


Efficacy of Protein Baits with Fipronil to Control Vespa velutina nigrithorax (Lepeletier, 1836) in Apiaries
 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
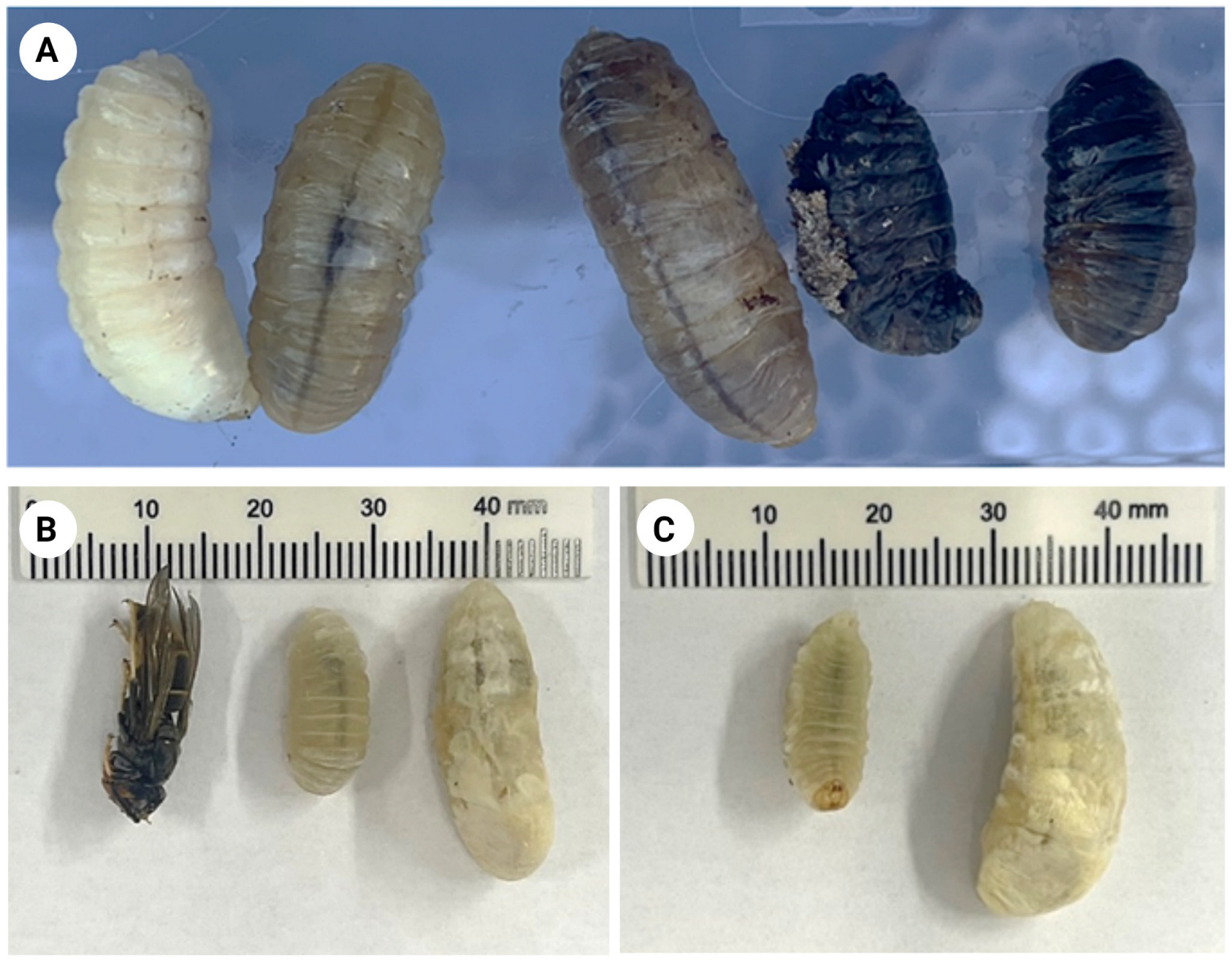
2.1. Larvae Collection
2.2. Larval Inactivation Assays in the Laboratory
2.3. Analysis of Fipronil Content in Dead Larvae
2.4. Study Area and Selection of Beekeepers
2.5. Testing the Efficacy of Baits in Apiaries
2.6. Statistical Analyses
3. Results
3.1. Larval Inactivation Assays in the Laboratory
3.2. Analysis of the Fipronil Content in Dead Larvae
3.3. Testing the Efficacy of Protein Baits with Fipronil in Apiaries
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bertelsmeier, C. Globalization and the anthropogenic spread of invasive social insects. Curr. Opin. Insect Sci. 2021, 46, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Keller, R.P.; Geist, J.; Jeschke, J.M.; Kühn, I. Invasive species in Europe: Ecology, status, and policy. Environ. Sci. Eur. 2011, 23, 23. [Google Scholar] [CrossRef] [Green Version]
- Puebla, H.; Roy, P.K.; Velasco-Perez, A.; Gonzalez-Brambila, M.M. Biological pest control using a model-based robust feedback. IET Syst. Biol. 2018, 12, 233–240. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species. 2020. Available online: https://portals.iucn.org/library/sites/library/files/documents/2000-126-Es.pdf (accessed on 12 June 2023).
