
Role of Macrophages in the Pathogenesis of Genotype VII Newcastle Disease Virus in Chickens
by

Jie Ni
1,2,3,
Jing Deng
1,2,3,4,
Qing Chen
1,2,3,
Tianxing Liao
1,2,3,
Jiao Hu
1,2,3,
Yu Chen
1,2,3,
Shunlin Hu
1,2,3,
Zenglei Hu
1,2,3,4,* and
Xiufan Liu
1,2,3,4,* 1
Key Laboratory of Animal Infectious Diseases, School of Veterinary Medicine, Yangzhou University, Yangzhou 225009, China
2
Jiangsu Co-Innovation Center for Prevention and Control of Important Animal Infectious Diseases and Zoonosis, Yangzhou University, Yangzhou 225009, China
3
Jiangsu Key Laboratory of Zoonosis, Yangzhou University, Yangzhou 225009, China
4
Joint International Research Laboratory of Agriculture and Agri-Product Safety, The Ministry of Education of China, Yangzhou University, Yangzhou 225009, China
*
Authors to whom correspondence should be addressed.
Animals 2023, 13(13), 2239; https://doi.org/10.3390/ani13132239
Submission received: 2 June 2023
/
Revised: 3 July 2023
/
Accepted: 4 July 2023
/
Published: 7 July 2023
(This article belongs to the Special Issue Viral Infections in Poultry)
Abstract
:Simple Summary
Genotype VII Newcastle disease virus (NDV) is renowned for its high tropism to the immune organs of chickens, and macrophages are a critical component in the antiviral innate immune system. The objective of this study is to investigate the role of macrophages in the pathogenesis of genotype VII NDV in chickens. Genotype VII NDV has significantly higher infectivity in macrophages compared to genotype IV NDV. Macrophage depletion resulted in no changes in virulence and virus replication but a significant deterioration of tissue damage in the spleen. Therefore, macrophages play an important role in alleviating tissue lesions caused by genotype VII NDV infection.
Abstract
Long-term evolution of Newcastle disease virus (NDV) results in substantial alteration in viral pathogenesis. NDVs of genotype VII, a late genotype, show marked tropism to lymphoid tissues, especially to macrophages in chickens. However, the role of macrophages in the pathogenesis of genotype VII NDV is still unclear. Herein, NDV infectivity in macrophages and the role of macrophages in the pathogenesis of genotype VII NDV in chickens were investigated. We reported that NDV strains of genotype VII (JS5/05) and IV (Herts/33) can replicate in the adherent (predominantly macrophages) and non-adherent cells (predominantly lymphocytes) derived from chicken peripheral blood mononuclear cells (PBMCs), and significantly higher virus gene copy was detected in the adherent cells. In addition, JS5/05 had significantly higher infectivity in PBMC-derived adherent cells than Herts/33, correlating with its enhanced tropism to macrophages in the spleen of chickens. Interestingly, the depletion of 68% of macrophages exerted no significant impact on clinical signs, mortality and the systematic replication of JS5/05 in chickens, which may be associated with the contribution of non-depleted macrophages and other virus-supportive cells to virus replication. Macrophage depletion resulted in a marked exacerbation of tissue damage and apoptosis in the spleen caused by JS5/05. These findings indicated that macrophages play a critical role in alleviating tissue damage caused by genotype VII NDV in chickens. Our results unveiled new roles of macrophages in NDV pathogenesis in chickens.
1. Introduction
Newcastle disease (ND) is an acute and highly contagious infectious disease caused by velogenic Newcastle disease virus (NDV). ND causes high mortality in a variety of poultry species and is a serious threat to the development of the poultry industry. NDV is a member of the genus Orthoavulavirus in the family Paramyxoviridae [1]. Velogenic NDV strains induce systematic infection and severe tissue lesions in the respiratory, gastrointestinal, neurological, reproductive and immune systems of poultry [2,3], suggesting viral tropism to a wide range of cell types.
NDV can replicate in various cell types such as epithelial cells, macrophages, dendritic cells and glial cells (neurons, astrocytes, oligodendrocytes and microglia) [4,5,6,7,8]. Specifically, in NDV-infected chickens, extensive distribution of the viral antigens can be detected in macrophage-like cells in lymphoid tissues, indicating the tropism of NDV to macrophages [9,10]. In addition, independent in vitro studies explored the mechanism of NDV infectivity in macrophages from various perspectives. A recent study showed that NDV entry into chicken macrophages is associated with a pH-dependent, dynamin- and caveola-mediated endocytosis pathway, and Rab5 is required for this process [8]. Velogenic NDV induces an M1-/M2-like mixed polarized activation of chicken macrophages through the inhibition of toll-like receptor 7, resulting in enhanced replication compared to lentogenic viruses [11]. Additionally, the fusion and hemagglutinin-neuraminidase proteins are determinants for the macrophage host range of NDV [12]. These studies suggested that macrophages, the main target cells for NDV infection, may play an important role in viral pathogenesis.
NDV has undergone substantial evolution since its emergence in 1926, resulting in prominent alternation of the pathogenesis. Genotype VII NDV, a late genotype emerging in the 1990s [13,14,15], shows high tropism to lymphoid tissues of chickens compared to strains of early genotypes, such as genotype IV. The induction of an aberrant innate immune response contributes to the pathology of the genotype VII virus [16,17,18]. Macrophages, as a key component of the innate immune system, play a crucial antiviral role through clearing apoptotic cells or virus-infected cells and producing cytokines. Additionally, extensive distribution of the viral antigen in macrophage-like cells and remarkable necrosis of these cells were detected in the spleen of chickens infected with genotype VII NDV [9]. Therefore, the critical role of macrophages in the innate immune response and high tropism of genotype VII NDV to this cell type prompted us to hypothesize that macrophages may be engaged in determining the pathogenesis of genotype VII NDV in chickens.
In this study, we reported that genotype VII NDV had higher infectivity in macrophages in vitro and in vivo compared to the genotype IV virus. Macrophage depletion had no influence on the disease outcomes or replication of genotype VII NDV in chickens but exacerbated virus-induced tissue damage and apoptosis in the spleen. This indicates a critical role of macrophages in protection against pathology caused by NDV in chickens. Our results identified new roles of macrophages in the pathogenesis of genotype VII NDV in chickens.
2. Materials and Methods
2.1. Cells, Viruses and Plasmid
Chicken peripheral blood mononuclear cells (PBMCs) were isolated from 2-month-old specific pathogen-free (SPF) white leghorn chickens through density gradient centrifugation using a chicken PBMC isolation kit (TBD Science, Tianjin, China). PBMCs were grown in RPMI 1640 medium supplemented with 10% fetal bovine serum (FBS) (ThermoFisher Scientific, Waltham, MA, USA) at 37 °C, 5% CO2. Chicken embryo fibroblasts (CEFs) were prepared using 9-day-old SPF chicken embryos and cultured in M199 medium supplemented with 4% FBS. Genotype VII NDV JS5/05 and genotype IV NDV Herts/33 were reported previously [16] and were propagated in 9-day-old SPF embryonated chicken eggs (ECEs). Virus titers were measured as 50% tissue culture infectious dose (TCID50) in CEFs and as 50% embryo infectious dose (EID50) in ECEs. The plasmid containing the full-length genome of JS5/05 was generated previously [19], and was used as a standard plasmid in absolute quantitative real-time PCR (qRT-PCR) for virus titration.
2.2. Isolation and Identification of Macrophages from PBMCs
Blood was collected from three 2-month-old SPF white leghorn chickens, pooled and used for PBMC isolation. Macrophages were isolated from PBMCs as previously reported [20]. PBMCs were seeded in a 6-well plate, and the non-adherent cells were removed by washing with PBS at 2 and 24 h after seeding. To determine the ratio of macrophages, the adherent cells were trypsinized to prepare single cells, and 1 × 107 single cells were labeled with FITC-conjugated mouse anti-chicken KUL01 antibody (1:100) (SouthernBiotech, Birmingham, AL, USA) for 45 min at 4 °C in FACS buffer (PBS containing 2% FBS). Then, the cells were washed and resuspended in PBS, and 1 × 104 cells were used for flow cytometry analysis.
