
Time to Step Up Conservation: Climate Change Will Further Reduce the Suitable Habitats for the Vulnerable Species Marbled Polecat (Vormela peregusna)
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
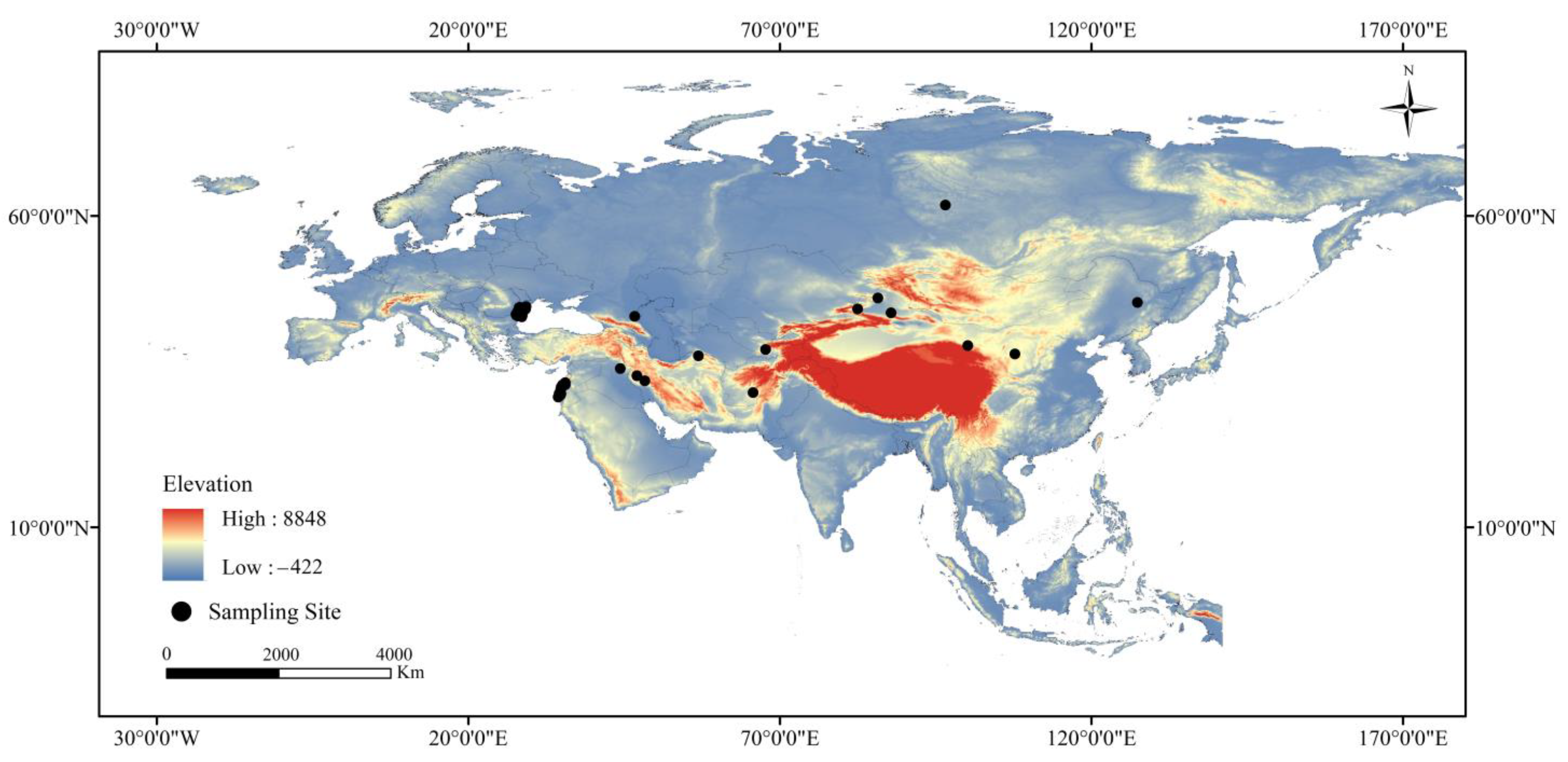
2.1. Occurrence Data
2.2. Selection and Processing of Environmental Variables
2.3. Model Construction
2.4. Changes in the Spatial Pattern of the Suitable Distribution Ranges of Species
3. Results
3.1. Model Accuracy
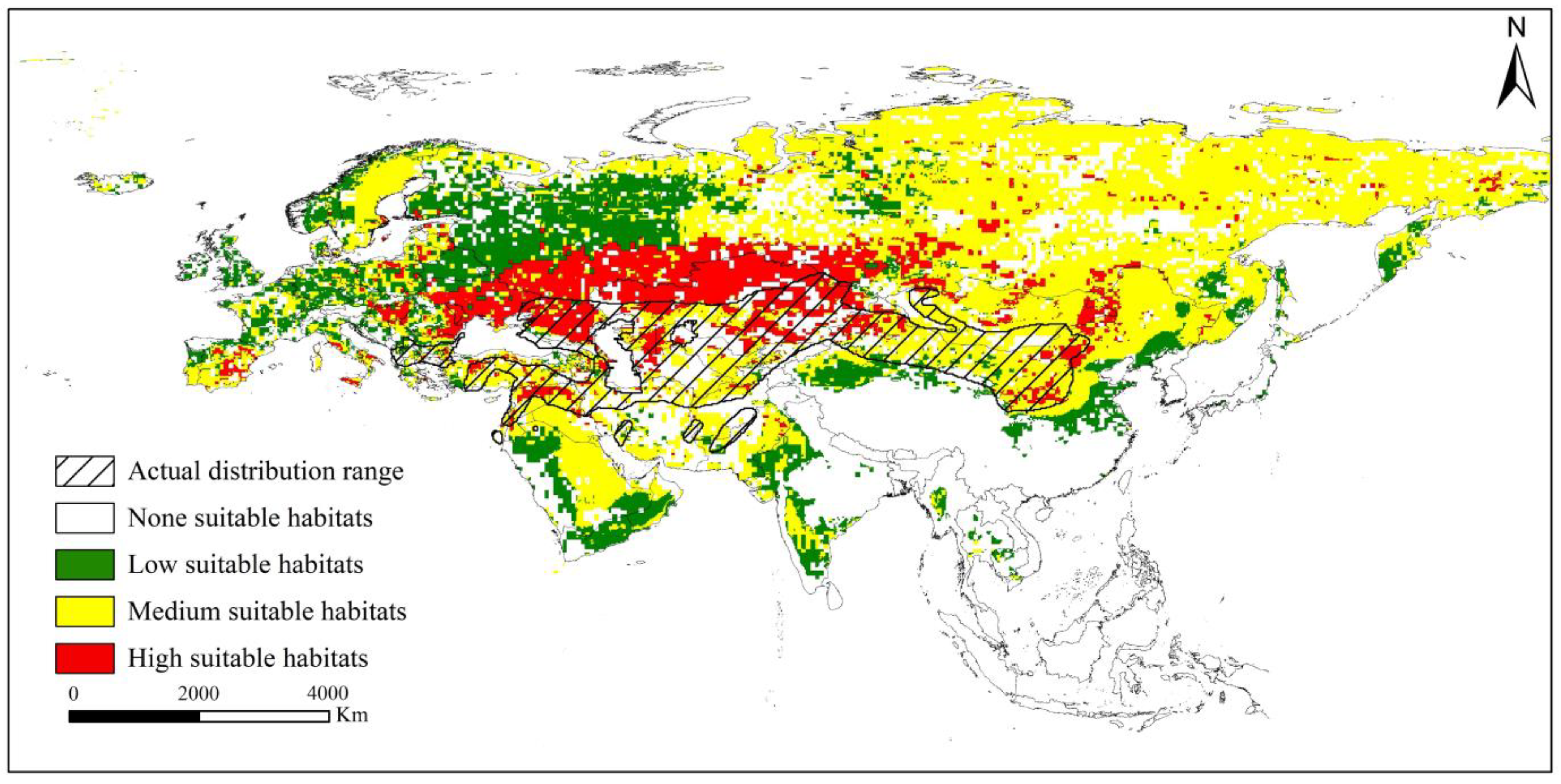
3.2. Current Distribution Range
3.3. Future Changes in Suitable Habitat Area
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thapa, A.; Wu, R.; Hu, Y.; Nie, Y.; Singh, P.B.; Khatiwada, J.R.; Yan, L.; Gu, X.; Wei, F. Predicting the potential distribution of the endangered red panda across its entire range using MaxEnt modeling. Ecol. Evol. 2018, 8, 10542–10554. [Google Scholar] [CrossRef] [Green Version]
- Scherrer, D.; Christe, P.; Guisan, A. Modelling bat distributions and diversity in a mountain landscape using focal predictors in ensemble of small models. Divers. Distrib. 2019, 25, 770–782. [Google Scholar] [CrossRef] [Green Version]
- International Union for Conservation of Nature. The IUCN Red List of Threatened Species. Version 2022-2. Available online: https://www.iucnredlist.org (accessed on 15 December 2022).