- Haxaire, J.; Bouguet, J.; Tamisier, J.P. Vespa velutina Lepeletier, 1836, une redoutable nouveauté pour la faune de France et d’Europe (Hym., Vespidae). Bull. Société Entomol. Fr. 2006, 111, 194. [Google Scholar] [CrossRef]
- Villemant, C.; Haxaire, J.; Streito, J.C. Premier bilan de l’invasion de Vespa velutina Lepeletier en France (Hymenoptera, Vespidae). Bull. Société Entomol. Fr. 2006, 111, 535–538. [Google Scholar] [CrossRef]
- Laurino, D.; Leza, M.; Carisio, L.; Manino, A.; Porporato, M. Vespa velutina: An alien driver of honey bee colony losses. Diversity 2020, 12, 5. [Google Scholar] [CrossRef] [Green Version]
- Dillane, E.; Hayden, R.; O’Hanlon, A.; Butler, F.; Harrison, S. The first recorded occurrence of the Asian hornet (Vespa velutina) in Ireland, genetic evidence for a continued single invasion across Europe. J. Hymenopt. Res. 2022, 93, 131–138. [Google Scholar] [CrossRef]
- Inventaire National du Patrimoine Naturel, 2023. Le frelon asiatique arrive en Suisse. Available online: https://frelonasiatique.mnhn.fr/le-frelon-asiatique-arrive-en-suisse (accessed on 12 June 2023).
- Villemant, C.; Barbet-Massin, M.; Perrard, A.; Muller, F.; Gargominy, O.; Jiguet, F.; Rome, Q. Predicting the invasion risk by the alien bee-hawking yellow-legged hornet Vespa velutina nigrithorax across Europe and other continents with niche models. Biol. Conserv. 2011, 144, 2142–2150. [Google Scholar] [CrossRef]
- Bunker, S. The Asian Hornet Handbook; Psocid Press: UK, 2019; 164p, Available online: https://www.asianhornethandbook.com/ (accessed on 20 June 2022).
- Pérez-de-Heredia, I.; Darrouzet, E.; Goldarazena, A.; Romón, P.; Iturrondobeitia, J.C. Differentiating between gynes and workers in the invasive hornet Vespa velutina (Hymenoptera, Vespidae) in Europe. J. Hymenopt. Res. 2017, 60, 119–133. [Google Scholar] [CrossRef] [Green Version]
- Chauzat, M.P.; Martin, S. A foreigner in France: The Asian hornet. Biologist 2009, 56, 86–91. [Google Scholar]
- Monceau, K.; Bonnard, O.; Thiery, D. Vespa velutina: A new invasive predator of honeybees in Europe. J. Pest Sci. 2014, 87, 1–16. [Google Scholar] [CrossRef]
- Rome, Q.; Muller, F.J.; Touret-Alby, A.; Darrouzet, E.; Perrard, A.; Villemant, C. Caste differentiation and seasonal changes in Vespa velutina (Hym.: Vespidae) colonies in its introduced range. J. Appl. Entomol. 2015, 139, 771–782. [Google Scholar] [CrossRef]
- Leza, M.; Miranda, M.A.; Colomar, V. First detection of Vespa velutina nigrithorax (Hymenoptera: Vespidae) in the Balearic Islands (Western Mediterranean): A challenging study case. Biol. Invasions 2017, 20, 1643–1649. [Google Scholar] [CrossRef]
- Leza, M.; Herrera, C.; Marques, A.; Roca, P.; Sastre-Serra, J.; Pons, D.G. The impact of the invasive species Vespa velutina on honeybees: A new approach based on oxidative stress. Sci. Total Environ. 2019, 689, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Lioy, S.; Bergamino, C.; Porporato, M. The invasive hornet Vespa velutina: Distribution, impacts and management options. CABI Rev. 2022, 1643–1649. [Google Scholar] [CrossRef]
