
Genome-Wide Association Study Identifies the Crucial Candidate Genes for Teat Number in Crossbred Commercial Pigs
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animal Samples and Genotyping and Quality Control
2.3. Population Structure
2.4. Genome-Wide Association Study
2.5. Functional Candidate Genes Search
3. Results
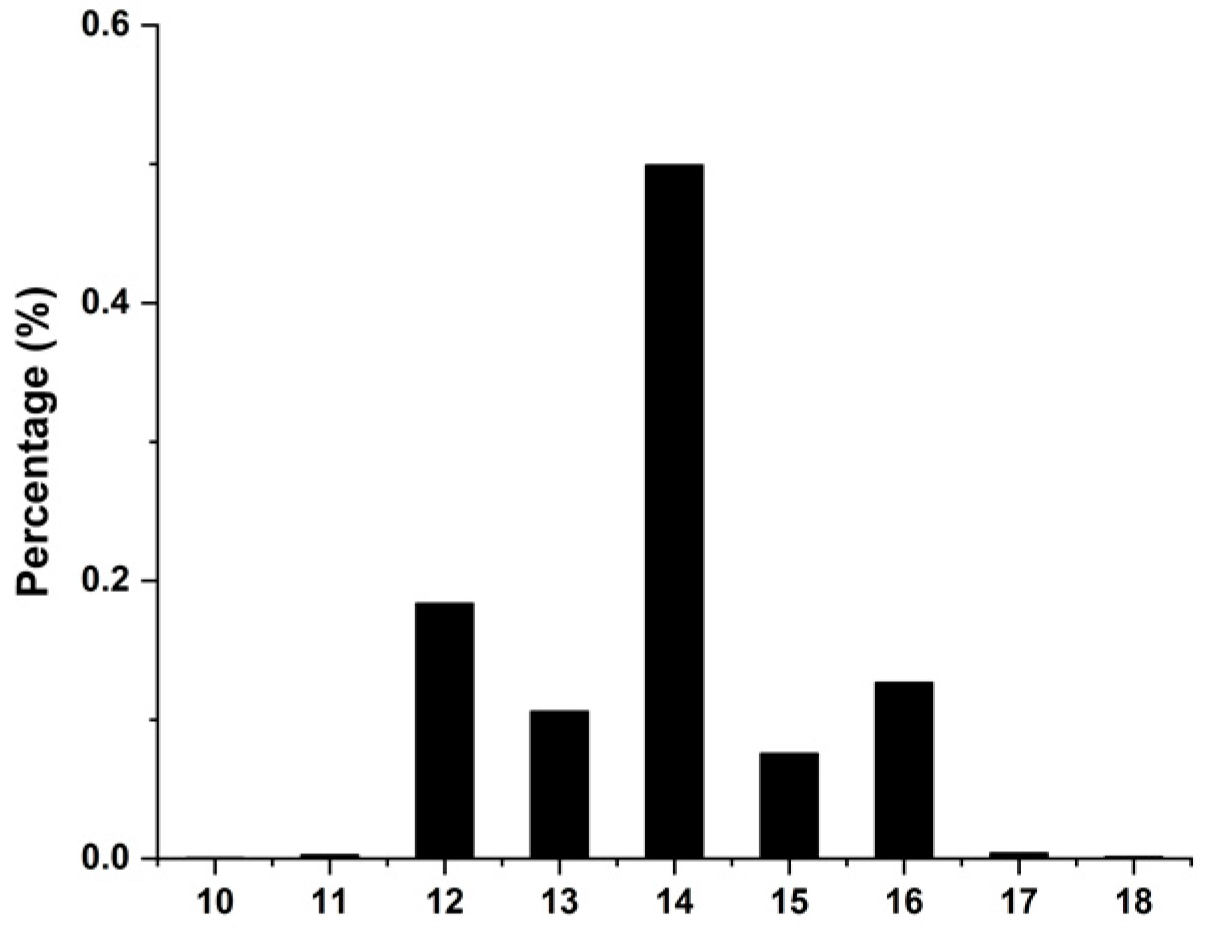
3.1. Phenotype Statistic
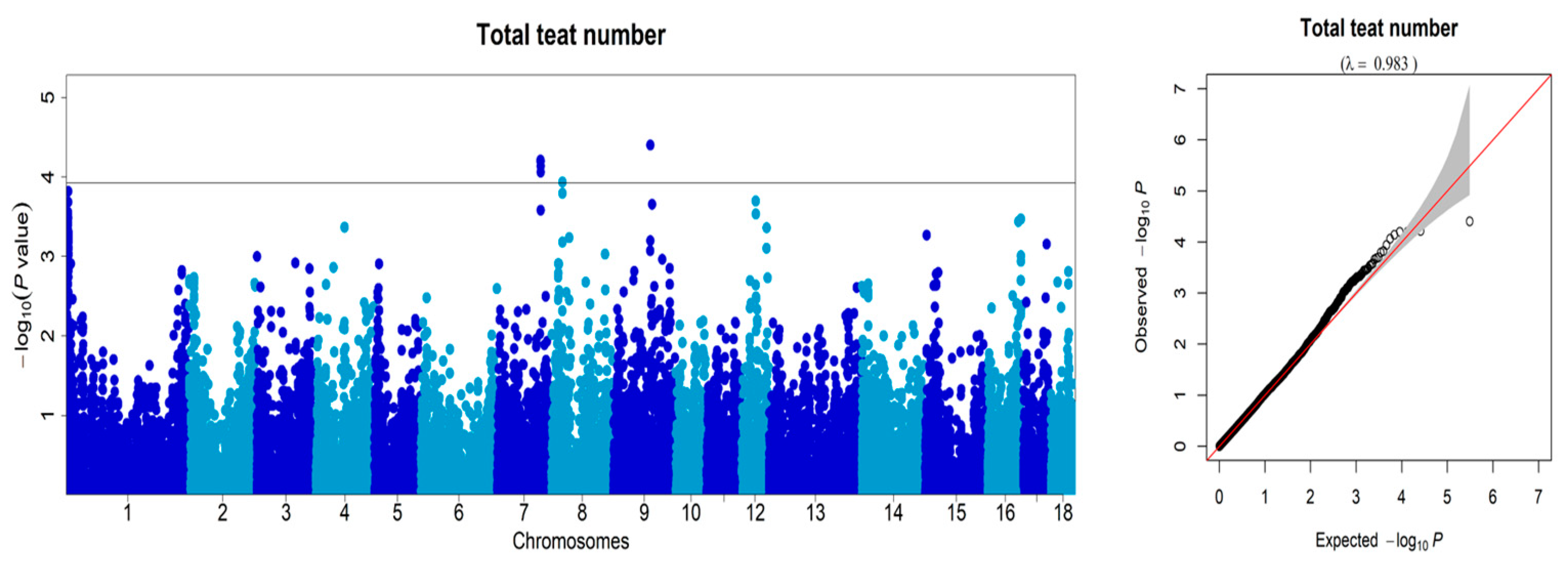
3.2. Genome-Wide Association Studies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, J.; Chen, C.; Yang, B.; Guo, Y.; Ai, H.; Ren, J.; Peng, Z.; Tu, Z.; Yang, X.; Meng, Q.; et al. A systems genetics study of swine illustrates mechanisms underlying human phenotypic traits. BMC Genom. 2015, 16, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohrer, G.A.; Nonneman, D.J. Genetic analysis of teat number in pigs reveals some developmental pathways independent of vertebra number and several loci which only affect a specific side. Genet. Sel. Evol. 2017, 49, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, Z.; Ding, R.; Peng, L.; Wu, J.; Ye, Y.; Zhou, S.; Wang, X.; Quan, J.; Zheng, E.; Cai, G.; et al. Genome-wide association analyses identify known and novel loci for teat number in Duroc pigs using single-locus and multi-locus models. BMC Genom. 2020, 21, 344. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, Y.; Zhang, T.; Zhang, L.; Yan, H.; Liu, X.; Wang, L. Genotyping by sequencing reveals a new locus for pig teat number. Anim. Genet. 2017, 48, 470–472. [Google Scholar] [CrossRef]
- Tomiyama, M.; Kanetani, T.; Tatsukawa, Y.; Mori, H.; Oikawa, T. Genetic parameters for preweaning and early growth traits in Berkshire pigs when creep feeding is used. J. Anim. Sci. 2010, 88, 879–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Ding, X.; Tan, Z.; Xing, K.; Yang, T.; Wang, Y.; Sun, D.; Wang, C. Genome-wide association study for reproductive traits in a Large White pig population. Anim. Genet. 2018, 49, 127–131. [Google Scholar] [CrossRef] [Green Version]
- Akanno, E.C.; Chen, L.; Abo-Ismail, M.K.; Crowley, J.J.; Wang, Z.; Li, C.; Basarab, J.A.; MacNeil, M.D.; Plastow, G.S. Genome-wide association scan for heterotic quantitative trait loci in multi-breed and crossbred beef cattle. Genet. Sel. Evol. 2018, 50, 48. [Google Scholar] [CrossRef] [Green Version]
- Georges, M. Mapping, fine mapping, and molecular dissection of quantitative trait Loci in domestic animals. Annu. Rev. Genom. Hum. Genet. 2007, 8, 131–162. [Google Scholar] [CrossRef] [Green Version]
- Andersson, L. Molecular consequences of animal breeding. Curr. Opin. Genet. Dev. 2013, 23, 295–301. [Google Scholar] [CrossRef]