- Hoekstra, J.M.; Boucher, T.M.; Ricketts, T.H.; Roberts, C. Confronting a biome crisis: Global disparities of habitat loss and protection. Ecol. Lett. 2005, 8, 23–29. [Google Scholar] [CrossRef]
- He, F.; Hubbell, S.P. Species-area relationships always overestimate extinction rates from habitat loss. Nature 2011, 473, 368–371. [Google Scholar] [CrossRef] [PubMed]
- Coristine, L.E.; Kerr, J.T. Habitat loss, climate change, and emerging conservation challenges in Canada. Can. J. Zool. 2011, 89, 435–451. [Google Scholar] [CrossRef]
- Galante, P.J.; Triguero, S.C.; Paz, A.; Aiello-Lammens, M.; Gerstner, B.E.; Johnson, B.A.; Kass, J.M.; Merow, C.; Noguera-Urbano, E.A.; Pinilla-Buitrago, G.E.; et al. changeRangeR: An R package for reproducible biodiversity change metrics from species distribution estimates. Conserv. Sci. Pract. 2022, 5, e12863. [Google Scholar] [CrossRef]
- Brodie, S.; Smith, J.A.; Muhling, B.A.; Barnett, L.A.K.; Carroll, G.; Fiedler, P.; Bograd, S.J.; Hazen, E.L.; Jacox, M.G.; Andrews, K.S.; et al. Recommendations for quantifying and reducing uncertainty in climate projections of species distributions. Glob. Chang. Biol. 2022, 28, 6586–6601. [Google Scholar] [CrossRef] [PubMed]
- Hodel, R.G.J.; Soltis, D.E.; Soltis, P.S. Hindcast-validated species distribution models reveal future vulnerabilities of mangroves and salt marsh species. Ecol. Evol. 2022, 12, e9252. [Google Scholar] [CrossRef]
- Norberg, A.; Abrego, N.; Blanchet, F.G.; Adler, F.R.; Anderson, B.J.; Anttila, J.; Araújo, M.B.; Dallas, T.; Dunson, D.; Elith, J.; et al. A comprehensive evaluation of predictive performance of 33 species distribution models at species and community levels. Ecol. Monogr. 2019, 89, e01370. [Google Scholar] [CrossRef]
- Tez, C.; Gündüz, I.; Kefelioğlu, H. Contributions to distribution, reproduction biology and ecology of Vormela peregusna (Güldenstaedt, 1770) (Mammalia: Carnivora) in Turkey. Pak. J. Biol. Sci. 2001, 4, 74–76. [Google Scholar] [CrossRef]
- Gama, M.; Crespo, D.; Dolbeth, M.; Anastácio, P. Predicting global habitat suitability for Corbicula fluminea using species distribution models: The importance of different environmental datasets. Ecol. Model. 2016, 319, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Di Cola, V.; Broennimann, O.; Petitpierre, B.; Breiner, F.T.; D’Amen, M.; Randin, C.; Engler, R.; Pottier, J.; Pio, D.; Dubuis, A.; et al. ecospat: An R package to support spatial analyses and modeling of species niches and distributions. Ecography 2017, 40, 774–787. [Google Scholar] [CrossRef]
- Hao, T.; Elith, J.; Lahoz-Monfort, J.J.; Guillera-Arroita, G. Testing whether ensemble modelling is advantageous for maximising predictive performance of species distribution models. Ecography 2020, 43, 549–558. [Google Scholar] [CrossRef] [Green Version]
- Schickele, A.; Leroy, B.; Beaugrand, G.; Goberville, E.; Hattab, T.; Francour, P.; Raybaud, V. Modelling European small pelagic fish distribution: Methodological insights. Ecol. Model. 2020, 416, 108902. [Google Scholar] [CrossRef]
- Puzachenko, A.Y.; Abramov, A.V.; Rozhnov, V.V. Cranial variation and taxonomic content of the marbled polecat Vormela peregusna (Mustelidae, Carnivora). Mamm. Biol. 2017, 83, 10–20. [Google Scholar] [CrossRef]