- Rojas-Nossa, S.V.; Calviño-Cancela, M. The invasive hornet Vespa velutina affects pollination of a wild plant through changes in abundance and behaviour of floral visitors. Biol. Invasions 2020, 22, 2609–2618. [Google Scholar] [CrossRef]
- Feas, X. Human fatalities caused by hornet, wasp and bee stings in Spain: Epidemiology at state and sub-state level from 1999 to 2018. Biology 2021, 10, 73. [Google Scholar] [CrossRef]
- Schwartz, C.; Villemant, C.; Rome, Q.; Muller, F. Vespa velutina (frelon asiatique): Un nouvel hyménoptère en France. Rev. Fr. Allergol. 2012, 52, 397–401. [Google Scholar] [CrossRef]
- de Haro, L.; Labadie, M.; Chanseau, P.; Cabot, C.; Blanc-Brisset, I.; Penouil, F. Medical consequences of the Asian black hornet (Vespa velutina) invasion in Southwestern France. Toxicon 2010, 55, 650–652. [Google Scholar] [CrossRef]
- Ruiz-Cristi, I.; Berville, L.; Darrouzet, E. Characterizing thermal tolerance in the invasive yellow-legged hornet (Vespa velutina nigrithorax): The first step toward a green control method. PLoS ONE 2020, 15, e0239742. [Google Scholar] [CrossRef]
- Dalmon, A.; Gayral, P.; Decante, D.; Klopp, C.; Bigot, D.; Thomasson, M.; Herniou, E.A.; Alaux, C.; Le, C.Y. Viruses in the invasive hornet Vespa velutina. Viruses 2019, 11, 1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turchi, L.; Derijard, B. Options for the biological and physical control of Vespa velutina nigrithorax (Hym.: Vespidae) in Europe: A review. J. Appl. Entomol. 2018, 142, 553–562. [Google Scholar] [CrossRef]
- Pazos, T.; Alvarez-Figueiro, P.; Cortes-Vazquez, J.A.; Jacome, M.A.; Servia, M.J. Of fears and budgets: Strategies of control in Vespa velutina invasion and lessons for best management practices. Environ. Manag. 2022, 70, 605–617. [Google Scholar] [CrossRef] [PubMed]
- Leza, M.; Herrera, C.; Pico, G.; Morro, T.; Colomar, V. Six years of controlling the invasive species Vespa velutina in a Mediterranean island: The promising results of an eradication plan. Pest Manag. Sci. 2021, 77, 2375–2384. [Google Scholar] [CrossRef] [PubMed]
- Lioy, S.; Laurino, D.; Capello, M.; Romano, A.; Manino, A.; Porporato, M. Effectiveness and selectiveness of traps and baits for catching the invasive hornet Vespa velutina. Insects 2020, 11, 706. [Google Scholar] [CrossRef]
- Rojas-Nossa, S.V.; Novoa, N.; Serrano, A.; Calviño-Cancela, M. Performance of baited traps used as control tools for the invasive hornet Vespa velutina and their impact on non-target insects. Apidologie 2018, 49, 872–885. [Google Scholar] [CrossRef] [Green Version]
- Requier, F.; Rome, Q.; Villemant, C.; Henry, M. A biodiversity-friendly method to mitigate the invasive Asian hornet’s impact on European honey bees. J. Pest Sci. 2019, 93, 9. [Google Scholar] [CrossRef]
- Demichelis, S.; Manino, A.; Minuto, G.; Mariotti, M.; Porporato, M. Social wasp trapping in north west Italy: Comparison of different bait-traps and first detection of Vespa velutina. Bull. Insectology 2014, 67, 307–317. [Google Scholar]
- Rojas-Nossa, S.V.; Dasilva-Martins, D.; Mato, S.; Bartolomé, C.; Maside, X.; Garrido, J. Effectiveness of electric harps in reducing Vespa velutina predation pressure and consequences for honey bee colony development. Pest Manag. Sci. 2022, 78, 5142–5149. [Google Scholar] [CrossRef]
