
Avian Leukosis: Will We Be Able to Get Rid of It?

Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Avian Sarcoma/Leukosis Virus (ASLV) in the Past and in the Present
1.1.1. ASLV in History
1.1.2. Current Relevance of ALV
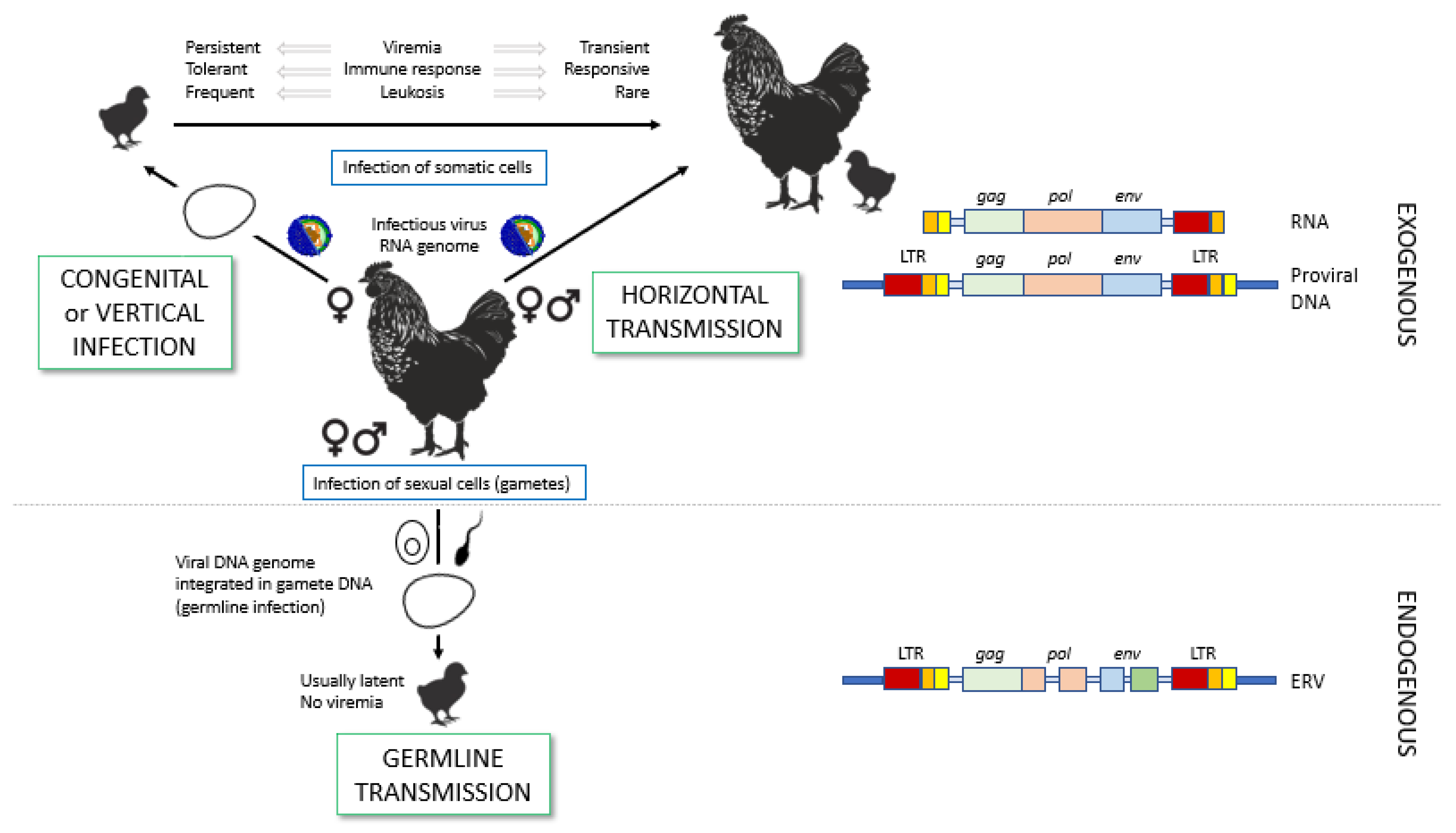
2. Transmission of ALV
2.1. Routes of Transmission: Horizontal Versus Vertical
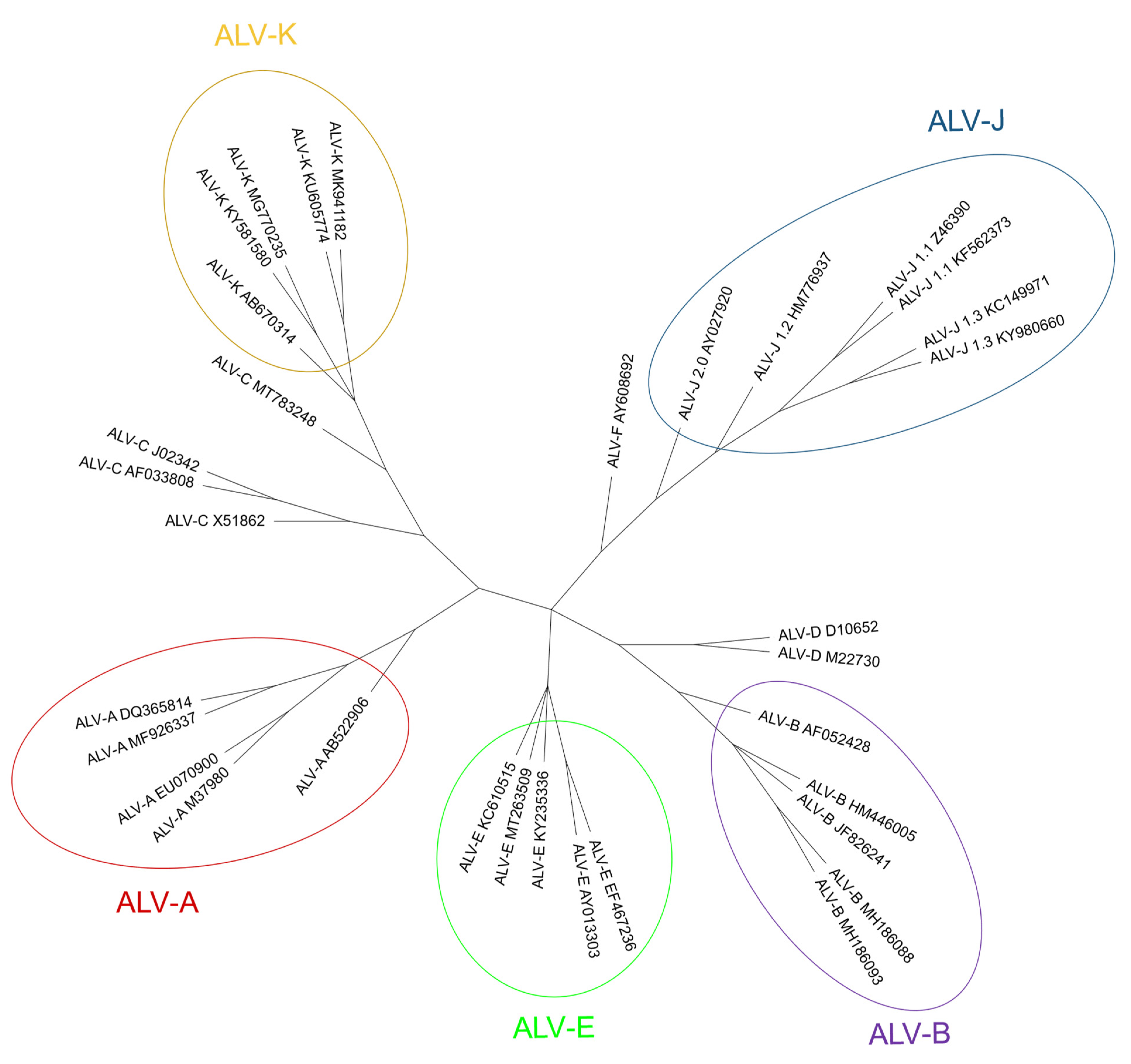
2.2. ALV Classification into Subgroups
2.2.1. Exogenous ALV Subgroups
2.2.2. Endogenous ALV Subgroup
3. The Avian Leukosis Virus, Its Characteristics and Properties
3.1. The ALV Genome
3.2. ALV Infection of the Cell
4. ALV Is Not Only Present in Chickens
4.1. Host Range
4.2. ALV Prevalence in Broilers and Layer Poultry (Commercial Poultry)
4.3. ALV Prevalence in Backyard, Fancy and Hobby Chickens (Non-Commercial Poultry)
4.4. ALV Prevalence in Non-Chicken Species
4.5. Endogenous ALVs in Non-Chicken Species
5. Different ALV Subgroups Use Different Cell Receptors