- Ben-David, M.; Pellis, S.M.; Pellis, V.C. Feeding habits and predatory behaviour in the marbled polecat (Vormela peregusna syriaca): I. Killing methods in relation to prey size and prey behaviour. Behaviour 1991, 118, 127–143. [Google Scholar] [CrossRef]
- Liu, G.; Zhao, S.; Tan, W.; Hornok, S.; Yuan, W.; Mi, L.; Wang, S.; Liu, Z.; Zhang, Y.; Hazihan, W.; et al. Rickettsiae in red fox (Vulpes vulpes), marbled polecat (Vormela peregusna) and their ticks in northwestern China. Parasit. Vectors 2021, 14, 204. [Google Scholar] [CrossRef]
- Abramov, A.V.; Kranz, A.; Maran, T. Vormela peregusna. In The IUCN Red List of Threatened Species 2016; International Union for Conservation of Nature and Natural Resources: Gland, Switzerland, 2016. [Google Scholar] [CrossRef]
- Zidarova, S. Present distribution of the marble polecat Vormela peregusna (Güldenstädt, 1770) (Carnivora: Mustelidae) in central Western Bulgaria, with an observation on its defensive behavior. Acta Zool. Bulg. 2022, 74, 403–408. [Google Scholar]
- Wright, P.G.R.; Croose, E.; Hunter, S.B.; MacPherson, J.; Çoraman, E.; Yarotskiy, V.; Moisieieva, V.; Karapandža, B.; Hoxha, B.; Madalina, P.; et al. Can social media be used to inform the distribution of the marbled polecat, Vormela peregusna? Mammal Res. 2023, 68, 295–304. [Google Scholar] [CrossRef]
- Ivanov, V.; Spassov, N. Some new data on the distribution, habitats and ecology of the threatened European mustelids Mustela eversmanii and Vormela peregusna in Bulgaria. Hist. Nat. Bulg. 2015, 21, 267–271. [Google Scholar]
- AL-Sheikhly, O.S.; Ararat, K.; Ahmed, S.H. New records for the first time in six decades of the Vulnerable Marbled Polecat Vormela peregusna in Iraq. Small Carniv. Conserv. 2022, 60, e60004. [Google Scholar]
- Dulamtseren, S.; Shar, S.; Murdoch, J.; Reading, R.P.; Gantulga, J.; Usukhjrargal, D.; Suuri, B. Contributions to the distribution of the marbled polecat (Vormela peregusna Guldenstaedt, 1770) in Mongolia. Small Carniv. Conserv. 2009, 40, 29–32. [Google Scholar]
- Wen, X.; Cheng, X.; Dong, Y.; Wang, Q.; Lin, X. Analysis of the activity rhythms of the great gerbil (Rhombomys opimus) and its predators and their correlations based on infrared camera technology. Glob. Ecol. Conserv. 2020, 24, e01337. [Google Scholar] [CrossRef]
- Werner, N.Y. Small carnivores, big database—Inferring possible small carnivore distribution and population trends in Israel from over 30 years of recorded sightings. Small Carniv. Conserv. 2012, 47, 17–25. [Google Scholar]
- He, B.; Tuya, W.; Qinchaoketu, S.; Nanzad, L.; Yong, M.; Kesi, T.; Sun, C. Climate Change Characteristics of Typical Grassland in the Mongolian Plateau from 1978 to 2020. Sustainability 2022, 14, 16529. [Google Scholar] [CrossRef]
- Vale, C.G.; Brito, J.C. Desert-adapted species are vulnerable to climate change: Insights from the warmest region on Earth. Glob. Ecol. Conserv. 2015, 4, 369–379. [Google Scholar] [CrossRef] [Green Version]