- Sánchez, O.; Arias, A. All that glitters is not gold: The other insects that fall into the Asian yellow-legged hornet Vespa velutina ‘specific’ traps. Biology 2021, 10, 448. [Google Scholar] [CrossRef]
- Cappa, F.; Cini, A.; Pepiciello, I.; Petrocelli, I.; Inghilesi, A.F.; Anfora, G.; Dani, F.R.; Bortolotti, L.; Wen, P.; Cervo, R. Female volatiles as sex attractants in the invasive population of Vespa velutina nigrithorax. J. Insect. Physiol. 2019, 119, 103952. [Google Scholar] [CrossRef]
- Couto, A.; Monceau, K.; Bonnard, O.; Thiery, D.; Sandoz, J.C. Olfactory attraction of the hornet Vespa velutina to honeybee colony odors and pheromones. PLoS ONE 2014, 9, e115943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, P.; Cheng, Y.N.; Dong, S.H.; Wang, Z.W.; Tan, K.; Nieh, J.C. The sex pheromone of a globally invasive honey bee predator, the Asian eusocial hornet, Vespa velutina. Sci. Rep. 2017, 7, 12956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beggs, J.; Brockerhoff, E.; Corley, J.; Kenis, M.; Masciocchi, M.; Muller, F.; Rome, Q.; Villemant, C. Ecological effects and management of invasive alien Vespidae. Biocontrol 2011, 56, 505–526. [Google Scholar] [CrossRef]
- Hanna, C.; Foote, D.; Kremen, C. Short- and long-term control of Vespula pensylvanica in Hawaii by fipronil baiting. Pest Manag. Sci. 2012, 68, 1026–1033. [Google Scholar] [CrossRef]
- Sackmann, P.; Rabinovich, M.; Corley, J.C. Successful removal of German yellowjackets (Hymenoptera: Vespidae) by toxic baiting. J. Econ. Entomol. 2001, 94, 811–816. [Google Scholar] [CrossRef]
- Spurr, E.B. Protein bait preferences of wasps (Vespula vulgaris and V. germanica) at Mt Thomas, Canterbury, New Zealand. N. Z. J. Zool. 1995, 22, 281–289. [Google Scholar] [CrossRef] [Green Version]
- Edwards, E.; Toft, R.; Joice, N.; Westbrooke, I. The efficacy of Vespex® wasp bait to control Vespula species (Hymenoptera: Vespidae) in New Zealand. Int. J. Pest Manag. 2017, 63, 266–272. [Google Scholar] [CrossRef]
- Edwards, E.D.; Woolly, E.F.; McLellan, R.M.; Keyzers, R.A. Non-detection of honeybee hive contamination following Vespula wasp baiting with protein containing fipronil. PLoS ONE 2018, 13, e0206385. [Google Scholar] [CrossRef] [Green Version]
- Lester, P.J.; Beggs, J.R. Invasion success and management strategies for social Vespula wasps. Annu. Rev. Entomol. 2019, 64, 51–71. [Google Scholar] [CrossRef]
- Poché, D.M.; Franckowiak, G.; Clarke, T.; Tseveenjav, B.; Polyakova, L.; Poche, R.M. Efficacy of a low dose fipronil bait against blacklegged tick (Ixodes scapularis) larvae feeding on white-footed mice (Peromyscus leucopus) under laboratory conditions. Parasit. Vectors 2020, 13, 391. [Google Scholar] [CrossRef]
- Peters, B.C.; Wibowo, D.; Yang, G.Z.; Hui, Y.; Middelberg, A.P.J.; Zhao, C.X. Evaluation of baiting fipronil-loaded silica nanocapsules against termite colonies in fields. Heliyon 2019, 5, e02277. [Google Scholar] [CrossRef] [Green Version]
- Anasac Ambiental, SA. Vespugard 1,5% DP. 2023. Available online: https://www.anasaccontrol.cl/producto/vespugard-15-dp/ (accessed on 1 February 2023).