2.3. In Vitro Infectivity of NDV in the Adherent and Non-Adherent Cells Derived from PBMC
To assess NDV infectivity in different cell fractions derived from PBMCs, 1 × 107 PBMCs were seeded in 6-well culture plates and then inoculated with JS5/05 or Herts/33 at a multiplicity of infection (MOI) of 0.01. At 8, 16 and 24 h post inoculation (p.i.), the adherent (predominantly macrophages) and non-adherent cells (predominantly lymphocytes) were separated, and total RNA was extracted from the cells and transcribed into cDNA using a PrimeScript RT Reagent Kit with gDNA Eraser (Takara, Shiga, Japan). Virus replication/transcription in the cells was determined by measuring the copy of the fusion (F) gene using qRT-PCR. The F gene was amplified using a pair of primers (sense: 5′-GGTCAATCATAGTCAAGTTGCTCC-3′; anti-sense: 5′-AACCCCAAGAGCTACACTGCC-3′) and a TaqMan probe (5′-FAM-AAGCGTYTYTGTCTCCTTCCTCC-BHQ-3′). The primers and the probe were designed according to the sequences of JS5/05 (accession no. JN631747.1) and Herts/33 (accession no. AY741404.1). In brief, the standard plasmid was 10-fold serially diluted, and the samples containing 102 to 109 copies were used to plot the standard curve. The qRT-PCR reaction system was composed of 10 μL of 2 × AceQ Universal U + Probe Master Mix (Vazyme, Nanjing, China), 0.7 μL of the sense and anti-sense primers (final concentration 0.35 μM), 0.7 μL of the probe, 2 μL of the cDNA and 5.8 μL of deionized water. PCR reactions were performed in triplicate using a LightCycler® 480 (Roche, Basel, Switzerland) with the following cycle profile: 1 cycle at 95 °C for 10 min followed by 40 cycles at 95 °C for 15 s and 60 °C for 35 s. The F gene copy in the NDV-inoculated cells was calculated based on the standard curve. The experiments were performed in triplicate.
2.4. In Vivo Infectivity of NDV in Macrophages
To determine NDV infectivity in macrophages in vivo, chickens were infected with NDV, and virus replication in macrophages in the spleen was measured using immunofluorescence assays (IFA). Briefly, nine 5-week-old SPF chickens were randomly divided into three groups, with three chickens per group. Three SPF chickens were inoculated with 105 EID50 of JS5/05 or Herts/33 via the intranasal and intraocular routes. Another three chickens were inoculated with PBS as the mock control. On day 3 p.i., the chickens were euthanized, and the spleens were collected for the preparation of formalin-fixed paraffin-embedded sections. The nucleoprotein (NP) protein of NDV was detected using mouse anti-NP monoclonal antibody (a gift from Dr. Chan Ding, Shanghai Veterinary Research Institute, China) and the macrophage marker KUL01 was probed using mouse anti-chicken KUL01 antibody (SouthernBiotech). The cell nucleus was stained with 2-(4-Amidinophenyl)-6-indolecarbamidine dihydrochloride (DAPI) (Beyotime, Nantong, China). The entire spleen section was scanned and the images were processed and analyzed using the CaseViewer software (version 2.3).
2.5. Macrophage Depletion in Chickens
Clodronate liposomes (CL lipo) (Yeasen, Shanghai, China) were used for macrophage depletion in chickens as reported previously [21]. Six 5-week-old SPF chickens were randomly divided into two groups, with three chickens in each group. Three 5-week-old SPF chickens were intravenously (i.v.) treated with 250 μL of CL lipo or the control reagent, PBS liposomes (PBS lipo), respectively. The efficiency of macrophage depletion was verified using IFA in the spleen on day 5 post-treatment (pt). The spleens were fixed with 10% buffered formalin and prepared into tissue sections. A mouse anti-chicken KUL01 monoclonal antibody (1:50) was added to the sections as the primary antibody and incubated at 4 °C overnight. After washing with PBS 3 times, the sections were then incubated with Alexa Fluor 647-conjugated goat anti-mouse antibody as the secondary antibody (1:200) (ThermoFisher Scientific) at 37 °C for 1 h. After washing with PBS, the cell nucleus was stained with DAPI. The images were captured using a fluorescence microscope (Leica).
2.6. Chicken Infection Study
Twenty-seven 5-week-old SPF chickens were randomly divided into three groups, nine chickens per group. Two groups of chickens were i.v. treated with 250 μL of CL lipo or PBS lipo, and chickens in the third group were i.v. inoculated with 250 μL of PBS as the mock control. Two days later, the CL lipo- and PBS lipo-treated chickens were inoculated with 105 EID50 of JS5/05 via the intranasal and intraocular routes. The mock chickens were inoculated with PBS via the same route. On days 1 and 3 p.i., three chickens per group were euthanized, and the liver, spleen, lung, thymus and intestine were collected. The remaining three chickens of each group were observed daily for clinical signs and mortality. All the tissues were used for virus load measurement. The tissue samples were homogenized in PBS supplemented with antibiotics (1 mg tissue in 0.3 mL PBS), and the homogenates were centrifugated to collect the supernatants. The supernatants were 10-fold serially diluted using M199 medium supplemented with 1% FBS and were inoculated into CEFs. At 72 h post inoculation, the culture supernatants of the cells were harvested and subjected to hemagglutination assays to determine virus infection. Virus titers were calculated as TCID50 per gram using the Reed and Muench method [22].
2.7. Histopathological Analysis
The spleens were selected for histopathological assessment because JS5/05 causes remarkable lesions in this tissue. The spleens collected on days 1 and 3 p.i. were fixed with 10% buffered formalin, prepared into tissue sections and stained with hematoxylin–eosin (HE). The histopathological changes were scored as reported previously [16]: −, normal; +, mild to moderate lymphocyte proliferation; ++, moderate to marked lymphocyte depletion and necrosis; +++, marked lymphocyte depletion, necrosis and tissue hyperplasia; ++++, extensive and severe lymphocyte depletion and necrosis.
2.8. TUNEL Staining
To evaluate whether macrophage depletion impacts apoptosis induced by NDV, the spleen tissues were examined using a terminal deoxnuceotidyl transferase-mediated dUTP biotin nick end labeling (TUNEL) kit according to the manufacturer’s instructions. Briefly, 50 μL of equilibration buffer was added to the tissue section and incubated at room temperature for 10 min. After removing the equilibration buffer, 56 μL of labeling solution containing recombinant TdT enzyme, FITC-12-dUTP labeling mix and equilibration buffer was added to the tissues and incubated at 37 °C for 1 h. After washing with PBS, the tissue sections were incubated with DAPI for nucleus staining. The images were captured using a fluorescence microscope (Leica).
3. Data analysis
The pathological and immunofluorescence images were processed and analyzed using CaseViewer and ImageJ software (1.52V), and the flow cytometry data were analyzed using FlowJo software (V10). Statistical analysis of the data was performed using GraphPad Prism. The data are presented as the mean values ± standard deviation (SD) of three independent experiments. Statistical differences were analyzed using the t-test or one-way ANOVA. p < 0.05 was considered a significant difference.
4. Results
4.1. NDV Has Higher Infectivity in Macrophages than Lymphocytes
The infectivity of the genotype VII and IV NDV strains in macrophages was assessed in vitro. As previously reported, the majority of the adherent and non-adherent cells derived from PBMCs or splenocytes are macrophages and lymphocytes, respectively [12,20,21]. Herein, macrophages were isolated from chicken PBMCs, and NDV replication/transcription in these cells was determined using qRT-PCR. A high percentage of the adherent cells (72.9%) derived from PBMCs were KUL01-positive (Figure 1A), indicating that the adherent cells were predominantly macrophages. Viral gene copies of JS5/05 and Herts/33 in the adherent and non-adherent cells steadily increased from 8 to 24 h p.i., suggesting that these two cell fractions were supportive for NDV replication/transcription (Figure 1B,C). Overall, JS5/05 had significantly higher virus gene copy in the adherent and non-adherent cells compared to Herts/33 (Figure 1B,C). In addition, at 16 and 24 h p.i., significantly higher viral copy of both NDV strains were detected in the adherent cells compared to those in the non-adherent cells (Figure 1B,C). These data demonstrated that the virus replication/transcription of the JS5/05 and Herts/33 strains was significantly higher in adherent cells compared to non-adherent cells, and JS5/05 had higher infectivity in both cell fractions compared to Herts/33.