- Iknayan, K.J.; Beissinger, S.R. Collapse of a desert bird community over the past century driven by climate change. Proc. Natl. Acad. Sci. USA 2018, 115, 8597–8602. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Bao, A.; Jiapaer, G.; Guo, H.; Zheng, G.; Jiang, L.; Chang, C.; Tuerhanjiang, L. Disentangling the relative impacts of climate change and human activities on arid and semiarid grasslands in central asia during 1982–2015. Sci. Total Environ. 2019, 53, 1311–1325. [Google Scholar] [CrossRef]
- Gherman, C.M.; Sándor, A.D.; Kalmár, Z.; Marinov, M.; Mihalca, A.D. First report of Borrelia burgdorferi sensu lato in two threatened carnivores: The Marbled polecat, Vormela peregusna and the European mink, Mustela lutreola (Mammalia: Mustelidae). BMC Vet. Res. 2012, 8, 137. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Yang, M.; Liu, G.; Zhao, S.; Yuan, W.; Xiao, R.; Hazihan, W.; Hornok, S.; Wang, Y. Molecular evidence of Rickettsia raoultii, “Candidatus Rickettsia barbariae” and a novel Babesia genotype in marbled polecats (Vormela peregusna) at the China-Kazakhstan border. Parasit. Vectors 2018, 11, 450. [Google Scholar] [CrossRef] [Green Version]
- Hadad, E.; Charter, M.; Kosicki, J.Z.; Yosef, R. Prey-base does not influence breeding success in eagle owls (Bubo bubo) in Judea, Israel. Animals 2022, 12, 1280. [Google Scholar] [CrossRef]
- Gray, A.; Brito, J.C.; Edwards, C.W.; Figueiró, H.V.; Koepfli, K.P. First complete mitochondrial genome of the Saharan striped polecat (Ictonyx libycus). Mitochondrial DNA Pt. B Resour. 2022, 7, 1957–1960. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Zhao, G.; Cheng, X.; Chang, G.; Dong, X.; Lin, X. Prediction of the potential distribution pattern of the great gerbil (Rhombomys opimus) under climate change based on ensemble modelling. Pest Manag. Sci. 2022, 78, 3128–3134. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, Y.; Shi, X.; Zhou, W.; Huang, A.; Mu, M.; Qiu, B.; Ji, J. Development of land surface model BCC_AVIM2.0 and its preliminary performance in LS3MIP/CMIP6. J. Meteorol. Res. 2019, 33, 851–869. [Google Scholar] [CrossRef]
- Graham, M.H. Confronting multicollinearity in ecological multiple regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef] [Green Version]
- Rose, E.L. Multivariate analysis of categorical data: Theory. Struct. Equ. Model. 1995, 2, 274–276. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Singh, K.; McClean, C.J.; Büker, P.; Hartley, S.E.; Hill, J.K. Mapping regional risks from climate change for rainfed rice cultivation in India. Agric. Syst. 2017, 156, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Fourcade, Y.; Besnard, A.G.; Secondi, J. Paintings predict the distribution of species, or the challenge of selecting environmental predictors and evaluation statistics. Glob. Ecol. Biogeogr. 2018, 27, 245–256. [Google Scholar] [CrossRef]
- Liu, C.; White, M.; Newell, G. Selecting thresholds for the prediction of species occurrence with presence-only data. J. Biogeogr. 2013, 40, 778–789. [Google Scholar] [CrossRef]