- Hong, Y.F.; Ye, J.; Dong, L.S.; Li, Y.L.; Yan, L.M.; Cai, G.Y.; Liu, D.W.; Tan, C.; Wu, Z.F. Genome-Wide Association Study for Body Length, Body Height, and Total Teat Number in Large White Pigs. Front. Genet. 2021, 12, 650370. [Google Scholar] [CrossRef]
- Mikawa, S.; Sato, S.; Nii, M.; Morozumi, T.; Yoshioka, G.; Imaeda, N.; Yamaguchi, T.; Hayashi, T.; Awata, T. Identification of a second gene associated with variation in vertebral number in domestic pigs. BMC Genet. 2011, 12, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uffelmann, E.; Huang, Q.Q.; Munung, N.S.; de Vries, J.; Okada, Y.; Martin, A.R.; Martin, H.C.; Lappalainen, T.; Posthuma, D. Genome-wide association studies. Nat. Rev. Methods Prim. 2021, 1, 59. [Google Scholar] [CrossRef]
- Duijvesteijn, N.; Veltmaat, J.M.; Knol, E.F.; Harlizius, B. High-resolution association mapping of number of teats in pigs reveals regions controlling vertebral development. BMC Genom. 2014, 15, 542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Son, M.; Lopes, M.S.; Martell, H.J.; Derks, M.; Gangsei, L.E.; Kongsro, J.; Wass, M.N.; Grindflek, E.H.; Harlizius, B. A QTL for Number of Teats Shows Breed Specific Effects on Number of Vertebrae in Pigs: Bridging the Gap Between Molecular and Quantitative Genetics. Front. Genet. 2019, 10, 272. [Google Scholar] [CrossRef] [PubMed]
- Veltmaat, J.M. Prenatal Mammary Gland Development in the Mouse: Research Models and Techniques for Its Study from Past to Present. Methods Mol. Biol. 2017, 1501, 21–76. [Google Scholar] [PubMed]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Stephens, M. Genome-wide efficient mixed-model analysis for association studies. Nat. Genet. 2012, 44, 821–824. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Lee, S.H.; Goddard, M.E.; Visscher, P.M. GCTA: A tool for genome-wide complex trait analysis. Am. J. Hum. Genet. 2011, 88, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.X.; Gao, G.X.; Zhou, Y.; Guo, C.X.; Li, B.; El-Ashram, S.; Li, Z.L. Genome-wide association studies uncover genes associated with litter traits in the pig. Animal 2022, 16, 100672. [Google Scholar] [CrossRef]
- Vasiliou, V.; Vasiliou, K.; Nebert, D.W. Human ATP-binding cassette (ABC) transporter family. Hum. Genom. 2009, 3, 281–290. [Google Scholar] [CrossRef]
- Fettelschoss, V.; Burda, P.; Sagne, C.; Coelho, D.; De Laet, C.; Lutz, S.; Suormala, T.; Fowler, B.; Pietrancosta, N.; Gasnier, B.; et al. Clinical or ATPase domain mutations in ABCD4 disrupt the interaction between the vitamin B (12)-trafficking proteins ABCD4 and LMBD1. J. Biol. Chem. 2017, 292, 11980–11991. [Google Scholar] [CrossRef] [Green Version]