4.2. Genotype VII NDV Presents an Enhanced Tropism to Macrophages in Chickens
To further verify NDV infectivity in macrophages in vivo, chickens were infected with NDV, and viral tropism to macrophages in the spleen was determined using IFA. No NP antigen was detected in the spleen of the mock-infected chickens (Figure 2A). An extensive distribution of the NP antigen in the spleen was observed in the JS5/05-infected chickens (Figure 2B), while the NP-positive area was markedly limited in the Herts/33-infected spleens (Figure 2C). In addition, in the virus-infected chicken spleens, the majority of the NP-positive cells were positive for the macrophage marker KUL01 (Figure 2D). The co-localization of the NP antigen and KUL01 marker verified virus tropism to macrophages in vivo (Figure 2E,F). These results showed that NDV exhibited tropism to macrophages in the spleen, and the genotype VII NDV strain JS5/05 presented enhanced infectivity in macrophages compared to the genotype IV NDV strain Herts/33.
4.3. Clodronate Liposome Treatment Causes Efficient Macrophage Depletion
To investigate the contribution of macrophages to NDV pathogenicity, macrophages were depleted in chickens with treatment with CL lipo, a widely-used reagent for in vivo macrophage clearance. Chickens were i.v. injected with 250 μL of CL lipo, and on day 3 p.t., the number of KUL01-positive cells in the spleen of chickens treated with CL lipo decreased significantly compared to the PBS lipo-treated chickens (Figure 3A), resulting in a depletion efficiency of 68%. These data showed that clodronate liposome treatment resulted in efficient macrophage clearance in chickens.
4.4. Macrophage Depletion Does Not Alter Disease Outcomes and NDV Replication in Chickens
Subsequently, chickens were treated with CL lipo, and inoculated with JS5/05 two days later. The CL lipo- and PBS lipo-treated chickens displayed similar clinical signs after NDV infection such as severe depression, apparent respiratory signs and diarrhea, and rapidly died from virus infection within 4 days p.i. (Figure 3B). In addition, no infectious virus was recovered from the spleen, liver, lung, thymus or intestine of the CL lipo- and PBS lipo-treated chickens on day 1 p.i. (Figure 3C), and on day 3 p.i., virus replication was detected in all the tested tissues of the treated chickens (Figure 3D). However, there were no significant differences in virus load in the tested tissues between the chickens treated with CL lipo and PBS lipo (Figure 3D). Together, these results suggested that macrophage depletion did not affect the disease outcomes or systematic replication of NDV in chickens.
4.5. Macrophage Depletion Exacerbates Tissue Damage Caused by NDV
As a key antiviral component, macrophages may be involved in the induction of the innate immune response and pathological lesions caused by NDV. To assess the role of macrophages in NDV pathogenicity, under the condition of macrophage depletion, tissue damage in the spleen caused by JS5/05 was assessed because this strain induces remarkable lesions in this organ. On day 1 p.i., no obvious pathological changes were seen in the non-infected chickens, and JS5/05 induced mild hemorrhage and lymphocyte necrosis in the chickens treated with CL lipo and PBS lipo (Figure 4A). On day 3 p.i., no tissue lesions were detected in the spleen of the non-infected chickens, whereas marked lymphocyte depletion and severe necrosis were observed in the JS5/05-infected chickens after PBS lipo treatment (Figure 4A). Of note, upon CL lipo treatment, JS5/05 infection caused more remarkable pathological changes in the spleen, characterized by enhanced lymphocyte depletion and large and extensive necrotic foci and hemorrhage, compared to that of the PBS lipo-treated chickens (Figure 4A,B) (Table 1). These findings showed that macrophage depletion deteriorated tissue lesion in the spleen caused by JS5/05, indicating a role of macrophages in alleviating tissue damage.
4.6. Macrophage Depletion Enhances Apoptosis Caused by NDV
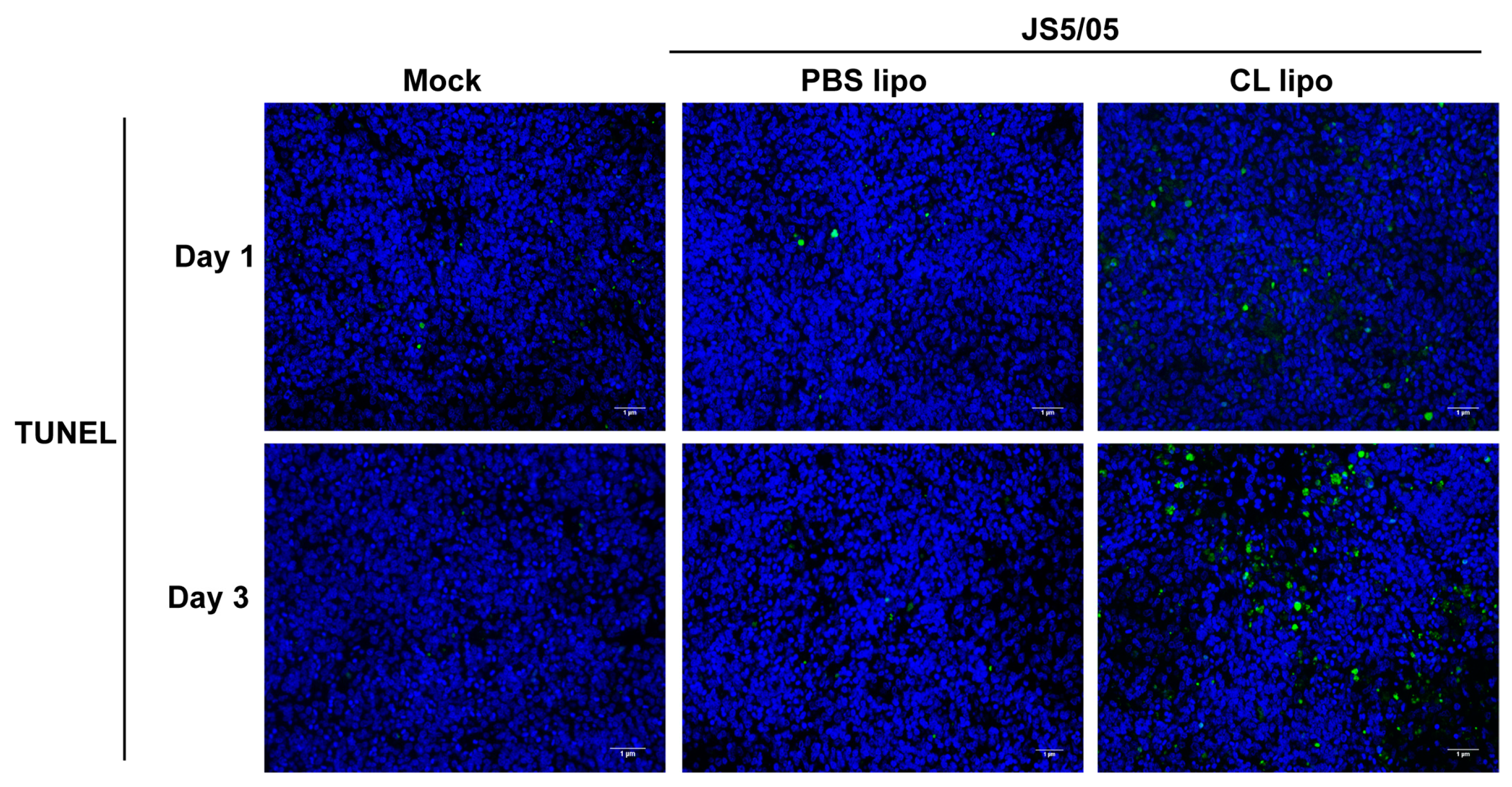
NDV infection induces apoptosis, and macrophages engulf apoptotic cells. To evaluate the impact of macrophage depletion on virus-induced apoptosis, apoptotic cells were detected using TUNEL staining in the spleen. On day 1 p.i., only a few apoptotic cells were detected in the spleen of the non-infected chickens and virus-infected chickens after PBS lipo treatment, and JS5/05 infection resulted in more apoptotic cells in the spleen of the CL lipo-treated chickens (Figure 5). On day 3 p.i., the number of apoptotic cells in the spleen of the chickens treated with CL lipo after JS5/05 infection was significantly increased compared with the PBS lipo-treated and non-infected chickens (Figure 5). Therefore, NDV-induced apoptosis in the spleen was exacerbated after macrophage depletion.