- Patasaraiya, M.K.; Devi, R.M.; Sinha, B.; Bisaria, J. Predicting impacts of climate change on teak and sal forests in central India using maximum entropy modeling: An approach for future conservation and silvicultural strategies. For. Sci. 2023, fxad014. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models: With Applications in R; Ecology, Biodiversity and Conservation); Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- Quintero, I.; Wiens, J.J. Rates of projected climate change dramatically exceed past rates of climatic niche evolution among vertebrate species. Ecol. Lett. 2013, 16, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- Christmas, M.J.; Breed, M.F.; Lowe, A.J. Constraints to and conservation implications for climate change adaptation in plants. Conserv. Genet. 2016, 17, 305–320. [Google Scholar] [CrossRef] [Green Version]
- Blair, M.E.; Le, M.D.; Xu, M. Species distribution modeling to inform transboundary species conservation and management under climate change: Promise and pitfalls. Front. Biogeogr. 2022, 14, e54662. [Google Scholar] [CrossRef]
- Ikeda, T.; Higashide, D.; Shichijo, T. Impact of human disturbance in Japan on the distribution and diel activity pattern of terrestrial mammals. J. Nat. Conserv. 2022, 70, 126293. [Google Scholar] [CrossRef]
- Kufa, C.A.; Bekele, A.; Atickem, A. Impacts of climate change on predicted habitat suitability and distribution of Djaffa Mountains Guereza (Colobus guereza gallarum, Neumann 1902) using MaxEnt algorithm in Eastern Ethiopian Highland. Glob. Ecol. Conserv. 2022, 35, e02094. [Google Scholar] [CrossRef]
- Baral, K.; Adhikari, B.; Bhandari, S.; Kunwar, R.M.; Sharma, H.P.; Aryal, A.; Ji, W. Impact of climate change on distribution of common leopard (Panthera pardus) and its implication on conservation and conflict in Nepal. Heliyon 2023, 9, e12807. [Google Scholar] [CrossRef]
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344, 1246752. [Google Scholar] [CrossRef]
- Guan, T.; Owens, J.R.; Kong, Y.; Jian, Y.; Gu, X.; Song, Y. The need for IUCN species distribution update—The case from takin (Budorcas taxicolor) in Southwest China. Ecol. Evol. 2022, 12, e9222. [Google Scholar] [CrossRef]
- Foden, W.B.; Butchart, S.H.; Stuart, S.N.; Vié, J.C.; Akçakaya, H.R.; Angulo, A.; DeVantier, L.M.; Gutsche, A.; Turak, E.; Cao, L.; et al. Identifying the world’s most climate change vulnerable species: A systematic trait-based assessment of all birds, amphibians and corals. PLoS ONE 2013, 8, e65427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheatley, C.J.; Beale, C.M.; Bradbury, R.B.; Pearce-Higgins, J.W.; Critchlow, R.; Thomas, C.D. Climate change vulnerability for species-assessing the assessments. Glob. Chang. Biol. 2017, 23, 3704–3715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akçakaya, H.R.; Butchart, S.H.M.; Watson, J.E.M.; Pearson, R.G. Preventing species extinctions resulting from climate change. Nat. Clim. Chang. 2014, 4, 1048–1049. [Google Scholar] [CrossRef]
- Wessely, J.; Hülber, K.; Gattringer, A.; Kuttner, M.; Moser, D.; Rabitsch, W.; Schindler, S.; Dullinger, S.; Essl, F. Habitat-based conservation strategies cannot compensate for climate-change-induced range loss. Nat. Clim. Chang. 2017, 7, 823–827. [Google Scholar] [CrossRef]