- Harper, G.; Joice, N.; Kelly, D.; Toft, R.; Clapperton, B. Effective distances of wasp (Vespula vulgaris) poisoning using clustered bait stations in beech forest. N. Z. J. Ecol. 2015, 40, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Castilhos, D.; Dombroski, J.L.D.; Bergamo, G.C.; Gramacho, K.P.; Gonçalves, L.S. Neonicotinoids and fipronil concentrations in honeybees associated with pesticide use in Brazilian agricultural areas. Apidologie 2019, 50, 657–668. [Google Scholar] [CrossRef]
- European Parliament and Council of the European Union. Regulation (EU) No 540/2011 of 25 May 2011 implementing Regulation (EC) No 1107/2009 of the European Parliament and of the Council as regards the list of approved active substances. OJEU 2011, L153, 1–186. [Google Scholar]
- European Commission. Analytical Quality Control and Method Validation Procedures for Pesticide Residues Analysis in Food and Feed SANTE 11312/2021. 2021. Available online: https://www.eurl-pesticides.eu/userfiles/file/EurlALL/SANTE_11312_2021.pdf (accessed on 1 February 2023).
- R Core Team. A Language and Environment for Statistical Computing. 2019. Available online: https://www.r-project.org/ (accessed on 1 November 2022).
- Rome, Q.; Sourdeau, C.; Muller, F.; Villemant, C. Le piégeage du frelon asiatique Vespa velutina. Intérêts et dangers. In Proceedings of the Journée Nationale des GTV, Nantes, France, 15 May 2013; pp. 783–788. [Google Scholar]
- Ulloa, A.; Araya, J.; Curkovic, T. Toxicidad oral de seis insecticidas en larvas de Vespula germanica (F.) en laboratorio. Agric. Técnica 2006, 66, 133–140. [Google Scholar] [CrossRef]
- Curkovic, T.; Santibañez, D.; Araya, J.; Contreras, A. Attraction of Vespula germanica workers to protein baits mixed with insecticideses. Chil. J. Agric. Anim. Sci. 2018, 34, 199–204. [Google Scholar] [CrossRef]
- Durham, E.W.; Siegfried, B.D.; Scharf, M.E. In vivo and in vitro metabolism of fipronil by larvae of the European corn borer Ostrinia nubilalis. Pest Manag. Sci. 2002, 58, 799–804. [Google Scholar] [CrossRef]
- Scharf, M.; Siegfried, B.; Meinke, L.; Chandler, L. Fipronil metabolism, oxidative sulfone formation and toxicity among organophosphate- and carbamate-resistant and susceptible western corn rootworm populations. Pest Manag. Sci. 2000, 56, 757–766. [Google Scholar] [CrossRef]
- Aajoud, A.; Ravanel, P.; Tissut, M. Fipronil metabolism and dissipation in a simplified aquatic ecosystem. J Agric. Food Chem. 2003, 51, 1347–1352. [Google Scholar] [CrossRef]
- Rust, M.K.; Choe, D.H.; Wilson-Rankin, E.; Campbell, K.; Kabashima, J.; Dimson, M. Controlling yellow jackets with fipronil-based protein baits in urban recreational areas. Int. J. Pest Manag. 2017, 63, 234–241. [Google Scholar] [CrossRef] [Green Version]
- Kishi, S.; Goka, K. Review of the invasive yellow-legged hornet, Vespa velutina nigrithorax (Hymenoptera: Vespidae), in Japan and its possible chemical control. Appl. Entomol. Zool. 2017, 52, 361–368. [Google Scholar] [CrossRef]
- European Food Safety Authority. Conclusion on the peer review of the pesticide risk assessment for bees for the active substance fipronil. EFSA J. 2013, 11, 3158. [Google Scholar] [CrossRef]