5. Discussion
Genotype VII NDV is characterized by its marked tropism to chicken lymphoid tissues, especially to macrophages. It seems to be plausible that macrophages, an important immune cell type, may play a critical role in regulating virus pathogenesis. Herein, we reported that NDV had higher tropism to the PBMC-derived adherent cells compared to non-adherent cells, and in particular, the genotype VII virus showed enhanced infectivity compared to the genotype IV virus in these cells. When macrophages were depleted by CL lipo treatment, no significant changes in the disease manifestation and virus replication of genotype VII NDV were detected, while virus-induced tissue damage was exacerbated. These results indicated that macrophages protect against pathology caused by NDV infection in chickens.
Macrophages, as an essential component in the innate immune system, are major target cells for NDV infection. Some studies in chickens showed that the viral antigen or nucleic acid of NDV was detected in multiple tissues, but most predominantly in macrophages associated with lymphoid tissues [9,10]. However, in these studies, the virus-infected cell type was assessed mainly based on histological observation. In the present study, a specific marker for chicken macrophages, KUL01, was used to identify the cell type of NDV infection. The genotype VII and IV strains could replicate in KUL01-positive cells in the spleen, confirming the tropism of NDV to macrophages. Of note, the NP antigen was predominantly found in macrophages in the ellipsoids in the white pulp but not in macrophages in the red pulp. This finding correlates with the previous results that NDV mainly replicates in the white pulp in the spleen [9,23,24]. The mechanism for the differential tropism of NDV to macrophages in the white and red pulps is still unknown. Moreover, several in vitro infection studies revealed the infectivity of NDV in primary macrophages or the macrophage cell line [11,12]. In these studies, velogenic strains were compared to lentogenic or mesogenic strains, suggesting that NDV tropism to macrophages correlates with the virulence. Here, we showed that two velogenic strains belonging to different genotypes had distinct infectivity to macrophages, with an enhanced tropism observed for the genotype VII virus. These results indicated that viral tropism to macrophages may change as a result of NDV evolution.
Based on the high tropism of genotype VII NDV to macrophages, the contribution of macrophages to virus replication in chickens was examined through cell depletion. CL lipo was used herein to deplete macrophages in chickens because this reagent is efficient for depleting macrophages in animals. The results showed that 68% of the KUL01-positive cells in the spleen were depleted, and this efficiency for macrophage clearance was comparable to those in previous reports [21,25,26]. Efficient depletion of macrophages is fundamental to examine their roles in NDV pathogenicity. Macrophage depletion resulted in no significant differences in virus load in chicken tissues, indicating that macrophage depletion did not affect the systematic infection of NDV. Similarly, the depletion of macrophages from the lungs via intranasal inoculation of CL lipo did not affect the replication of Middle East respiratory syndrome coronavirus (MERS-CoV) in mice [27]. In contrast, the replication of Marek’s disease virus (MDV) and H3N2 influenza virus in the host was significantly increased after CL lipo treatment [28,29]. The findings obtained in our study seem to be conflicting with the high infectivity of NDV in macrophages. There are three possible explanations for these results. First, after CL lipo treatment, there are still 32% of macrophages remaining in the spleen. This fraction of macrophages is still supportive for NDV infection and may contribute to virus replication in chickens. This possibility may at least partially explain why CL lipo treatment caused no significant decrease in virus load in the tissues. Second, NDV has tropism to various cell types, and macrophage reduction may lead to compensatory increases in virus replication in other cells, and thus the overall virus replication was not affected. Similarly, macrophage depletion resulted in more efficient infection of dendritic cells with measles virus, which increased virus load in the spleen in mice [30]. Third, macrophage depletion may impair the clearance of the virus-infected cells, promoting virus dissemination in chickens.
On the other hand, macrophage depletion markedly deteriorated tissue damage in the spleen caused by NDV. A genotype VII strain was used in this study, and it produces severe necrosis in the lymphoid tissues in chickens [16,31]. Many previous studies revealed that intense cytokine response contributes to pathology induced by genotype VII NDV infection [16,17,18]. Macrophages produce cytokines after virus infection. Thus, it seems to be plausible that macrophage depletion may lead to decreased production of cytokines and, thus, impairment of tissue injury. However, our findings showed that macrophage depletion resulted in marked aggravation of tissue damage caused by JS5/05. We speculated that virus infection and spreading in the spleen may accelerate because of the reduced amount of macrophages. In addition, NDV infection induces apoptosis, which is associated with tissue lesions. Macrophages function to engulf and clear virus-induced apoptotic cells. NDV infection after macrophage depletion enhanced splenic pathology, which may be also related to the impaired capability to clear apoptotic and necrotic cells induced by virus infection due to the decreased amount of macrophages. These findings were consistent with the results for many other pathogens, including measles virus, H3N2 influenza virus, MERS-CoV and MDV [27,28,29,30]. Therefore, macrophages function to protect against NDV-mediated pathology. CL lipo treatment causes macrophage depletion through inducing cell apoptosis, and previous studies demonstrated that NDV infection can also trigger apoptosis of macrophages [5,23]. Thus, it is reasonable to speculate that genotype VII NDV may induce apoptosis of macrophages, which undermines the host antiviral response and promotes tissue damage caused by the virus. Further studies are required to investigate the effect of NDV-induced cell death of macrophages on viral pathogenicity.
6. Conclusions
Our study demonstrated that genotype VII NDV had high tropism to macrophages, whereas the contribution of these cells to virus replication could be compensated for by other cells when they were depleted. Alternatively, macrophages played an important role in alleviating NDV-induced tissue damage. Macrophage depletion exacerbated virus-induced pathology without impairing virus replication. Our findings identified new roles of macrophages in NDV replication and pathogenicity in chickens.