6. Immune Response against ALV Infection
6.1. Innate Immunity
6.2. Humoral Immune Response: The Role of Antibodies
6.3. Cellular Immune Response: The Role of T-Cell Dependent Cytotoxicity
7. What Can Be Done to Control ALV Infections?
7.1. Prevention through Vaccines
7.2. Selection of ALV-Free Poultry
7.3. Resistance to ALV through Manipulation of Bird Genomes
7.3.1. Boosting of Host Resistance by Cell Receptor Genetic Edition
7.3.2. Other Determinants as Targets for ALV Resistance
7.3.3. Role of ALV-E Provirus in ALV Resistance
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Payne, L.N.; Nair, V. The Long View: 40 Years of Avian Leukosis Research. Avian. Pathol. 2012, 41, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Zhou, D.; Meng, W.; Li, G.; Zhuang, P.; Pan, Z.; Wang, G.; Cheng, Z. Growth Retardation Induced by Avian Leukosis Virus Subgroup J Associated with Down-Regulated Wnt/β-Catenin Pathway. Microb. Pathog. 2017, 104, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Salter, D.; Balander, R.; Crittenden, L. Evaluation of Japanese Quail as a Model System for Avian Transgenesis Using Avian Leukosis Viruses. Poult. Sci. 1999, 78, 230–234. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, L.; Li, T.; Li, L.; Kan, Q.; Yao, X.; Xie, Q.; Wan, Z.; Shao, H.; Qin, A.; et al. Synergistic Pathogenesis of Chicken Infectious Anemia Virus and J Subgroup of Avian Leukosis Virus. Poult. Sci. 2021, 100, 101468. [Google Scholar] [CrossRef] [PubMed]
- Nair, V. Leukosis/Sarcoma Group. In Diseases of Poultry; Swayne, D.E., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; pp. 587–625. [Google Scholar]
- Ellermann, V.; Bang, O. Experimentelle Leukamie Bei Huhnern. Zentralbl. Bakteriol. Parasitenkd. Infectionskr. Hyg. Abt. Orig. 2008, 46, 596–609. [Google Scholar] [CrossRef]
- Rous, P. A Sarcoma of the Fowl Transmissible Byan Agent Separable from the Tumor Cells. J. Exp. Med. 1911, 13, 397–411. [Google Scholar] [CrossRef] [Green Version]
- Rubin, H.; Vogt, P.K. An Avian Leukosis Virus Asso-Ciated with Stocks of Rous Sarcoma Virus. Virology 1962, 17, 184–194. [Google Scholar] [CrossRef]
- Temin, H.M.; Rubin, H. Characteristics of an Assay for Rous Sarcoma Virus and Rous Sarcoma Cells in Tissue Culture. Virology 1958, 6, 669–688. [Google Scholar] [CrossRef]
- Rubin, H. The Early History of Tumor Virology: Rous, RIF, and RAV. Proc. Natl. Acad. Sci. USA 2011, 108, 14389–14396. [Google Scholar] [CrossRef]
- Temin, H.M.; Mizutani, S. Viral RNA-Dependent DNA Polymerase: RNA-Dependent DNA Polymerase in Virions of Rous Sarcoma Virus. Nature 1970, 226, 1211–1213. [Google Scholar] [CrossRef]
- Baltimore, D. Viral RNA-Dependent DNA Polymerase: RNA-Dependent DNA Polymerase in Virions of RNA Tumour Viruses. Nature 1970, 226, 1209–1211. [Google Scholar] [CrossRef] [PubMed]
- Engelbreth-Holm, J.; Meyer, A.R. On the Connection between Erythroblastosis (Haemocytoplastosis), Myelosis and Sarcoma in Chicken. Acta Pathol. Microbiol. Scand. 1935, 12, 352–365. [Google Scholar] [CrossRef]
- Coffin, J.M.; Hughes, S.H.; Varmus, H.E. A Brief Chronicle of Retrovirology. In Retroviruses; Cold Spring Harbor Laboratory Press: Woodbury, NY, USA, 1997; pp. 6–16. ISBN 0-87969-571-4. [Google Scholar]
- Frossard, J. Retroviridae. In Veterinary Microbiology; McVey, D.S., Kennedy, M., Chengappa, M.M., Wilkes, R., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2022; pp. 698–727. ISBN 978-1-119-65075-1. [Google Scholar]
- Malhotra, S.; Justice, J.; Lee, N.; Li, Y.; Zavala, G.; Ruano, M.; Morgan, R.; Beemon, K. Complete Genome Sequence of an American Avian Leukosis Virus Subgroup j Isolate That Causes Hemangiomas and Myeloid Leukosis. Genome Announc. 2015, 3, e01586-14. [Google Scholar] [CrossRef] [Green Version]
- Pandiri, A.R.; Gimeno, I.M.; Mays, J.K.; Reed, W.M.; Fadly, A.M. Reversion to Subgroup J Avian Leukosis Virus Viremia in Seroconverted Adult Meat-Type Chickens Exposed to Chronic Stress by Adrenocorticotrophin Treatment. Avian Dis. 2012, 56, 578–582. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.-P.; Teng, M.; Li, G.-X.; Zhang, W.-K.; Wang, W.-D.; Liu, J.-L.; Li, L.-Y.; Yao, Y.; Nair, V.; Luo, J. Current Epidemiology and Co-Infections of Avian Immunosuppressive and Neoplastic Diseases in Chicken Flocks in Central China. Viruses 2022, 14, 2599. [Google Scholar] [CrossRef]
- Dong, X.; Zhao, P.; Li, W.; Chang, S.; Li, J.; Li, Y.; Ju, S.; Sun, P.; Meng, F.; Liu, J.; et al. Diagnosis and Sequence Analysis of Avian Leukosis Virus Subgroup J Isolated from Chinese Partridge Shank Chickens. Poult. Sci. 2015, 94, 668–672. [Google Scholar] [CrossRef]
- Freick, M.; Schreiter, R.; Weber, J.; Vahlenkamp, T.W.; Heenemann, K. Avian Leukosis Virus (ALV) Is Highly Prevalent in Fancy-Chicken Flocks in Saxony. Arch. Virol. 2022, 167, 1169–1174. [Google Scholar] [CrossRef]
- Mo, G.; Wei, P.; Hu, B.; Nie, Q.; Zhang, X. Advances on Genetic and Genomic Studies of ALV Resistance. J. Anim. Sci. Biotechnol. 2022, 13, 123. [Google Scholar] [CrossRef]
- Deng, Q.; Li, Q.; Li, M.; Zhang, S.; Wang, P.; Fu, F.; Zhu, W.; Wei, T.; Mo, M.; Huang, T.; et al. The Emergence, Diversification, and Transmission of Subgroup J Avian Leukosis Virus Reveals That the Live Chicken Trade Plays a Critical Role in the Adaption and Endemicity of Viruses to the Yellow-Chickens. J. Virol. 2022, 96, e00717-22. [Google Scholar] [CrossRef]
- Gilka, F.; Spencer, J.L. Chronic Myocarditis and Circulatory Syndrome in a White Leghorn Strain Induced by an Avian Leukosis Virus: Light and Electron Microscopic Study. Avian Dis. 1990, 34, 174. [Google Scholar] [CrossRef]
- Chesters, P.M.; Smith, L.P.; Nair, V. E (XSR) Element Contributes to the Oncogenicity of Avian Leukosis Virus (Subgroup J). J. Gen. Virol. 2006, 87, 2685–2692. [Google Scholar] [CrossRef] [PubMed]
- Hayward, W.S.; Neel, B.G.; Astrin, S.M. Activation of a Cellular Onc Gene by Promoter Insertion in ALV-Induced Lymphoid Leukosis. Nature 1981, 290, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.I.; Johnson, J.A.; Da Silva Freire, M.; Heneine, W. Identification and Characterization of Avian Retroviruses in Chicken Embryo-Derived Yellow Fever Vaccines: Investigation of Transmission to Vaccine Recipients. J. Virol. 2003, 77, 1105–1111. [Google Scholar] [CrossRef] [Green Version]
- Shahabuddin, M.; Sears, J.F.; Khan, A.S. No Evidence of Infectious Retroviruses in Measles Virus Vaccines Produced in Chicken Embryo Cell Cultures. J. Clin. Microbiol. 2001, 39, 675–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, A.I.; Shanmugam, V.; Switzer, W.M.; Tsang, S.X.; Fadly, A.; Thea, D.; Helfand, R.; Bellini, W.J.; Folks, T.M.; Heneine, W. Lack of Evidence of Endogenous Avian Leukosis Virus and Endogenous Avian Retrovirus Transmission to Measles, Mumps, and Rubella Vaccine Recipients. Emerg. Infect. Dis. 2001, 7, 66–72. [Google Scholar] [CrossRef] [Green Version]
- Schat, K.A.; Erb, H.N. Lack of Evidence That Avian Oncogenic Viruses Are Infectious for Humans: A Review. Avian Dis. 2014, 58, 345–358. [Google Scholar] [CrossRef]
- Silva, R.F.; Fadly, A.M.; Taylor, S.P. Development of a Polymerase Chain Reaction to Differentiate Avian Leukosis Virus (ALV) Subgroups: Detection of an ALV Contaminant in Commercial Marek’s Disease Vaccines. Avian Dis. 2007, 51, 663–667. [Google Scholar] [CrossRef]