- Ray, J.G.; Blom, H.J. Vitamin B12 insufficiency and the risk of fetal neural tube defects. QJM Int. J. Med. 2003, 96, 289–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groenen, P.M.; van Rooij, I.A.; Peer, P.G.; Gooskens, R.H.; Zielhuis, G.A.; Steegers-Theunissen, R.P. Marginal maternal vitamin B12 status increases the risk of offspring with spina bifida. Am. J. Obstet. Gynecol. 2004, 191, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Zhang, H.; Zhang, Z.; Gao, J.; Yang, J.; Wu, Z.; Fan, Y.; Xing, Y.; Li, L.; Xiao, S.; et al. VRTN is Required for the Development of Thoracic Vertebrae in Mammals. Int. J. Biol. Sci. 2018, 14, 667–681. [Google Scholar] [CrossRef]
- Spina, E.; Cowin, P. Embryonic mammary gland development. Semin. Cell Dev. Biol. 2021, 114, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Robinson, G.W. Cooperation of signalling pathways in embryonic mammary gland development. Nat. Rev. Genet. 2007, 8, 963–972. [Google Scholar] [CrossRef]
- Capuco, A.V.; Ellis, S.E. Comparative aspects of mammary gland development and homeostasis. Annu. Rev. Anim. Biosci. 2013, 1, 179–202. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Huang, L.; Yang, M.; Fan, Y.; Li, L.; Fang, S.; Deng, W.; Cui, L.; Zhang, Z.; Ai, H.; et al. Possible introgression of the VRTN mutation increasing vertebral number, carcass length and teat number from Chinese pigs into European pigs. Sci. Rep. 2016, 6, 19240. [Google Scholar] [CrossRef] [Green Version]
- Schaid, D.J.; Chen, W.; Larson, N.B. From genome-wide associations to candidate causal variants by statistical fine-mapping. Nat. Rev. Genet. 2018, 19, 491–504. [Google Scholar] [CrossRef]
- Hiremath, B.; Subramaniam, N.; Chandrashekhar, N. Giant accessory breast: A rare occurrence reported, with a review of the literature. BMJ Case Rep. 2015, 2015, bcr2015210918. [Google Scholar] [CrossRef] [Green Version]
- Chomwisarutkun, K.; Murani, E.; Brunner, R.; Ponsuksili, S.; Wimmers, K. QTL region-specific microarrays reveal differential expression of positional candidate genes of signaling pathways associated with the liability for the inverted teat defect. Anim. Genet. 2013, 44, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Kessaris, N.; Jamen, F.; Rubin, L.L.; Richardson, W.D. Cooperation between sonic hedgehog and fibroblast growth factor/MAPK signalling pathways in neocortical precursors. Development 2004, 131, 1289–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikitin, S.V.; Kniazev, S.P.; Ermolaev, V.I. Model of genetic control of the number and location of nipples in domestic pig. Russ. J. Genet. 2012, 48, 1128–1140. [Google Scholar] [CrossRef]