Author Contributions
Conceptualization: J.N., Z.H. and X.L.; methodology: J.N. and Z.H.; validation: J.D. and Q.C.; investigation: J.N., J.D. and T.L.; writing—original draft preparation: J.N. and Z.H.; writing—review and editing: J.H., Y.C. and S.H. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Jiangsu Provincial Natural Science Fund (BK20201433), the National Natural Science Foundation of China (31702243), the Earmarked Fund for China Agriculture Research System (No. CARS-40), Postgraduate Research & Practice Innovation Program of Jiangsu Province (KYCX22_3554), the “High-end Talent Support Program” of Yangzhou University and by a project funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Institutional Review Board Statement
All animal experiments were approved by the Jiangsu Administrative Committee for Laboratory Animals (Permission number: SYXK-SU-2007-0005) and complied with the guidelines of Jiangsu laboratory animal welfare and ethics of Jiangsu Administrative Committee of Laboratory Animals. Experiments involving virulent NDVs were performed in animal biosafety level 3 facilities.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to acknowledge Ya Huang, Yanyan Zhang, Xiaozheng He and Huafen Zheng for the help with the chicken infection studies.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Amarasinghe, G.K.; Ayllón, M.A.; Bào, Y.; Basler, C.F.; Bavari, S.; Blasdell, K.R.; Briese, T.; Brown, P.A.; Bukreyev, A.; Balkema-Buschmann, A.; et al. Taxonomy of the order Mononegavirales: Update 2019. Arch. Virol. 2019, 164, 1967–1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, P.J.; Koch, G. Newcastle Disease. In Diseases of Poultry, 14th ed.; Swayne, D.E., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; pp. 112–134. [Google Scholar]
- Zhang, D.; Ding, Z.; Xu, X. Pathologic mechanisms of the Newcastle disease virus. Viruses 2023, 15, 864. [Google Scholar] [CrossRef]
- Kaiser, A.; Willer, T.; Sid, H.; Petersen, H.; Baumgartner, W.; Steinberg, P.; Rautenschlein, S. Susceptibility of primary chicken intestinal epithelial cells for low pathogenic avian influenza virus and velogenic viscerotropic Newcastle disease virus. Virus Res. 2016, 225, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Lam, K.M. Growth of Newcastle disease virus in chicken macrophages. J. Comp. Pathol. 1996, 115, 253–263. [Google Scholar] [CrossRef]
- Butt, S.L.; Moura, V.; Susta, L.; Miller, P.J.; Hutcheson, J.M.; Cardenas-Garcia, S.; Brown, C.C.; West, F.D.; Afonso, C.L.; Stanton, J.B. Tropism of Newcastle disease virus strains for chicken neurons, astrocytes, oligodendrocytes, and microglia. BMC Vet. Res. 2019, 15, 317. [Google Scholar] [CrossRef] [Green Version]
- Xiang, B.; Zhu, W.; Li, Y.; Gao, P.; Liang, J.; Liu, D.; Ding, C.; Liao, M.; Kang, Y.; Ren, T. Immune responses of mature chicken bone-marrow-derived dendritic cells infected with Newcastle disease virus strains with differing pathogenicity. Arch. Virol. 2018, 163, 1407–1417. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Shi, Q.K.; Han, Z.X.; Fan, Z.; Ai, H.; Chen, L.N.; Li, L.; Liu, T.Y.; Sun, J.F.; Liu, S.W. Newcastle disease virus entry into chicken macrophages via a pH-dependent, dynamin and caveola-mediated endocytic pathway that requires Rab5. J. Virol. 2021, 95, e02288-20. [Google Scholar] [CrossRef]
- Susta, L.; Miller, P.J.; Afonso, C.L.; Brown, C.C. Clinicopathological characterization in poultry of three strains of Newcastle disease virus isolated from recent outbreaks. Vet. Pathol. 2011, 48, 349–360. [Google Scholar] [CrossRef]
- Brown, C.; King, D.J.; Seal, B.S. Pathogenesis of Newcastle disease in chickens experimentally infected with viruses of different virulence. Vet. Pathol. 1999, 36, 125–132. [Google Scholar] [CrossRef]
- Zhang, P.Z.; Ding, Z.; Liu, X.X.; Chen, Y.Y.; Li, J.J.; Tao, Z.; Fei, Y.D.; Xue, C.; Qian, J.; Wang, X.L.; et al. Enhanced replication of virulent Newcastle disease virus in chicken macrophages is due to polarized activation of cells by inhibition of TLR7. Front. Immunol. 2018, 9, 366. [Google Scholar] [CrossRef]
- Cornax, I.; Diel, D.G.; Rue, C.A.; Estevez, C.; Yu, Q.; Miller, P.J.; Afonso, C.L. Newcastle disease virus fusion and haemagglutinin-neuraminidase proteins contribute to its macrophage host range. J. Gen. Virol. 2013, 94, 1189–1194. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.J.; Decanini, E.L.; Afonso, C.L. Newcastle disease: Evolution of genotypes and the related diagnostic challenges. Infect. Genet. Evol. 2010, 10, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.F.; Wan, H.Q.; Ni, X.X.; Wu, Y.T.; Liu, W.B. Pathotypical and genotypical characterization of strains of Newcastle disease virus isolated from outbreaks in chicken and goose flocks in some regions of China during 1985–2001. Arch. Virol. 2003, 148, 1387–1403. [Google Scholar] [CrossRef]
- Mase, M.; Imai, K.; Sanada, Y.; Sanada, N.; Yuasa, N.; Imada, T.; Tsukamoto, K.; Yamaguchi, S. Phylogenetic analysis of Newcastle disease virus genotypes isolated in Japan. J. Clin. Microbiol. 2002, 40, 3826–3830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Z.; Hu, J.; Hu, S.; Song, Q.; Ding, P.; Zhu, J.; Liu, X.; Wang, X.; Liu, X. High levels of virus replication and an intense inflammatory response contribute to the severe pathology in lymphoid tissues caused by Newcastle disease virus genotype VIId. Arch. Virol. 2015, 160, 639–648. [Google Scholar] [CrossRef]
- Ecco, R.; Brown, C.; Susta, L.; Cagle, C.; Cornax, I.; Pantin-Jackwood, M.; Miller, P.J.; Afonso, C.L. In vivo transcriptional cytokine responses and association with clinical and pathological outcomes in chickens infected with different Newcastle disease virus isolates using formalin-fixed paraffin-embedded samples. Vet. Immunol. Immunopathol. 2011, 141, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Rasoli, M.; Yeap, S.K.; Tan, S.W.; Moeini, H.; Ideris, A.; Bejo, M.H.; Alitheen, N.B.; Kaiser, P.; Omar, A.R. Alteration in lymphocyte responses, cytokine and chemokine profiles in chickens infected with genotype VII and VIII velogenic Newcastle disease virus. Comp. Immunol. Microbiol. Infect. Dis. 2014, 37, 11–21. [Google Scholar] [CrossRef]
- Hu, Z.; Hu, S.; Meng, C.; Wang, X.; Zhu, J.; Liu, X. Generation of a genotype VII Newcastle disease virus vaccine candidate with high yield in embryonated chicken eggs. Avian Dis. 2011, 55, 391–397. [Google Scholar] [CrossRef]
- Reddy, V.R.; Trus, I.; Desmarets, L.M.; Li, Y.; Theuns, S.; Nauwynck, H.J. Productive replication of nephropathogenic infectious bronchitis virus in peripheral blood monocytic cells, a strategy for viral dissemination and kidney infection in chickens. Vet. Res. 2016, 47, 70. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Liang, Y.; Wang, N.; Cui, L.; Chen, Z.; Wu, H.; Zhu, C.; Wang, Z.; Liu, S.; Li, H. Avian flavivirus infection of monocytes/macrophages by extensive subversion of host antiviral innate immune responses. J. Virol. 2019, 93, e00978-19. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H.A. A simple method of estimating fifty percent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Anis, Z.; Morita, T.; Azuma, K.; Ito, H.; Ito, T.; Shimada, A. Histopathological alterations in immune organs of chickens and ducks after experimental infection with virulent 9a5b newcastle disease virus. J. Comp. Pathol. 2013, 149, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Hamid, H.; Campbell, R.S.; Parede, L. Studies of the pathology of velogenic Newcastle disease: Virus infection in non-immune and immune birds. Avian Pathol. 1991, 20, 561–575. [Google Scholar] [CrossRef] [PubMed]
- Kameka, A.M.; Haddadi, S.; Jamaldeen, F.J.; Moinul, P.; He, X.T.; Nawazdeen, F.H.P.; Bonfield, S.; Sharif, S.; van Rooijen, N.; Abdul-Careem, M.F. Clodronate treatment significantly depletes macrophages in chickens. Can. J. Vet. Res. 2014, 78, 274–282. [Google Scholar] [PubMed]
- Abdul-Cader, M.S.; Ehiremen, G.; Nagy, E.; Abdul-Careem, M.F. Low pathogenic avian influenza virus infection increases the staining intensity of KUL01+cells including macrophages yet decrease of the staining intensity of KUL01+cells using clodronate liposomes did not affect the viral genome loads in chickens. Vet. Immunol. Immunopathol. 2018, 198, 37–43. [Google Scholar] [CrossRef]
- Coleman, C.M.; Sisk, J.M.; Halasz, G.; Zhong, J.; Beck, S.E.; Matthews, K.L.; Venkataraman, T.; Rajagopalan, S.; Kyratsous, C.A.; Frieman, M.B. CD8+ T cells and macrophages regulate pathogenesis in a mouse model of Middle East respiratory syndrome. J. Virol. 2017, 91, e01825-16. [Google Scholar] [CrossRef] [Green Version]
- Rivas, C.; Djeraba, A.; Musset, E.; van Rooijen, N.; Baaten, B.; Quéré, P. Intravenous treatment with liposome-encapsulated dichloromethylene bisphosphonate (Cl2MBP) suppresses nitric oxide production and reduces genetic resistance to Marek’s disease. Avian Pathol. 2003, 32, 139–149. [Google Scholar]
- Tate, M.D.; Pickett, D.L.; van Rooijen, N.; Brooks, A.G.; Reading, P.C. Critical role of airway macrophages in modulating disease severity during influenza virus infection of mice. J. Virol. 2010, 84, 7569–7580. [Google Scholar] [CrossRef] [Green Version]
- Roscic-Mrkic, B.; Schwendener, R.A.; Odermatt, B.; Zuniga, A.; Pavlovic, J.; Billeter, M.A.; Cattaneo, R. Roles of macrophages in measles virus infection of genetically modified mice. J. Virol. 2001, 75, 3343–3351. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Duan, Z.; Hu, S.; Kai, Y.; Wang, X.; Song, Q.; Zhong, L.; Sun, Q.; Wu, Y.; Liu, X. Lack of detection of host associated differences in Newcastle disease viruses of genotype VIId isolated from chickens and geese. Virol. J. 2012, 9, 197. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Ratio of macrophages in chicken PBMCs and NDV infectivity in different fractions of PBMCs. (A) Percentage of KUL01-positive cells in chicken PBMCs. The adherent cells were isolated from chicken PBMCs and the percentage of KUL01-positive cells in the adherent cells was determined using flow cytometry. Left panel: unlabeled cells; right panel, KUL01-positive cells. (B,C) NDV replication/transcription in the adherent and non-adherent cells derived from PBMCs. (B) JS5/05; (C) Herts/33. PBMCs were inoculated with NDV at a 0.01 multiplicity of infection, and at 8, 16 and 24 post-inoculation, the adherent and non-adherent cells were separated. The fusion gene copy was measured in these two cell fractions using quantitative real-time PCR. The data are presented as the mean values ± standard error of three independent experiments. Asterisk (*) stands for a significant difference (p < 0.05).