- Wang, P.; Li, M.; Li, H.; Bi, Y.; Lin, L.; Shi, M.; Huang, T.; Mo, M.; Wei, T.; Wei, P. ALV-J-Contaminated Commercial Live Vaccines Induced Pathogenicity in Three-Yellow Chickens: One of the Transmission Routes of ALV-J to Commercial Chickens. Poult. Sci. 2021, 100, 101027. [Google Scholar] [CrossRef]
- Mao, Y.; Su, Q.; Li, J.; Jiang, T.; Wang, Y. Avian Leukosis Virus Contamination in Live Vaccines: A Retrospective Investigation in China. Vet. Microbiol. 2020, 246, 108712. [Google Scholar] [CrossRef]
- Tang, S.; Li, J.; Chang, Y.-F.; Lin, W. Avian Leucosis Virus-Host Interaction: The Involvement of Host Factors in Viral Replication. Front. Immunol. 2022, 13, 907287. [Google Scholar] [CrossRef]
- Rubin, H.; Fanshier, L.; Cornelius, A.; Hughes, W.F. Tolerance and Immunity in Chickens after Congenital and Contact Infection with an Avian Leukosis Virus. Virology 1962, 17, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cui, S.; Li, W.; Wang, Y.; Cui, Z.; Zhao, P.; Chang, S. Vertical Transmission of Avian Leukosis Virus Subgroup J (ALV-J) from Hens Infected through Artificial Insemination with ALV-J Infected Semen. BMC Vet. Res. 2017, 13, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacon, L.D.; Smith, E.; Crittenden, L.B.; Havenstein, G.B. Association of the Slow Feathering (K) and an Endogenous Viral (Ev 21) Gene on the Z Chromosome of Chickens. Poult. Sci. 1988, 67, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Huda, A.; Polavarapu, N.; Jordan, I.K.; McDonald, J.F. Endogenous Retroviruses of the Chicken Genome. Biol. Direct. 2008, 3, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, A.S.; Miedzinska, K.; Kebede, A.; Bamidele, O.; Al-Jumaili, A.S.; Dessie, T.; Hanotte, O.; Smith, J. Diversity of Endogenous Avian Leukosis Virus Subgroup E (ALVE) Insertions in Indigenous Chickens. Genet. Sel. Evol. 2020, 52, 29. [Google Scholar] [CrossRef]
- Federspiel, M.J. Reverse Engineering Provides Insights on the Evolution of Subgroups A to E Avian Sarcoma and Leukosis Virus Receptor Specificity. Viruses 2019, 11, 497. [Google Scholar] [CrossRef] [Green Version]
- Fenton, S.P.; Reddy, M.R.; Bagust, T.J. Single and Concurrent Avian Leukosis Virus Infections with Avian Leukosis Virus-J and Avian Leukosis Virus-A in Australian Meat-Type Chickens. Avian Pathol. 2005, 34, 48–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Q.; Li, M.; He, C.; Lu, Q.; Gao, Y.; Li, Q.; Shi, M.; Wang, P.; Wei, P. Genetic Diversity of Avian Leukosis Virus Subgroup J (ALV-J): Toward a Unified Phylogenetic Classification and Nomenclature System. Virus Evol. 2021, 7, veab037. [Google Scholar] [CrossRef]
- Chang, S.-W.; Hsu, M.-F.; Wang, C.-H. Gene Detection, Virus Isolation, and Sequence Analysis of Avian Leukosis Viruses in Taiwan Country Chickens. Avian Dis. 2013, 57, 172–177. [Google Scholar] [CrossRef]
- Su, Q.; Li, Y.; Li, W.; Cui, S.; Tian, S.; Cui, Z.; Zhao, P.; Chang, S. Molecular Characteristics of Avian Leukosis Viruses Isolated from Indigenous Chicken Breeds in China. Poult. Sci. 2018, 97, 2917–2925. [Google Scholar] [CrossRef]
- Burmester, B.R.; Walter, W.G.; Gross, M.A.; Fontes, A.K. The Oncogenic Spectrum of Two Pure Strains of Avian Leukosis. J. Natl. Cancer Inst. 1959, 23, 277–291. [Google Scholar] [PubMed]
- Purchase, H.G.; Okazaki, W.; Vogt, P.K.; Hanafusa, H.; Burmester, B.R.; Crittenden, L.B. Oncogenicity of Avian Leukosis Viruses of Different Subgroups and of Mutants of Sarcoma Viruses. Infect. Immun. 1977, 15, 423–428. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, S.; Ochiai, K.; Abe, A.; Kishi, S.; Takayama, K.; Sunden, Y. Astrocytic Growth through the Autocrine/Paracrine Production of IL-1β in the Early Infectious Phase of Fowl Glioma-Inducing Virus. Avian Pathol. 2014, 43, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Yu, M.; Chang, F.; Xing, L.; Bao, Y.; Wang, S.; Farooque, M.; Li, X.; Liu, P.; Chen, Y.; et al. Molecular Characterization of Avian Leukosis Virus Subgroup J in Chinese Local Chickens between 2013 and 2018. Poult. Sci. 2020, 99, 5286–5296. [Google Scholar] [CrossRef] [PubMed]
- Payne, L.N.; Brown, S.R.; Bumstead, N.; Howes, K.; Frazier, J.A.; Thouless, M.E. A Novel Subgroup of Exogenous Avian Leukosis Virus in Chickens. J. Gen. Virol. 1991, 72 Pt 4, 801–807. [Google Scholar] [CrossRef]
- Xu, B.; Dong, W.; Yu, C.; He, Z.; Lv, Y.; Sun, Y.; Feng, X.; Li, N.; Lee, L.F.; Li, M. Occurrence of Avian Leukosis Virus Subgroup J in Commercial Layer Flocks in China. Avian Pathol. 2004, 33, 13–17. [Google Scholar] [CrossRef]
- Lai, H.; Zhang, H.; Ning, Z.; Chen, R.; Zhang, W.; Qing, A.; Xin, C.; Yu, K.; Cao, W.; Liao, M. Isolation and Characterization of Emerging Subgroup J Avian Leukosis Virus Associated with Hemangioma in Egg-Type Chickens. Vet. Microbiol. 2011, 151, 275–283. [Google Scholar] [CrossRef]
- Bai, J.; Payne, L.N.; Skinner, M.A. HPRS-103 (Exogenous Avian Leukosis Virus, Subgroup J) Has an Env Gene Related to Those of Endogenous Elements EAV-0 and E51 and an E Element Found Previously Only in Sarcoma Viruses. J. Virol. 1995, 69, 779–784. [Google Scholar] [CrossRef] [Green Version]
- Benson, S.J.; Ruis, B.L.; Garbers, A.L.; Fadly, A.M.; Conklin, K.F. Independent Isolates of the Emerging Subgroup J Avian Leukosis Virus Derive from a Common Ancestor. J. Virol. 1998, 72, 10301–10304. [Google Scholar] [CrossRef]
- Yao, Y.; Smith, L.P.; Nair, V.; Watson, M. An Avian Retrovirus Uses Canonical Expression and Processing Mechanisms to Generate Viral MicroRNA. J. Virol. 2014, 88, 2–9. [Google Scholar] [CrossRef] [Green Version]
- Meng, F.; Li, X.; Fang, J.; Gao, Y.; Zhu, L.; Xing, G.; Tian, F.; Gao, Y.; Dong, X.; Chang, S.; et al. Genomic Diversity of the Avian Leukosis Virus Subgroup J Gp85 Gene in Different Organs of an Infected Chicken. J. Vet. Sci. 2016, 17, 497–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borodin, A.M.; Emanuilova, Z.V.; Smolov, S.V.; Ogneva, O.A.; Konovalova, N.V.; Terentyeva, E.V.; Serova, N.Y.; Efimov, D.N.; Fisinin, V.I.; Greenberg, A.J.; et al. Eradication of Avian Leukosis Virus Subgroups J and K in Broiler Cross Chickens by Selection against Infected Birds Using Multilocus PCR. PLoS ONE 2022, 17, e0269525. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Lin, L.; Li, H.; Yang, Y.; Huang, T.; Wei, P. Diversity and Evolution Analysis of Glycoprotein GP85 from Avian Leukosis Virus Subgroup J Isolates from Chickens of Different Genetic Backgrounds during 1989–2016: Coexistence of Five Extremely Different Clusters. Arch. Virol. 2018, 163, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, P.; Cui, Z.-Z. Identification of a new subgroup of avian leukosis virus isolated from Chinese indigenous chicken breeds. Chin. J. Virol. 2012, 28, 609–614. [Google Scholar]
- Li, X.; Lin, W.; Chang, S.; Zhao, P.; Zhang, X.; Liu, Y.; Chen, W.; Li, B.; Shu, D.; Zhang, H.; et al. Isolation, Identification and Evolution Analysis of a Novel Subgroup of Avian Leukosis Virus Isolated from a Local Chinese Yellow Broiler in South China. Arch. Virol. 2016, 161, 2717–2725. [Google Scholar] [CrossRef]
- Chen, J.; Li, J.; Dong, X.; Liao, M.; Cao, W. The Key Amino Acid Sites 199-205, 269, 319, 321 and 324 of ALV-K Env Contribute to the Weaker Replication Capacity of ALV-K than ALV-A. Retrovirology 2022, 19, 19. [Google Scholar] [CrossRef]
- Liang, X.; Gu, Y.; Chen, X.; Li, T.; Gao, Y.; Wang, X.; Fang, C.; Fang, S.; Yang, Y. Identification and Characterization of a Novel Natural Recombinant Avian Leucosis Virus from Chinese Indigenous Chicken Flock. Virus Genes 2019, 55, 726–733. [Google Scholar] [CrossRef]