- Aliouane, Y.; El Hassani, A.K.; Gary, V.; Armengaud, C.; Lambin, M.; Gauthier, M. Subchronic exposure of honeybees to sublethal doses of pesticides: Effects on behavior. Environ. Toxicol. Chem. 2009, 28, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Villemant, C.; Muller, F.; Rome, Q.; Perrard, A.; Barbet-Massin, M.; Jiguet, F. Estimating the potential range expansion and environmental impact of the invasive bee-hawking hornet, Vespa velutina nigrithorax. In Silico Bees, 1st ed.; Devillers, J., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 269–287. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fipronil % (w/w) | Fed Larvae | Inactive Larvae (%) at 24 h |
|---|---|---|
| 0.003 | 26 | 42.3 |
| 0.004 | 33 | 63.6 |
| 0.007 | 48 | 68.8 |
| 0.01 | 44 | 84.1 |
| Control | 46 | 0 |
| Year | No. Beekeepers | No. Apiaries | No. Hives | No. Baitings | Bait (kg) | Questionaries Fulfilled |
|---|---|---|---|---|---|---|
| 2019 | 96 | 153 | 1562 | 169 | 23.4 | 34 |
| 2020 | 97 | 117 | 1004 | 123 | 14.2 | 37 |
| 2021 | 29 | 31 | 290 | 36 | 7.2 | 19 |
| Group of Apiaries | Day 0 | Day +2 | Day +7 | Day +14 | |
|---|---|---|---|---|---|
| All apiaries | Ⴟ (SD) | 22.6 (24.5) | 10.2 (9.2) | 11.1 (13.1) | 10.7 (9.7) |
| RE% | 41.8 | 43.4 | 38.8 | ||
| Apiaries <10 hornets | Ⴟ (SD) | 6.5 (2.7) | 5.6 (3.9) | 5.2 (3.1) | 7.1 (6.5) |
| RE% | 26.4 | 32.4 | 25.1 | ||
| Apiaries 10–30 hornets | Ⴟ (SD) | 20.9 (6.4) | 13.1 (10.4) | 14.1 (13.0) | 13.7 (10.3) |
| RE% | 45.4 | 43.8 | 41.7 | ||
| Apiaries >30 hornets | Ⴟ (SD) | 67.1 (28.9) | 15.6 (10.7) | 19.2 (21.2) | 14.9 (13.7) |
| RE% | 74.0 | 70.1 | 77.4 |
| Group of Apiaries | Estimate | Std. Error | z-Value | p-Value | |
|---|---|---|---|---|---|
| Apiaries <10 hornets | Intercept | 1.55763 | 0.13537 | 11.506 | <0.001 *** |
| Day +2 | −0.06645 | 0.12064 | −0.551 | 0.582 NS | |
| Day +7 | −0.03637 | 0.11971 | −0.304 | 0.761 NS | |
| Day +14 | 0.26343 | 0.11328 | 2.325 | 0.020 * | |
| Apiaries 10–30 hornets | Intercept | 2.86838 | 0.09516 | 30.144 | <0.001 *** |
| Day +2 | −0.56898 | 0.05706 | −9.971 | <0.001 *** | |
| Day +7 | −0.50479 | 0.05626 | −8.972 | <0.001 *** | |
| Day +14 | −0.49360 | 0.05901 | −8.365 | <0.001 *** | |
| Apiaries >30 hornets | Intercept | 3.99687 | 0.10193 | 39.21 | <0.001 *** |
| Day +2 | −1.27011 | 0.06120 | −20.75 | <0.001 *** | |
| Day +7 | −1.16684 | 0.05938 | −19.65 | <0.001 *** | |
| Day +14 | −0.86979 | 0.06366 | −13.66 | <0.001 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barandika, J.F.; de la Hera, O.; Fañanás, R.; Rivas, A.; Arroyo, E.; Alonso, R.M.; Alonso, M.L.; Galartza, E.; Cevidanes, A.; García-Pérez, A.L. Efficacy of Protein Baits with Fipronil to Control Vespa velutina nigrithorax (Lepeletier, 1836) in Apiaries. Animals 2023, 13, 2075. https://doi.org/10.3390/ani13132075
Barandika JF, de la Hera O, Fañanás R, Rivas A, Arroyo E, Alonso RM, Alonso ML, Galartza E, Cevidanes A, García-Pérez AL. Efficacy of Protein Baits with Fipronil to Control Vespa velutina nigrithorax (Lepeletier, 1836) in Apiaries. Animals. 2023; 13(13):2075. https://doi.org/10.3390/ani13132075
Chicago/Turabian StyleBarandika, Jesús F., Omaira de la Hera, Roberto Fañanás, Arrate Rivas, Eugenia Arroyo, Rosa M. Alonso, M. Luz Alonso, Egoitz Galartza, Aitor Cevidanes, and Ana L. García-Pérez. 2023. "Efficacy of Protein Baits with Fipronil to Control Vespa velutina nigrithorax (Lepeletier, 1836) in Apiaries" Animals 13, no. 13: 2075. https://doi.org/10.3390/ani13132075
APA StyleBarandika, J. F., de la Hera, O., Fañanás, R., Rivas, A., Arroyo, E., Alonso, R. M., Alonso, M. L., Galartza, E., Cevidanes, A., & García-Pérez, A. L. (2023). Efficacy of Protein Baits with Fipronil to Control Vespa velutina nigrithorax (Lepeletier, 1836) in Apiaries. Animals, 13(13), 2075. https://doi.org/10.3390/ani13132075