Figure 1.
Ratio of macrophages in chicken PBMCs and NDV infectivity in different fractions of PBMCs. (A) Percentage of KUL01-positive cells in chicken PBMCs. The adherent cells were isolated from chicken PBMCs and the percentage of KUL01-positive cells in the adherent cells was determined using flow cytometry. Left panel: unlabeled cells; right panel, KUL01-positive cells. (B,C) NDV replication/transcription in the adherent and non-adherent cells derived from PBMCs. (B) JS5/05; (C) Herts/33. PBMCs were inoculated with NDV at a 0.01 multiplicity of infection, and at 8, 16 and 24 post-inoculation, the adherent and non-adherent cells were separated. The fusion gene copy was measured in these two cell fractions using quantitative real-time PCR. The data are presented as the mean values ± standard error of three independent experiments. Asterisk (*) stands for a significant difference (p < 0.05).
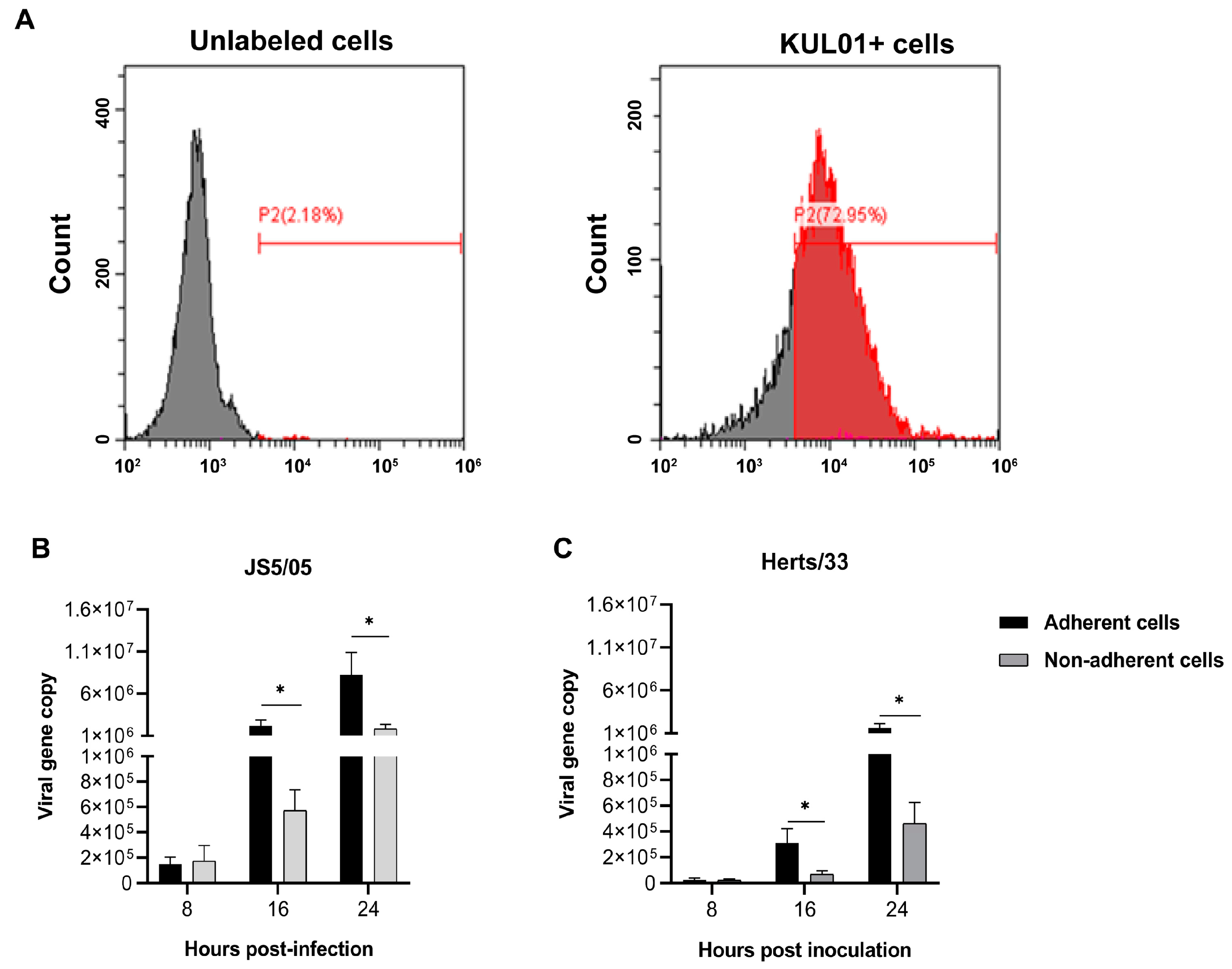
Figure 2.
NDV tropism to macrophages in the spleen. (A–C) Replication of NDV in the spleen of chickens. (A) The non-infected mock chickens; (B) the JS5/05-infected chickens; (C) the Herts/33-infected chickens. Chickens were infected with JS5/05 or Herts/33, and on day 3 post-infection, the spleens were harvested for preparation of tissue sections. Virus replication in the spleen was assessed using immunofluorescence assays. The NP protein of NDV (green) and the marker of chicken macrophages KUL01 (magenta) were detected, and the cell nucleus was stained with DAPI (blue). The entire spleen sections were scanned, and representative fields of the spleen sections of three chickens are shown. Scale bar, 200 µm. (D) Virus tropism to macrophages. The NP antigen and KUL01 signals in the representative regions in the spleen from the NDV-infected chickens are shown. Co-localization of the NP and KUL01 was observed. Co-localization of the NP and KUL01 is indicated by arrows. Scale bar, 10 µm. (E) Co-localization profiles of the NP and KUL01 in JS5/05-infected spleen; (F) co-localization profiles of the NP and KUL01 in Herts/33-infected spleen.
Figure 2.
NDV tropism to macrophages in the spleen. (A–C) Replication of NDV in the spleen of chickens. (A) The non-infected mock chickens; (B) the JS5/05-infected chickens; (C) the Herts/33-infected chickens. Chickens were infected with JS5/05 or Herts/33, and on day 3 post-infection, the spleens were harvested for preparation of tissue sections. Virus replication in the spleen was assessed using immunofluorescence assays. The NP protein of NDV (green) and the marker of chicken macrophages KUL01 (magenta) were detected, and the cell nucleus was stained with DAPI (blue). The entire spleen sections were scanned, and representative fields of the spleen sections of three chickens are shown. Scale bar, 200 µm. (D) Virus tropism to macrophages. The NP antigen and KUL01 signals in the representative regions in the spleen from the NDV-infected chickens are shown. Co-localization of the NP and KUL01 was observed. Co-localization of the NP and KUL01 is indicated by arrows. Scale bar, 10 µm. (E) Co-localization profiles of the NP and KUL01 in JS5/05-infected spleen; (F) co-localization profiles of the NP and KUL01 in Herts/33-infected spleen.