- Mason, A.S.; Fulton, J.E.; Hocking, P.M.; Burt, D.W. A New Look at the LTR Retrotransposon Content of the Chicken Genome. BMC Genom. 2016, 17, 688. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Zhu, W.; Chen, S.; Liu, Y.; Sun, Z.; Geng, T.; Song, C.; Gao, B.; Wang, X.; Qin, A.; et al. Expression Patterns of Endogenous Avian Retrovirus ALVE1 and Its Response to Infection with Exogenous Avian Tumour Viruses. Arch. Virol. 2017, 162, 89–101. [Google Scholar] [CrossRef]
- Mason, A.S.; Lund, A.R.; Hocking, P.M.; Fulton, J.E.; Burt, D.W. Identification and Characterisation of Endogenous Avian Leukosis Virus Subgroup E (ALVE) Insertions in Chicken Whole Genome Sequencing Data. Mob. DNA 2020, 11, 22. [Google Scholar] [CrossRef]
- Smith, E.J.; Fadly, A.M.; Levin, I.; Crittenden, L.B. The Influence of Ev 6 on the Immune Response to Avian Leukosis Virus Infection in Rapid-Feathering Progeny of Slow- and Rapid-Feathering Dams. Poult. Sci. 1991, 70, 1673–1678. [Google Scholar] [CrossRef] [PubMed]
- Elferink, M.G.; Vallée, A.A.A.; Jungerius, A.P.; Crooijmans, R.P.M.A.; Groenen, M.A.M. Partial Duplication of the PRLR and SPEF2 Genes at the Late Feathering Locus in Chicken. BMC Genom. 2008, 9, 391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu, G.; Huang, G.; Fu, H.; Li, J.; Huang, S.; Wang, Y. Characterization of the Novel Duplicated PRLR Gene at the Late-Feathering K Locus in Lohmann Chickens. J. Mol. Endocrinol. 2013, 51, 261–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, D.L.; Garwood, V.A.; Lowe, P.C.; Hester, P.Y.; Crittenden, L.B.; Fadly, A.M. Influence of Sex-Linked Feathering Phenotypes of Parents and Progeny Upon Lymphoid Leukosis Virus Infection Status and Egg Production. Poult. Sci. 1984, 63, 401–413. [Google Scholar] [CrossRef]
- Mays, J.K.; Black-Pyrkosz, A.; Mansour, T.; Schutte, B.C.; Chang, S.; Dong, K.; Hunt, H.D.; Fadly, A.M.; Zhang, L.; Zhang, H. Endogenous Avian Leukosis Virus in Combination with Serotype 2 Marek’s Disease Virus Significantly Boosted the Incidence of Lymphoid Leukosis-Like Bursal Lymphomas in Susceptible Chickens. J. Virol. 2019, 93, e00861-19. [Google Scholar] [CrossRef] [Green Version]
- Cao, W.; Mays, J.; Kulkarni, G.; Dunn, J.; Fulton, R.M.; Fadly, A. Further Observations on Serotype 2 Marek’s Disease Virus-Induced Enhancement of Spontaneous Avian Leukosis Virus-like Bursal Lymphomas in ALVA6 Transgenic Chickens. Avian Pathol. 2015, 44, 23–27. [Google Scholar] [CrossRef]
- Ramoutar, V.V.; Johnson, Y.J.; Kohrt, L.J.; Bahr, J.M.; Iwai, A.; Caporali, E.H.G.; Myint, M.S.; Szigetvari, N.; Stewart, M.C. Retroviral Association with Ovarian Adenocarcinoma in Laying Hens. Avian Pathol. 2022, 51, 113–119. [Google Scholar] [CrossRef]
- Fulton, J.E.; Mason, A.S.; Wolc, A.; Arango, J.; Settar, P.; Lund, A.R.; Burt, D.W. The Impact of Endogenous Avian Leukosis Viruses (ALVE) on Production Traits in Elite Layer Lines. Poult. Sci. 2021, 100, 101121. [Google Scholar] [CrossRef]
- Gavora, J.S.; Kuhnlein, U.; Crittenden, L.B.; Spencer, J.L.; Sabour, M.P. Endogenous Viral Genes: Association with Reduced Egg Production Rate and Egg Size in White Leghorns. Poult. Sci. 1991, 70, 618–623. [Google Scholar] [CrossRef]
- Kuhnlein, U.; Sabour, M.; Gavora, J.S.; Fairfull, R.W.; Bernon, D.E. Influence of Selection for Egg Production and Marek’s Disease Resistance on the Incidence of Endogenous Viral Genes in White Leghorns. Poult. Sci. 1989, 68, 1161–1167. [Google Scholar] [CrossRef]
- Fox, W.; Smyth, J.R. The Effects of Recessive White and Dominant White Genotypes on Early Growth Rate. Poult. Sci. 1985, 64, 429–433. [Google Scholar] [CrossRef] [PubMed]
- Ka, S.; Kerje, S.; Bornold, L.; Liljegren, U.; Siegel, P.B.; Andersson, L.; Hallböök, F. Proviral Integrations and Expression of Endogenous Avian Leucosis Virus during Long Term Selection for High and Low Body Weight in Two Chicken Lines. Retrovirology 2009, 6, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aswad, A.; Katzourakis, A. Paleovirology and Virally Derived Immunity. Trends Ecol. Evol. 2012, 27, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Chiu, E.S.; VandeWoude, S. Endogenous Retroviruses Drive Resistance and Promotion of Exogenous Retroviral Homologs. Annu. Rev. Anim. Biosci. 2021, 9, 225–248. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Hu, X.; Cui, I.H.; Wu, S.; Dou, C.; Liu, Y.; Sun, Z.; Xue, S.; Geng, T.; Liu, Z.; et al. An Endogenous Retroviral Element Exerts an Antiviral Innate Immune Function via the Derived LncRNA Lnc-ALVE1-AS1. Antivir. Res. 2019, 170, 104571. [Google Scholar] [CrossRef]
- Benkel, B.F. Locus-Specific Diagnostic Tests for Endogenous Avian Leukosis-Type Viral Loci in Chickens. Poult. Sci. 1998, 77, 1027–1035. [Google Scholar] [CrossRef]
- Rutherford, K.; Meehan, C.J.; Langille, M.G.I.; Tyack, S.G.; McKay, J.C.; McLean, N.L.; Benkel, K.; Beiko, R.G.; Benkel, B. Discovery of an Expanded Set of Avian Leukosis Subgroup E Proviruses in Chickens Using Vermillion, a Novel Sequence Capture and Analysis Pipeline. Poult. Sci. 2016, 95, 2250–2258. [Google Scholar] [CrossRef]
- Mason, A.S. The Abundance and Diversity of Endogenous Retroviruses in the Chicken Genome. Ph.D. Thesis, University of Edinburgh, Edinburgh, UK, 2018. [Google Scholar]
- Shao, H.; Wang, L.; Sang, J.; Li, T.; Liu, Y.; Wan, Z.; Qian, K.; Qin, A.; Ye, J. Novel Avian Leukosis Viruses from Domestic Chicken Breeds in Mainland China. Arch. Virol. 2017, 162, 2073–2076. [Google Scholar] [CrossRef]
- Wang, P.; Niu, J.; Xue, C.; Han, Z.; Abdelazez, A.; Xinglin, Z. Two Novel Recombinant Avian Leukosis Virus Isolates from Luxi Gamecock Chickens. Arch. Virol. 2020, 165, 2877–2881. [Google Scholar] [CrossRef]
- Cai, L.; Shen, Y.; Wang, G.; Guo, H.; Liu, J.; Cheng, Z. Identification of Two Novel Multiple Recombinant Avian Leukosis Viruses in Two Different Lines of Layer Chicken. J. Gen. Virol. 2013, 94, 2278–2286. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Liu, L.; Niu, X.; Li, J.; Kang, Z.; Han, C.; Gao, Y.; Qi, X.; Wang, Y.; Wang, X.; et al. Research Note: A Novel Recombinant Subgroup E Isolate of the Avian Leukosis Virus with a Subgroup B-like Gp85 Region in China. Poult. Sci. 2021, 100, 101137. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, T.; Zavala, G.; Cheng, S. Molecular Characterization of Three Recombinant Isolates of Avian Leukosis Virus Obtained from Contaminated Marek’s Disease Vaccines. Avian Dis. 2008, 52, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Shi, M.; He, C.; Lin, L.; Li, H.; Gu, Z.; Li, M.; Gao, Y.; Huang, T.; Mo, M.; et al. A Novel Recombinant Avian Leukosis Virus Isolated from Gamecocks Induced Pathogenicity in Three-Yellow Chickens: A Potential Infection Source of Avian Leukosis Virus to the Commercial Chickens. Poult. Sci. 2019, 98, 6497–6504. [Google Scholar] [CrossRef] [PubMed]
- M. Landman, W.J.; Nieuwenhuisen-van Wilgen, J.L.; Koch, G.; Dwars, R.M.; Ultee, A.; Gruys, E. Avian Leukosis Virus Subtype J in Ovo-Infected Specific Pathogen Free Broilers Harbour the Virus in Their Feathers and Show Feather Abnormalities. Avian Pathol. 2001, 30, 675–684. [Google Scholar] [CrossRef]
- Murphy, B. Retroviridae. In Fenner’s Veterinary Virology; Elsevier: Amsterdam, The Netherlands, 2017; pp. 269–297. ISBN 978-0-12-800946-8. [Google Scholar]