- Fernandez, A.; Toro, M.; Rodriguez, C.; Silio, L. Heterosis and epistasis for teat number and fluctuating asymmetry in crosses between Jiaxing and Iberian pigs. Heredity 2004, 93, 222–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Zhao, W.; Fu, Y.; Fang, X.; Ren, S.; Ren, J. Genome-wide detection of genetic loci and candidate genes for teat number and body conformation traits at birth in Chinese Sushan pigs. Anim. Genet. 2019, 50, 753–756. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.S.; Bastiaansen, J.W.; Harlizius, B.; Knol, E.F.; Bovenhuis, H. A genome-wide association study reveals dominance effects on number of teats in pigs. PLoS ONE 2014, 9, e105867. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| chr | SNP ID | Position (bp) | p-Value | Gene | Region |
|---|---|---|---|---|---|
| 9 | ALGA0054033 | 82,853,264 | 3.94 × 10−5 | / | / |
| 7 | Affx-114687136 | 97,568,284 | 6.12 × 10−5 | ABCD4 | 3′ UTR |
| 7 | Affx-115258151 | 97,595,573 | 6.29 × 10−5 | ABCD4 | 5′ UTR |
| 7 | Affx-114892585 | 97,575,068 | 6.32 × 10−5 | ABCD4 | Intron 7-8 |
| 7 | WU_10.2_7_103460706 | 97,617,907 | 7.26 × 10−5 | VRTN | Intron 1-2 |
| 7 | WU_10.2_7_103232787 | 97,584,287 | 8.67 × 10−5 | ABCD4 | Intron 1-2 |
| 8 | ASGA0094767 | 24,709,455 | 1.16 × 10−4 | / | / |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Li, X.; Zhuang, Z.; Zhou, S.; Wu, J.; Xu, C.; Ruan, D.; Qiu, Y.; Zhao, H.; Zheng, E.; et al. Genome-Wide Association Study Identifies the Crucial Candidate Genes for Teat Number in Crossbred Commercial Pigs. Animals 2023, 13, 1880. https://doi.org/10.3390/ani13111880
Yang L, Li X, Zhuang Z, Zhou S, Wu J, Xu C, Ruan D, Qiu Y, Zhao H, Zheng E, et al. Genome-Wide Association Study Identifies the Crucial Candidate Genes for Teat Number in Crossbred Commercial Pigs. Animals. 2023; 13(11):1880. https://doi.org/10.3390/ani13111880
Chicago/Turabian StyleYang, Lijuan, Xuehua Li, Zhanwei Zhuang, Shenping Zhou, Jie Wu, Cineng Xu, Donglin Ruan, Yibin Qiu, Hua Zhao, Enqin Zheng, and et al. 2023. "Genome-Wide Association Study Identifies the Crucial Candidate Genes for Teat Number in Crossbred Commercial Pigs" Animals 13, no. 11: 1880. https://doi.org/10.3390/ani13111880
APA StyleYang, L., Li, X., Zhuang, Z., Zhou, S., Wu, J., Xu, C., Ruan, D., Qiu, Y., Zhao, H., Zheng, E., Cai, G., Wu, Z., & Yang, J. (2023). Genome-Wide Association Study Identifies the Crucial Candidate Genes for Teat Number in Crossbred Commercial Pigs. Animals, 13(11), 1880. https://doi.org/10.3390/ani13111880