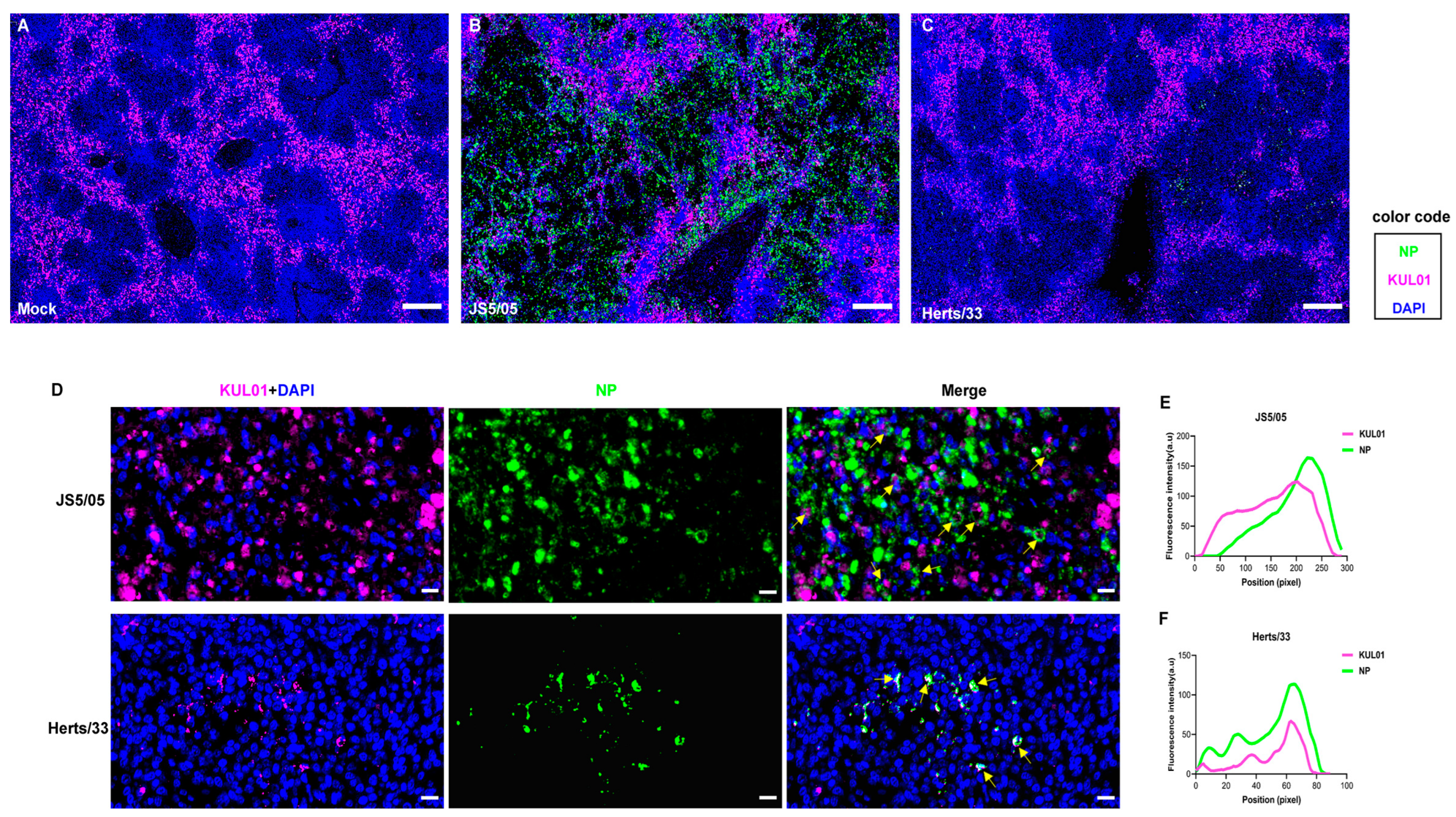
Figure 3.
Impact of macrophage depletion on NDV pathogenicity and replication in chickens. (A) Efficiency of macrophage depletion in the spleen. Chickens were treated with clodronate liposomes (CL lipo) or PBS liposomes (PBS lipo), and on day 5 post-treatment, the percentage of macrophages in the spleen was determined to determine the efficiency of macrophage depletion. Red, KUL01; blue, cell nucleus. Scale bar, 10 µm. (B) Survival of the chickens treated with CL lipo or PBS lipo after NDV infection. On day 2 after the treatment with CL lipo or PBS lipo, chickens were inoculated with JS5/05 and animal survival was determined. (C,D) Virus load in the CL lipo- and PBS lipo-treated chickens. The liver, spleen, lung, intestine and thymus were collected on days 1 (C) and 3 (D) post-infection for measurement of viral load. The dotted lines indicate the detection limit of the virus titration assay (1.0 log10 TCID50/g). The data of virus load are presented as the mean values ± standard error of three independent experiments. ns, non-significant differences.
Figure 3.
Impact of macrophage depletion on NDV pathogenicity and replication in chickens. (A) Efficiency of macrophage depletion in the spleen. Chickens were treated with clodronate liposomes (CL lipo) or PBS liposomes (PBS lipo), and on day 5 post-treatment, the percentage of macrophages in the spleen was determined to determine the efficiency of macrophage depletion. Red, KUL01; blue, cell nucleus. Scale bar, 10 µm. (B) Survival of the chickens treated with CL lipo or PBS lipo after NDV infection. On day 2 after the treatment with CL lipo or PBS lipo, chickens were inoculated with JS5/05 and animal survival was determined. (C,D) Virus load in the CL lipo- and PBS lipo-treated chickens. The liver, spleen, lung, intestine and thymus were collected on days 1 (C) and 3 (D) post-infection for measurement of viral load. The dotted lines indicate the detection limit of the virus titration assay (1.0 log10 TCID50/g). The data of virus load are presented as the mean values ± standard error of three independent experiments. ns, non-significant differences.
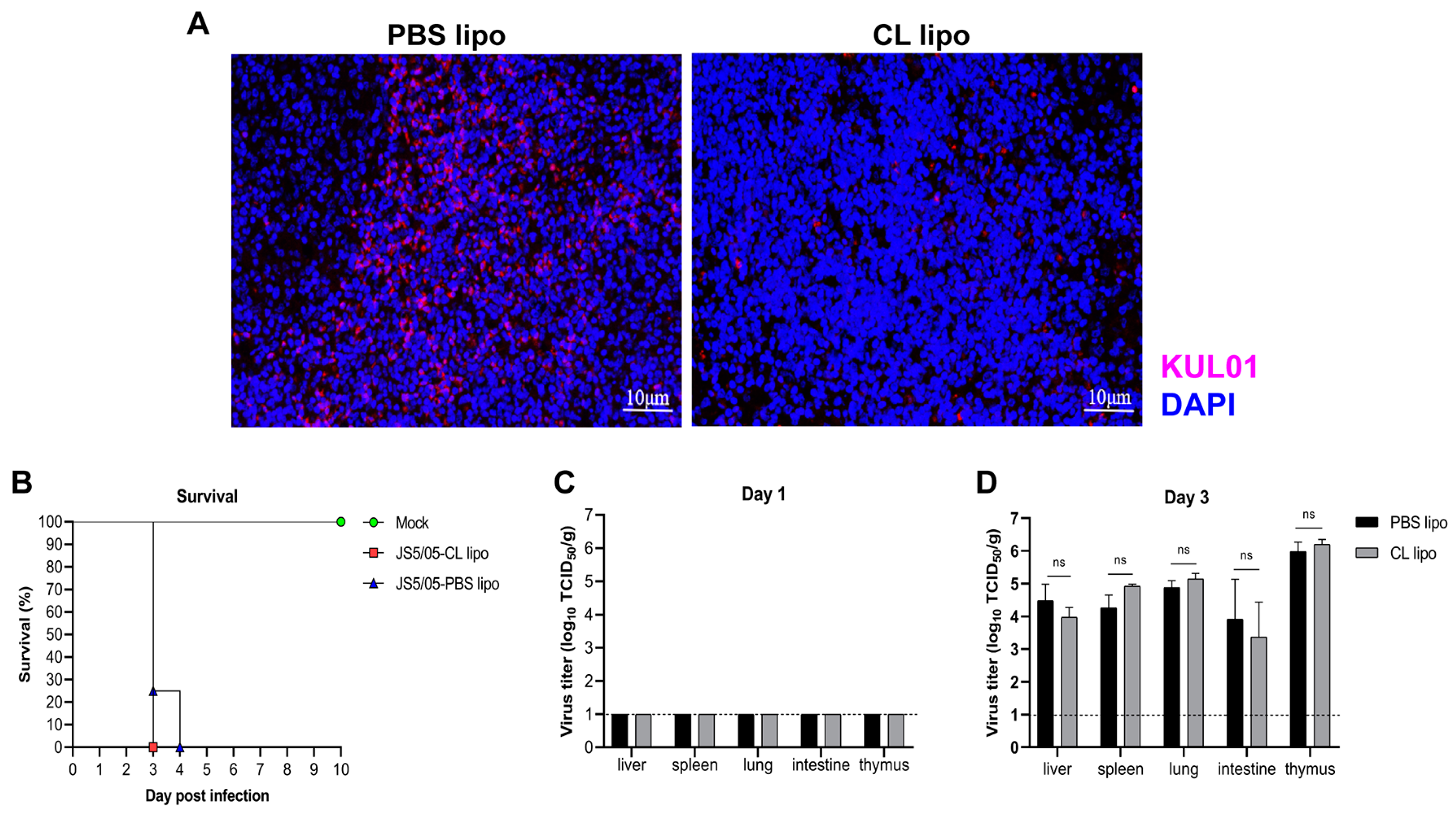
Figure 4.