- Ronfort, C.; Afanassieff, M.; Chebloune, Y.; Dambrine, G.; Nigon, V.M.; Verdier, G. Identification and Structure Analysis of Endogenous Proviral Sequences in a Brown Leghorn Chicken Strain. Poult. Sci. 1991, 70, 2161–2175. [Google Scholar] [CrossRef]
- Mothes, W.; Boerger, A.L.; Narayan, S.; Cunningham, J.M.; Young, J.A.T. Retroviral Entry Mediated by Receptor Priming and Low PH Triggering of an Envelope Glycoprotein. Cell 2000, 103, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Svoboda, J. Cell Association in Rous Sarcoma Virus (RSV) Rescue and Cell Infection. Folia Biol. 2015, 61, 161–167. [Google Scholar]
- Brown, D.W.; Blais, B.P.; Robinson, H.L. Long Terminal Repeat (LTR) Sequences, Env, and a Region near the 5’ LTR Influence the Pathogenic Potential of Recombinants between Rous-Associated Virus Types 0 and 1. J. Virol. 1988, 62, 3431–3437. [Google Scholar] [CrossRef] [Green Version]
- Mingzhang, R.; Zijun, Z.; Lixia, Y.; Jian, C.; Min, F.; Jie, Z.; Ming, L.; Weisheng, C. The Construction and Application of a Cell Line Resistant to Novel Subgroup Avian Leukosis Virus (ALV-K) Infection. Arch. Virol. 2018, 163, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Rao, M.; Liao, M.; Cao, W. Phylogenetic Analysis and Pathogenicity Assessment of the Emerging Recombinant Subgroup K of Avian Leukosis Virus in South China. Viruses 2018, 10, 194. [Google Scholar] [CrossRef] [Green Version]
- Swanstrom, R.; Graham, W.D.; Zhou, S. Sequencing the Biology of Entry: The Retroviral Env Gene. Curr. Top Microbiol. Immunol. 2017, 407, 65–82. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, H.; Fang, X.; Gao, K.; Fang, C.; Gu, Y.; Gao, Y.; Wang, X.; Huang, H.; Liang, X.; et al. Identification of a Novel Epitope Specific for Gp85 Protein of Avian Leukosis Virus Subgroup K. Vet. Immunol. Immunopathol. 2020, 230, 110143. [Google Scholar] [CrossRef] [PubMed]
- Böhnlein, S.; Hauber, J.; Cullen, B.R. Identification of a U5-Specific Sequence Required for Efficient Polyadenylation within the Human Immunodeficiency Virus Long Terminal Repeat. J. Virol. 1989, 63, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Ruddell, A. Transcription Regulatory Elements of the Avian Retroviral Long Terminal Repeat. Virology 1995, 206, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Deffaud, C.; Darlix, J.-L. Rous Sarcoma Virus Translation Revisited: Characterization of an Internal Ribosome Entry Segment in the 5′ Leader of the Genomic RNA. J. Virol. 2000, 74, 11581–11588. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Lucia, E.; Ocaña, J.; Benitez, L.; Fandiño, S.; Domenech, A. In Silico Analysis Reveals the Similarity of Transcription Binding Site Motifs in Endogenous and Exogenous Gammaretroviruses from Different Animal Species. Biochem. Genet. 2023; under review. [Google Scholar]
- Gómez-Lucía, E.; Collado, V.M.; Miró, G.; Doménech, A. Effect of Type-I Interferon on Retroviruses. Viruses 2009, 1, 545–573. [Google Scholar] [CrossRef] [Green Version]
- Fandiño, S.; Gomez-Lucia, E.; Lumbreras, J.; Benitez, L.; Domenech, A. Analysis of Transcription Binding Sites Present in the Long Terminal Repeats of Different Avian Leukosis Viruses in Various Bird Species. manuscript in preparation.
- Platanias, L.C. Mechanisms of Type-I- and Type-II-Interferon-Mediated Signalling. Nat. Rev. Immunol. 2005, 5, 375–386. [Google Scholar] [CrossRef]
- Uddin, S.; Lekmine, F.; Sassano, A.; Rui, H.; Fish, E.N.; Platanias, L.C. Role of Stat5 in Type I Interferon-Signaling and Transcriptional Regulation. Biochem. Biophys. Res. Commun. 2003, 308, 325–330. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-Like Receptor Signaling Pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [Green Version]
- De Bosscher, K.; Vanden Berghe, W.; Vermeulen, L.; Plaisance, S.; Boone, E.; Haegeman, G. Glucocorticoids Repress NF-ΚB-Driven Genes by Disturbing the Interaction of P65 with the Basal Transcription Machinery, Irrespective of Coactivator Levels in the Cell. Proc. Natl. Acad. Sci. USA 2000, 97, 3919–3924. [Google Scholar] [CrossRef]
- Chikhirzhina, G.I.; Al-Shekhadat, R.I.; Chikhirzhina, E.V. Transcription Factors of the NF1 Family: Role in Chromatin Remodeling. Mol. Biol. 2008, 42, 342–356. [Google Scholar] [CrossRef]
- Ma, X.; Montaner, L.J. Proinflammatory Response and IL-12 Expression in HIV-1 Infection. J. Leukoc. Biol. 2000, 68, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, S.; Winans, S.; Lam, G.; Justice, J.; Morgan, R.; Beemon, K. Selection for Avian Leukosis Virus Integration Sites Determines the Clonal Progression of B-Cell Lymphomas. PLoS Pathog. 2017, 13, e1006708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clurman, B.E.; Hayward, W.S. Multiple Proto-Oncogene Activations in Avian Leukosis Virus- Induced Lymphomas: Evidence for Stage-Specific Events. Mol. Cell Biol. 1989, 9, 2657–2664. [Google Scholar] [CrossRef]
- Tam, W.; Dahlberg, J.E. MiR-155/BIC as an Oncogenic MicroRNA. Genes Chromosom. Cancer 2006, 45, 211–212. [Google Scholar] [CrossRef]
- Miklík, D.; Šenigl, F.; Hejnar, J. Proviruses with Long-Term Stable Expression Accumulate in Transcriptionally Active Chromatin Close to the Gene Regulatory Elements: Comparison of ASLV-, HIV- and MLV-Derived Vectors. Viruses 2018, 10, 116. [Google Scholar] [CrossRef] [Green Version]
- Justice, J.; Malhotra, S.; Ruano, M.; Li, Y.; Zavala, G.; Lee, N.; Morgan, R.; Beemon, K. The MET Gene Is a Common Integration Target in Avian Leukosis Virus Subgroup J-Induced Chicken Hemangiomas. J. Virol. 2015, 89, 4712–4719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bande, F.; Arshad, S.S.; Omar, A.R. Isolation and Metagenomic Identification of Avian Leukosis Virus Associated with Mortality in Broiler Chicken. Adv. Virol. 2016, 2016, 9058403. [Google Scholar] [CrossRef] [Green Version]
- Brojatsch, J.; Naughton, J.; Adkins, H.B.; Young, J.A.T. TVB Receptors for Cytopathic and Noncytopathic Subgroups of Avian Leukosis Viruses Are Functional Death Receptors. J. Virol. 2000, 74, 11490–11494. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Zeng, X.; Hua, Y.; Gao, Q.; Fan, Z.; Chai, H.; Wang, Q.; Qi, X.; Wang, Y.; Gao, H.; et al. Genetic Diversity and Phylogenetic Analysis of Glycoprotein Gp85 of Avian Leukosis Virus Subgroup J Wild-Bird Isolates from Northeast China. Arch. Virol. 2014, 159, 1821–1826. [Google Scholar] [CrossRef]
- Meng, F.; Li, Q.; Zhang, Y.; Zhang, Z.; Tian, S.; Cui, Z.; Chang, S.; Zhao, P. Characterization of Subgroup J Avian Leukosis Virus Isolated from Chinese Indigenous Chickens. Virol. J. 2018, 15, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yehia, N.; El-Sayed, H.S.; Omar, S.E.; Amer, F. Genetic Variability of the Avian Leukosis Virus Subgroup J Gp85 Gene in Layer Flocks in Lower Egypt. Vet. World 2020, 13, 1065–1072. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Sun, S.; Zhang, Z.; Meng, S. Simultaneous Endemic Infections with Subgroup J Avian Leukosis Virus and Reticuloendotheliosis Virus in Commercial and Local Breeds of Chickens. Avian Pathol. 2009, 38, 443–448. [Google Scholar] [CrossRef]
- Payne, L.N. Developments in Avian Leukosis Research. Leukemia 1992, 6 (Suppl. S3), 150S–152S. [Google Scholar] [PubMed]