- Bonnot, T.W.; Cox, W.A.; Thompson, F.R.; Millspaugh, J.J. Threat of climate change on a songbird population through its impacts on breeding. Nat. Clim. Chang. 2018, 8, 718–722. [Google Scholar] [CrossRef]
- Liu, C.; White, M.; Newell, G. Measuring and comparing the accuracy of species distribution models with presence—Absence data. Ecography 2011, 34, 232–243. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Code | Environmental Variable | Variable Importance |
|---|---|---|
| bio16 | Precipitation in wettest quarter | 23.34 |
| t_caco3 | Topsoil calcium carbonate content | 14.46 |
| bio18 | Precipitation in warmest quarter | 13.94 |
| t_teb | Topsoil teb. | 12.36 |
| bio3 | Isothermality | 10.65 |
| elev | Elevation | 7.27 |
| t_cec_clay | Topsoil CEC (CLAY) | 4.77 |
| bio19 | Precipitation in coldest quarter | 4.26 |
| bio17 | Precipitation in driest quarter | 2.18 |
| bio15 | Precipitation seasonality | 2.04 |
| t_caco4 | Topsoil gypsum content | 1.00 |
| slope | Slope | 0.99 |
| bio5 | Max. temperature | 0.66 |
| t_ece | Topsoil salinity (Elco) | 0.56 |
| t_gravel | Topsoil gravel content | 0.43 |
| t_oc | Topsoil organic carbon | 0.41 |
| t_esp | Topsoil sodicity (ESP) | 0.37 |
| t-sand | Topsoil sand fraction | 0.30 |
| Periods | Climate Scenario | Suitable Habitat Area (×104 km2) | Loss (×104 km2) | Stable (×104 km2) | Gain (×104 km2) | Species Range Change (%) | Percentage Loss (%) | Percentage Gain (%) |
|---|---|---|---|---|---|---|---|---|
| Current | 3067.93 | |||||||
| 2050 | SSP126 | 2226.43 | 978.64 | 2089.29 | 137.14 | −27.43 | 31.90 | 4.47 |
| SSP245 | 2553.79 | 646.18 | 2421.75 | 132.04 | −16.76 | 21.06 | 4.30 | |
| SSP585 | 2226.66 | 926.29 | 2141.64 | 85.02 | −27.42 | 30.19 | 2.77 | |
| 2090 | SSP126 | 2702.43 | 546.16 | 2521.77 | 180.66 | −11.91 | 17.80 | 5.89 |
| SSP245 | 2038.78 | 1099.21 | 1968.72 | 70.06 | −33.55 | 35.83 | 2.28 | |
| SSP585 | 2213.48 | 978.59 | 2089.34 | 124.14 | −27.85 | 31.90 | 4.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, X.; Han, Y.; Lin, J.; Jiang, F.; Cai, Q.; Shi, Y.; Cui, D.; Wen, X. Time to Step Up Conservation: Climate Change Will Further Reduce the Suitable Habitats for the Vulnerable Species Marbled Polecat (Vormela peregusna). Animals 2023, 13, 2341. https://doi.org/10.3390/ani13142341
Cheng X, Han Y, Lin J, Jiang F, Cai Q, Shi Y, Cui D, Wen X. Time to Step Up Conservation: Climate Change Will Further Reduce the Suitable Habitats for the Vulnerable Species Marbled Polecat (Vormela peregusna). Animals. 2023; 13(14):2341. https://doi.org/10.3390/ani13142341
Chicago/Turabian StyleCheng, Xiaotian, Yamin Han, Jun Lin, Fan Jiang, Qi Cai, Yong Shi, Dongyang Cui, and Xuanye Wen. 2023. "Time to Step Up Conservation: Climate Change Will Further Reduce the Suitable Habitats for the Vulnerable Species Marbled Polecat (Vormela peregusna)" Animals 13, no. 14: 2341. https://doi.org/10.3390/ani13142341
APA StyleCheng, X., Han, Y., Lin, J., Jiang, F., Cai, Q., Shi, Y., Cui, D., & Wen, X. (2023). Time to Step Up Conservation: Climate Change Will Further Reduce the Suitable Habitats for the Vulnerable Species Marbled Polecat (Vormela peregusna). Animals, 13(14), 2341. https://doi.org/10.3390/ani13142341