Histopathological changes caused by NDV in chicken spleens after macrophage depletion. (A) Pathological lesions of the spleens. The spleens of the NDV-infected chickens were collected and prepared into tissue sections, which were subjected to H&E staining. Upper panel, day 1 post-infection; lower panel, day 3 post-infection. Arrows indicate necrotic foci in the spleen. Scale bar, 100 µm. (B) Quantitation of the area of necrotic foci in the spleens. The area of necrosis in the spleens collected on day 3 post-infection was calculated using ImageJ software (1.52V). Asterisk (*) stands for a significant difference (p < 0.05).
Figure 4.
Histopathological changes caused by NDV in chicken spleens after macrophage depletion. (A) Pathological lesions of the spleens. The spleens of the NDV-infected chickens were collected and prepared into tissue sections, which were subjected to H&E staining. Upper panel, day 1 post-infection; lower panel, day 3 post-infection. Arrows indicate necrotic foci in the spleen. Scale bar, 100 µm. (B) Quantitation of the area of necrotic foci in the spleens. The area of necrosis in the spleens collected on day 3 post-infection was calculated using ImageJ software (1.52V). Asterisk (*) stands for a significant difference (p < 0.05).
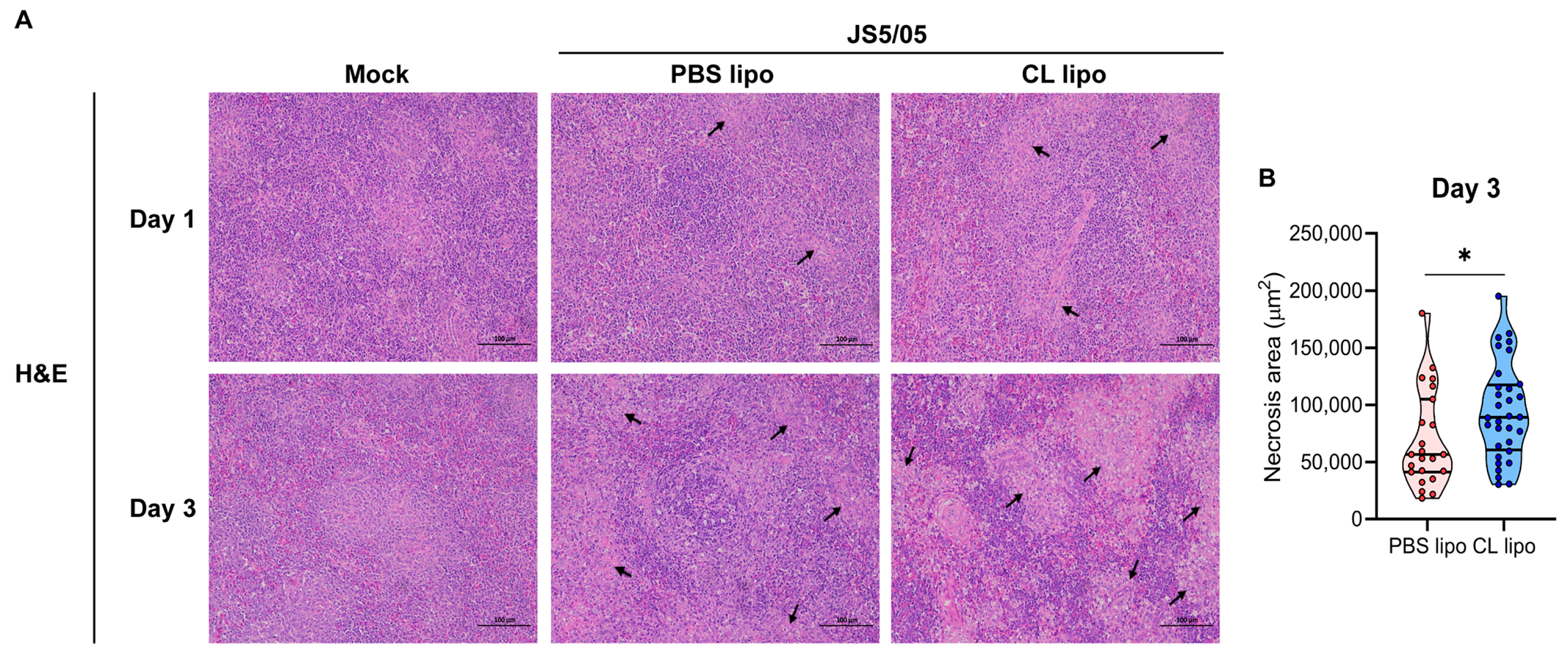
Figure 5.
Apoptosis in the spleens of NDV-infected chickens after macrophage depletion. The spleen sections were used for TUNEL staining to detect apoptotic cells. Apoptotic cells and cell nuclei are shown in green and blue fluorescence, respectively. Upper panel, day 1 post-infection; lower panel, day 3 post-infection. Scale bar, 1 µm.
Figure 5.
Apoptosis in the spleens of NDV-infected chickens after macrophage depletion. The spleen sections were used for TUNEL staining to detect apoptotic cells. Apoptotic cells and cell nuclei are shown in green and blue fluorescence, respectively. Upper panel, day 1 post-infection; lower panel, day 3 post-infection. Scale bar, 1 µm.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Scoring of the histopathology changes of the spleen.
| Group | dpi a | Lesion Severity b |
|---|---|---|
| PBS liposomes | 1 | + |
| 3 | +++ | |
| Clodronate liposomes | 1 | ++ |
| 3 | ++++ | |
| Control | 1 | − |
| 3 | − |
a dpi: day post infection; b scoring standards for tissue lesions: −, normal; +, mild to moderate lymphocyte proliferation; ++, moderate to marked lymphocyte depletion and necrosis; +++, marked lymphocyte depletion, necrosis and tissue hyperplasia; ++++, extensive and severe lymphocyte depletion and necrosis.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ni, J.; Deng, J.; Chen, Q.; Liao, T.; Hu, J.; Chen, Y.; Hu, S.; Hu, Z.; Liu, X. Role of Macrophages in the Pathogenesis of Genotype VII Newcastle Disease Virus in Chickens. Animals 2023, 13, 2239. https://doi.org/10.3390/ani13132239
AMA Style
Ni J, Deng J, Chen Q, Liao T, Hu J, Chen Y, Hu S, Hu Z, Liu X. Role of Macrophages in the Pathogenesis of Genotype VII Newcastle Disease Virus in Chickens. Animals. 2023; 13(13):2239. https://doi.org/10.3390/ani13132239
Chicago/Turabian StyleNi, Jie, Jing Deng, Qing Chen, Tianxing Liao, Jiao Hu, Yu Chen, Shunlin Hu, Zenglei Hu, and Xiufan Liu. 2023. "Role of Macrophages in the Pathogenesis of Genotype VII Newcastle Disease Virus in Chickens" Animals 13, no. 13: 2239. https://doi.org/10.3390/ani13132239
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.