- Dong, X.; Zhao, P.; Xu, B.; Fan, J.; Meng, F.; Sun, P.; Ju, S.; Li, Y.; Chang, S.; Shi, W.; et al. Avian Leukosis Virus in Indigenous Chicken Breeds, China. Emerg. Microbes Infect. 2015, 4, e76. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Li, T.; Hu, M.; Deng, J.; Liu, Y.; Xie, Q.; Shao, H.; Ye, J.; Qin, A. A Recombination Efficiently Increases the Pathogenesis of the Novel K Subgroup of Avian Leukosis Virus. Vet. Microbiol. 2019, 231, 214–217. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Li, Y.; Cui, Z.; Chang, S.; Zhao, P. The Emerging Novel Avian Leukosis Virus with Mutations in the Pol Gene Shows Competitive Replication Advantages Both in Vivo and in Vitro. Emerg. Microbes Infect. 2018, 7, 117. [Google Scholar] [CrossRef]
- Ayala, A.J.; Yabsley, M.J.; Hernandez, S.M. A Review of Pathogen Transmission at the Backyard Chicken–Wild Bird Interface. Front. Vet. Sci. 2020, 7, 539925. [Google Scholar] [CrossRef]
- Pohjola, L.; Rossow, L.; Huovilainen, A.; Soveri, T.; Hänninen, M.-L.; Fredriksson-Ahomaa, M. Questionnaire Study and Postmortem Findings in Backyard Chicken Flocks in Finland. Acta Vet. Scand. 2015, 57, 3. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.P.; Petheridge, L.; Nair, V.; Wood, A.; Welchman, D. Avian Leukosis Virus Subgroup J-Associated Myelocytoma in a Hobby Chicken. Vet. Rec. 2018, 182, 23. [Google Scholar] [CrossRef]
- Williams, S.M.; Barbosa, T.; Hafner, S.; Zavala, G. Myxosarcomas Associated with Avian Leukosis Virus Subgroup A Infection in Fancy Breed Chickens. Avian Dis. 2010, 54, 1319–1322. [Google Scholar] [CrossRef]
- Mete, A.; Giannitti, F.; Barr, B.; Woods, L.; Anderson, M. Causes of Mortality in Backyard Chickens in Northern California: 2007–2011. Avian Dis. 2013, 57, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Cadmus, K.J.; Mete, A.; Harris, M.; Anderson, D.; Davison, S.; Sato, Y.; Helm, J.; Boger, L.; Odani, J.; Ficken, M.D.; et al. Causes of Mortality in Backyard Poultry in Eight States in the United States. J. VET Diagn. Investg. 2019, 31, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Wunderwald, C.; Hoop, R.K. Serological Monitoring of 40 Swiss Fancy Breed Poultry Flocks. Avian Pathol. 2002, 31, 157–162. [Google Scholar] [CrossRef] [Green Version]
- de Wit, J.J.; van Eck, J.H.; Crooijmans, R.P.; Pijpers, A. A Serological Survey for Pathogens in Old Fancy Chicken Breeds in Central and Eastern Part of The Netherlands. Tijdschr. Diergeneeskd 2004, 129, 324–327. [Google Scholar]
- Payne, L.N.; Howes, K.; Gillespie, A.M.; Smith, L.M. Host Range of Rous Sarcoma Virus Pseudotype RSV(HPRS-103) in 12 Avian Species: Support for a New Avian Retrovirus Envelope Subgroup, Designated J. J. Gen. Virol. 1992, 73, 2995–2997. [Google Scholar] [CrossRef] [PubMed]
- Melder, D.C.; Pankratz, V.S.; Federspiel, M.J. Evolutionary Pressure of a Receptor Competitor Selects Different Subgroup a Avian Leukosis Virus Escape Variants with Altered Receptor Interactions. J. Virol. 2003, 77, 10504–10514. [Google Scholar] [CrossRef] [Green Version]
- Plachý, J.; Reinišová, M.; Kučerová, D.; Šenigl, F.; Stepanets, V.; Hron, T.; Trejbalová, K.; Elleder, D.; Hejnar, J. Identification of New World Quails Susceptible to Infection with Avian Leukosis Virus Subgroup J. J. Virol. 2017, 91, e02002-16. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Hu, W.; Li, B.; Chen, R.; Shen, W.; Guo, H.; Guo, H.; Li, H. Comparison of Viremia, Cloacal Virus Shedding, Antibody Responses and Pathological Lesions in Adult Chickens, Quails, and Pigeons Infected with ALV-A. Sci. Rep. 2019, 9, 3027. [Google Scholar] [CrossRef] [Green Version]
- Venugopal, K.; Howes, K.; Flannery, D.M.J.; Payne, L.N. Subgroup J Avian Leukosis Virus Infection in Turkeys: Induction of Rapid Onset Tumours by Acutely Transforming Virus Strain 966. Avian Pathol. 2000, 29, 319–325. [Google Scholar] [CrossRef]
- Zeghdoudi, M.; Aoun, L.; Merdaci, L.; Bouzidi, N. Epidemiological Features and Pathological Study of Avian Leukosis in Turkeys’ Flocks. Vet. World 2017, 10, 1135–1138. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Qin, L.; Gao, H.; Yang, B.; Liu, W.; Qi, X.; Wang, Y.; Zeng, X.; Liu, S.; Wang, X.; et al. Avian Leukosis Virus Subgroup A and B Infection in Wild Birds of Northeast China. Vet. Microbiol. 2013, 163, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; He, M.; Zhang, J.; Zhao, M.; Wang, G.; Cheng, Z. Cross-species Transmission of Avian Leukosis Virus Subgroup J. Chin. J. Virol. 2016, 32, 46–55. [Google Scholar]
- Shen, Y.; Cai, L.; Wang, Y.; Wei, R.; He, M.; Wang, S.; Wang, G.; Cheng, Z. Genetic Mutations of Avian Leukosis Virus Subgroup J Strains Extended Their Host Range. J. Gen. Virol. 2014, 95, 691–699. [Google Scholar] [CrossRef]
- Li, J.; Chen, J.; Dong, X.; Liang, C.; Guo, Y.; Chen, X.; Huang, M.; Liao, M.; Cao, W. Residues 140–142, 199–200, 222–223, and 262 in the Surface Glycoprotein of Subgroup A Avian Leukosis Virus Are the Key Sites Determining Tva Receptor Binding Affinity and Infectivity. Front. Microbiol. 2022, 13, 868377. [Google Scholar] [CrossRef]
- Han, C.; Hao, R.; Liu, L.; Zeng, X. Molecular Characterization of 3’UTRs of J Subgroup Avian Leukosis Virus in Passerine Birds in China. Arch. Virol. 2015, 160, 845–849. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Liu, L.; Hao, R.; Han, C. Detection and Molecular Characterization of J Subgroup Avian Leukosis Virus in Wild Ducks in China. PLoS ONE 2014, 9, e94980. [Google Scholar] [CrossRef]
- Reinišová, M.; Plachý, J.; Kučerová, D.; Šenigl, F.; Vinkler, M.; Hejnar, J. Genetic Diversity of NHE1, Receptor for Subgroup J Avian Leukosis Virus, in Domestic Chicken and Wild Anseriform Species. PLoS ONE 2016, 11, e0150589. [Google Scholar] [CrossRef]
- Khordadmehr, M.; Firouzamandi, M.; Zehtab-Najafi, M.; Shahbazi, R. Naturally Occurring Co-Infection of Avian Leukosis Virus (Subgroups A–E) and Reticuloendotheliosis Virus in Green Peafowls (Pavo muticus). Rev. Bras. Cienc. Avic. 2017, 19, 609–614. [Google Scholar] [CrossRef] [Green Version]
- García-Fernández, R.A.; Pérez-Martínez, C.; Espinosa-Alvarez, J.; Escudero-Diez, A.; García-Marín, J.F.; Núñez, A.; García-Iglesias, M.J. Lymphoid Leukosis in an Ostrich (Struthio camelus). Vet. Rec. 2000, 146, 676–677. [Google Scholar] [CrossRef]
- Frisby, D.P.; Weiss, R.A.; Roussel, M.; Stehelin, D. The Distribution of Endogenous Chicken Retrovirus Sequences in the DNA of Galliform Birds Does Not Coincide with Avian Phylogenetic Relationships. Cell 1979, 17, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Fujita, D.J.; Tal, J.; Varmus, H.E.; Bishop, J.M. Env Gene of Chicken RNA Tumor Viruses: Extent of Conservation in Cellular and Viral Genomes. J. Virol. 1978, 27, 465–474. [Google Scholar] [CrossRef] [Green Version]
- Hanafusa, T.; Hanafusa, H. Isolation of Leukosis-Type Virus from Pheasant Embryo Cells: Possible Presence of Viral Genes in Cells. Virology 1973, 51, 247–251. [Google Scholar] [CrossRef]
- Fujita, D.J.; Chen, Y.C.; Friis, R.R.; Vogt, P.K. RNA Tumor Viruses of Pheasants: Characterization of Avian Leukosis Subgroups F and G. Virology 1974, 60, 558–571. [Google Scholar] [CrossRef] [PubMed]
- Payne, L.N. Biology of Avian Retroviruses. In The Retroviridae; Levy, J.A., Ed.; Plenum Press: New York, NY, USA, 1992; Volume 1, pp. 299–404. [Google Scholar]
- Dimcheff, D.E.; Drovetski, S.V.; Krishnan, M.; Mindell, D.P. Cospeciation and Horizontal Transmission of Avian Sarcoma and Leukosis Virus Gag Genes in Galliform Birds. J. Virol. 2000, 74, 3984–3995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimcheff, D.E.; Krishnan, M.; Mindell, D.P. Evolution and Characterization of Tetraonine Endogenous Retrovirus: A New Virus Related to Avian Sarcoma and Leukosis Viruses. J. Virol. 2001, 75, 2002–2009. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.; Jie, H.; Lian, L.; Qu, L.J.; Hou, Z.C.; Zheng, J.X.; Chen, S.Y.; Yang, N.; Liu, Y.P. Avian Sarcoma and Leukosis Virus Gag Gene in the Anser Anser Domesticus Genome. Genet. Mol. Res. 2015, 14, 14379–14386. [Google Scholar] [CrossRef]
