponding muscles in donkeys. The LTL, SM, BF and ST in the Plains zebra were found to have a lower contribution to the total carcass weight, when compared to findings reported for blue wildebeest (4.8–5.0%, 3.8–3.9%, 4.8–5.0% and 1.5%, respectively) [
62] and fallow deer (7.5%, 5.2%, 6.1% and 1.6%, respectively) [
60]. The LTL muscle had a higher contribution, the SM a lower contribution, the BF a higher contribution and the ST were found to have a similar contribution to the cold carcass weight when compared to eland (2.6%, 2.2%, 2.4% and 0.9%) [
45]. Higher percentages for the IS and SS muscle were reported for fallow deer (1.3% and 1.1%, respectively) [
60], eland (0.7% and 0.7%, respectively) [
45] and blue wildebeest (1.3–1.4% and 1.1–1.2%, respectively) [
62]; results that may be related to a more active behaviour, e.g., like jumping more frequently by fallow deer and eland.



{kind=link}