- Elleder, D.; Hejnar, J. Letter to the Editor: Avian sarcoma and leukosis virus gag gene—Genet. Mol. Res. 14 (4): 14379-14386 “Avian sarcoma and leukosis virus gag gene in the Anser anser domesticus genome”. Genet. Mol. Res. 2016, 15, 15014956. [Google Scholar] [CrossRef]
- Hao, R.; Han, C.; Liu, L.; Zeng, X. First Finding of Subgroup-E Avian Leukosis Virus from Wild Ducks in China. Vet. Microbiol. 2014, 173, 366–370. [Google Scholar] [CrossRef]
- Payne, L.N. Epizootiology of Avian Leukosis Virus Infections. In Avian Leukosis; Martinus Nijhoff Publishing: Boston, MA, USA, 1987; pp. 47–75. [Google Scholar]
- Bacon, L.D.; Hunt, H.D.; Cheng, H.H. A Review of the Development of Chicken Lines to Resolve Genes Determining Resistance to Diseases. Poult. Sci. 2000, 79, 1082–1093. [Google Scholar] [CrossRef]
- Himly, M.; Foster, D.N.; Bottoli, I.; Iacovoni, J.S.; Vogt, P.K. The DF-1 Chicken Fibroblast Cell Line: Transformation Induced by Diverse Oncogenes and Cell Death Resulting from Infection by Avian Leukosis Viruses. Virology 1998, 248, 295–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnard, R.J.O.; Elleder, D.; Young, J.A.T. Avian Sarcoma and Leukosis Virus-Receptor Interactions: From Classical Genetics to Novel Insights into Virus-Cell Membrane Fusion. Virology 2006, 344, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmen, S.L.; Federspiel, M.J. Selection of a Subgroup A Avian Leukosis Virus [ALV(A)] Envelope Resistant to Soluble ALV(A) Surface Glycoprotein. Virology 2000, 273, 364–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Přikryl, D.; Plachý, J.; Kučerová, D.; Koslová, A.; Reinišová, M.; Šenigl, F.; Hejnar, J. The Novel Avian Leukosis Virus Subgroup K Shares Its Cellular Receptor with Subgroup A. J. Virol. 2019, 93, e00580-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmen, S.L.; Melder, D.C.; Federspiel, M.J. Identification of Key Residues in Subgroup A Avian Leukosis Virus Envelope Determining Receptor Binding Affinity and Infectivity of Cells Expressing Chicken or Quail Tva Receptor. J. Virol. 2001, 75, 726–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.J.; Lee, K.Y.; Park, Y.H.; Choi, H.J.; Yao, Y.; Nair, V.; Han, J.Y. Acquisition of Resistance to Avian Leukosis Virus Subgroup B through Mutations on Tvb Cysteine-Rich Domains in DF-1 Chicken Fibroblasts. Vet. Res. 2017, 48, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munguia, A.; Federspiel, M.J. Efficient Subgroup C Avian Sarcoma and Leukosis Virus Receptor Activity Requires the IgV Domain of the Tvc Receptor and Proper Display on the Cell Membrane. J. Virol. 2008, 82, 11419–11428. [Google Scholar] [CrossRef] [Green Version]
- Chai, N.; Bates, P. Na+/H+ Exchanger Type 1 Is a Receptor for Pathogenic Subgroup J Avian Leukosis Virus. Proc. Natl. Acad. Sci. USA 2006, 103, 5531–5536. [Google Scholar] [CrossRef]
- Putney, L.K.; Denker, S.P.; Barber, D.L. The Changing Face of the Na + /H + Exchanger, NHE1: Structure, Regulation, and Cellular Actions. Annu. Rev. Pharmacol. Toxicol. 2002, 42, 527–552. [Google Scholar] [CrossRef]
- Kučerová, D.; Plachý, J.; Reinišová, M.; Šenigl, F.; Trejbalová, K.; Geryk, J.; Hejnar, J. Nonconserved Tryptophan 38 of the Cell Surface Receptor for Subgroup J Avian Leukosis Virus Discriminates Sensitive from Resistant Avian Species. J. Virol. 2013, 87, 8399–8407. [Google Scholar] [CrossRef] [Green Version]
- Mei, M.; Ye, J.; Qin, A.; Wang, L.; Hu, X.; Qian, K.; Shao, H. Identification of Novel Viral Receptors with Cell Line Expressing Viral Receptor-Binding Protein. Sci. Rep. 2015, 5, 7935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rup, B.J.; Hoelzer, J.D.; Bose, H.R. Helper Viruses Associated with Avian Acute Leukemia Viruses Inhibit the Cellular Immune Response. Virology 1982, 116, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Dai, M.; Xie, T.; Li, Z.; Shi, M.; Zhang, X. Innate Immune Responses in ALV-J Infected Chicks and Chickens with Hemangioma In Vivo. Front. Microbiol. 2016, 7, 786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, M.; Zhang, X. Immunity to Avian Leukosis Virus: Where Are We Now and What Should We Do? Front. Immunol. 2016, 7, 624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Li, W.; Zheng, S.J. Advances on Innate Immune Evasion by Avian Immunosuppressive Viruses. Front. Immunol. 2022, 13, 901913. [Google Scholar] [CrossRef]
- Haugh, K.A.; Shalginskikh, N.; Nogusa, S.; Skalka, A.M.; Katz, R.A.; Balachandran, S. The Interferon-Inducible Antiviral Protein Daxx Is Not Essential for Interferon-Mediated Protection against Avian Sarcoma Virus. Virol. J. 2014, 11, 100. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Ma, X.; Cui, X.; Zhou, J.; Li, C.; Huang, L.; Shang, Y.; Cheng, Z. Inhibition of Avian Tumor Virus Replication by CCCH-Type Zinc Finger Antiviral Protein. Oncotarget 2017, 8, 58865–58871. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Feng, W.; Cheng, Z.; Yang, J.; Bi, J.; Wang, X.; Wang, G. TRIM62-Mediated Restriction of Avian Leukosis Virus Subgroup J Replication Is Dependent on the SPRY Domain. Poult. Sci. 2019, 98, 6019–6025. [Google Scholar] [CrossRef]
- Zhou, J.-R.; Liu, J.-H.; Li, H.-M.; Zhao, Y.; Cheng, Z.; Hou, Y.-M.; Guo, H.-J. Regulatory Effects of Chicken TRIM25 on the Replication of ALV-A and the MDA5-Mediated Type I Interferon Response. Vet. Res. 2020, 51, 145. [Google Scholar] [CrossRef]
- Xie, T.; Feng, M.; Dai, M.; Mo, G.; Ruan, Z.; Wang, G.; Shi, M.; Zhang, X. Cholesterol-25-Hydroxylase Is a Chicken ISG That Restricts ALV-J Infection by Producing 25-Hydroxycholesterol. Viruses 2019, 11, 498. [Google Scholar] [CrossRef] [Green Version]
- Krchlíková, V.; Fábryová, H.; Hron, T.; Young, J.M.; Koslová, A.; Hejnar, J.; Strebel, K.; Elleder, D. Antiviral Activity and Adaptive Evolution of Avian Tetherins. J. Virol. 2020, 94, e00416-20. [Google Scholar] [CrossRef]
- Li, Z.; Luo, Q.; Xu, H.; Zheng, M.; Abdalla, B.A.; Feng, M.; Cai, B.; Zhang, X.; Nie, Q.; Zhang, X. MiR-34b-5p Suppresses Melanoma Differentiation-Associated Gene 5 (MDA5) Signaling Pathway to Promote Avian Leukosis Virus Subgroup J (ALV-J)-Infected Cells Proliferaction and ALV-J Replication. Front. Cell. Infect. Microbiol. 2017, 7, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Chen, B.; Feng, M.; Ouyang, H.; Zheng, M.; Ye, Q.; Nie, Q.; Zhang, X. MicroRNA-23b Promotes Avian Leukosis Virus Subgroup J (ALV-J) Replication by Targeting IRF1. Sci. Rep. 2015, 5, 10294. [Google Scholar] [CrossRef] [Green Version]
- Ren, C.; Xie, R.; Yao, Y.; Yu, M.; Chang, F.; Xing, L.; Zhang, Y.; Liu, Y.; Wang, S.; Farooque, M.; et al. MiR-125b Suppression Inhibits Apoptosis and Negatively Regulates Sema4D in Avian Leukosis Virus-Transformed Cells. Viruses 2019, 11, 728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, M.; Feng, M.; Xie, T.; Li, Y.; Ruan, Z.; Shi, M.; Liao, M.; Zhang, X. ALV-J Infection Induces Chicken Monocyte Death Accompanied with the Production of IL-1β and IL-18. Oncotarget 2017, 8, 99889–99900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazzolo, L.; Moscovici, C.; Moscovici, M.G. Persistence of Avian Oncoviruses in Chicken Macrophages. Infect. Immun. 1979, 23, 294–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, M.; Dai, M.; Cao, W.; Tan, Y.; Li, Z.; Shi, M.; Zhang, X. ALV-J Strain SCAU-HN06 Induces Innate Immune Responses in Chicken Primary Monocyte-Derived Macrophages. Poult. Sci. 2017, 96, 42–50. [Google Scholar] [CrossRef]
- Liu, D.; Dai, M.; Zhang, X.; Cao, W.; Liao, M. Subgroup J Avian Leukosis Virus Infection of Chicken Dendritic Cells Induces Apoptosis via the Aberrant Expression of MicroRNAs. Sci. Rep. 2016, 6, 20188. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Qiu, Q.; Zhang, X.; Dai, M.; Qin, J.; Hao, J.; Liao, M.; Cao, W. Infection of Chicken Bone Marrow Mononuclear Cells with Subgroup J Avian Leukosis Virus Inhibits Dendritic Cell Differentiation and Alters Cytokine Expression. Infect. Genet. Evol. 2016, 44, 130–136. [Google Scholar] [CrossRef]
- Mo, G.; Fu, H.; Hu, B.; Zhang, Q.; Xian, M.; Zhang, Z.; Lin, L.; Shi, M.; Nie, Q.; Zhang, X. SOCS3 Promotes ALV-J Virus Replication via Inhibiting JAK2/STAT3 Phosphorylation During Infection. Front. Cell Infect. Microbiol. 2021, 11, 748795. [Google Scholar] [CrossRef]
- Lin, W.; Xu, Z.; Yan, Y.; Zhang, H.; Li, H.; Chen, W.; Chen, F.; Xie, Q. Avian Leukosis Virus Subgroup J Attenuates Type I Interferon Production Through Blocking IκB Phosphorylation. Front. Microbiol. 2018, 9, 1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Wang, D.; Liu, Y.; Zhao, R.; Wu, T.; Hu, X.; Pan, Z.; Cui, H. Targeting the Histone Methyltransferase Disruptor of Telomeric Silencing 1-Like Restricts Avian Leukosis Virus Subgroup J Replication by Restoring the Innate Immune Response in Chicken Macrophages. Front. Microbiol. 2020, 11, 603131. [Google Scholar] [CrossRef]
- Fadly, A.M. Avian Leukosis Virus (ALV) Infection, Shedding, and Tumors in Maternal ALV Antibody-Positive and -Negative Chickens Exposed to Virus at Hatching. Avian Dis. 1988, 32, 89. [Google Scholar] [CrossRef] [PubMed]
- Maas, H.J.L.; De Boer, G.F.; Groenendal, J.E. Age Related Resistance to Avian Leukosis Virus. III. Infectious Virus, Neutralising Antibody and Tumours in Chickens Inoculated at Various Ages. Avian Pathol. 1982, 11, 309–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadly, A.M.; Smith, E.J. Isolation and Some Characteristics of a Subgroup J-like Avian Leukosis Virus Associated with Myeloid Leukosis in Meat-Type Chickens in the United States. Avian Dis. 1999, 43, 391–400. [Google Scholar] [CrossRef]
- Pandiri, A.R.; Mays, J.K.; Silva, R.F.; Hunt, H.D.; Reed, W.M.; Fadly, A.M. Subgroup J Avian Leukosis Virus Neutralizing Antibody Escape Variants Contribute to Viral Persistence in Meat-Type Chickens. Avian Dis. 2010, 54, 848–856. [Google Scholar] [CrossRef]
- Qualtiere, L.F.; Meyers, P. A Reexamination of Humoral Tolerance in Chickens Congenitally Infected with an Avian Leukosis Virus. J. Immunol. 1979, 122, 825–829. [Google Scholar] [CrossRef]
- Meyers, P. Antibody Response to Related Leukosis Viruses Induced in Chickens Tolerant to an Avian Leukosis Virus. J. Natl. Cancer Inst. 1976, 56, 381–386. [Google Scholar] [CrossRef]
- He, S.; Zheng, G.; Zhou, D.; Huang, L.; Dong, J.; Cheng, Z. High-Frequency and Activation of CD4+CD25+ T Cells Maintain Persistent Immunotolerance Induced by Congenital ALV-J Infection. Vet. Res. 2021, 52, 119. [Google Scholar] [CrossRef]
- He, S.; Zheng, G.; Zhou, D.; Li, G.; Zhu, M.; Du, X.; Zhou, J.; Cheng, Z. Clonal Anergy of CD117+chB6+ B Cell Progenitors Induced by Avian Leukosis Virus Subgroup J Is Associated with Immunological Tolerance. Retrovirology 2019, 16, 1. [Google Scholar] [CrossRef]
- Wang, Q.; Li, X.; Ji, X.; Wang, J.; Shen, N.; Gao, Y.; Qi, X.; Wang, Y.; Gao, H.; Zhang, S.; et al. A Recombinant Avian Leukosis Virus Subgroup J for Directly Monitoring Viral Infection and the Selection of Neutralizing Antibodies. PLoS ONE 2014, 9, e115422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandiri, A.R.; Reed, W.M.; Mays, J.K.; Fadly, A.M. Influence of Strain, Dose of Virus, and Age at Inoculation on Subgroup J Avian Leukosis Virus Persistence, Antibody Response, and Oncogenicity in Commercial Meat-Type Chickens. Avian Dis. 2007, 51, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Thacker, E.L.; Fulton, J.E.; Hunt, H.D. In Vitro Analysis of a Primary, Major Histocompatibility Complex (MHC)-Restricted, Cytotoxic T-Lymphocyte Response to Avian Leukosis Virus (ALV), Using Target Cells Expressing MHC Class I CDNA Inserted into a Recombinant ALV Vector. J. Virol. 1995, 69, 6439–6444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadly, A.M.; Lee, L.F.; Bacon, L.D. Immunocompetence of Chickens during Early and Tumorigenic Stages of Rous-Associated Virus-1 Infection. Infect. Immun. 1982, 37, 1156–1161. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Li, S.; Shi, K.; Liao, J.; Sun, H.; Liao, M. Systematic Identification of Host Immune Key Factors Influencing Viral Infection in PBL of ALV-J Infected SPF Chicken. Viruses 2020, 12, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Li, H.; Cheng, Z.; Liu, J.; Cui, Z. Influence of REV and ALV-J Co-Infection on Immunologic Function of T Lymphocytes and Histopathology in Broiler Chickens. Agric. Sci. China 2010, 9, 1667–1676. [Google Scholar] [CrossRef]
- Wang, F.; Wang, X.; Chen, H.; Liu, J.; Cheng, Z. The Critical Time of Avian Leukosis Virus Subgroup J-Mediated Immunosuppression during Early Stage Infection in Specific Pathogen-Free Chickens. J. Vet. Sci. 2011, 12, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, K.; Gao, Y.; Gao, L.; Zhong, L.; Zhang, Y.; Liu, C.; Zhang, Y.; Wang, X. Recombinant Marek’s Disease Virus as a Vector-Based Vaccine against Avian Leukosis Virus Subgroup J in Chicken. Viruses 2016, 8, 301. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Li, H.; Zhang, Z.; Sun, S.; Cheng, Z.; Liu, J.; Zhao, P.; Ren, Q.; Guo, H. Antibody Responses Induced by Recombinant ALV-A Gp85 Protein Vaccine Combining with CpG-ODN Adjuvant in Breeder Hens and the Protection for Their Offspring against Early Infection. Antivir. Res. 2015, 116, 20–26. [Google Scholar] [CrossRef]
- Liu, J.; Gao, K.; Li, D.; Zeng, Y.; Chen, X.; Liang, X.; Fang, C.; Gu, Y.; Wang, C.; Yang, Y. Recombinant Invasive Lactobacillus Plantarum Expressing the J Subgroup Avian Leukosis Virus Gp85 Protein Induces Protection against Avian Leukosis in Chickens. Appl. Microbiol. Biotechnol. 2022, 106, 729–742. [Google Scholar] [CrossRef]
- Meng, Q.-W.; Zhang, Z.-P.; Wang, W.; Tian, J.; Xiao, Z.-G. Enhanced Inhibition of Avian Leukosis Virus Subgroup J Replication by Multi-Target MiRNAs. Virol. J. 2011, 8, 556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, M.; Wu, S.; Feng, M.; Feng, S.; Sun, C.; Bai, D.; Gu, M.; Liao, M.; Cao, W. Recombinant Chicken Interferon-Alpha Inhibits the Replication of Exogenous Avian Leukosis Virus (ALV) in DF-1 Cells. Mol. Immunol. 2016, 76, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Su, Q.; Liu, B.; Li, Y.; Sun, W.; Liu, Y.; Xue, R.; Chang, S.; Wang, Y.; Zhao, P. Enhanced Antiviral Ability by a Combination of Zidovudine and Short Hairpin RNA Targeting Avian Leukosis Virus. Front. Microbiol. 2021, 12, 808982. [Google Scholar] [CrossRef] [PubMed]
- Witter, R.L.; Fadly, A.M. Reduction of Horizontal Transmission of Avian Leukosis Virus Subgroup J in Broiler Breeder Chickens Hatched and Reared in Small Groups. Avian Pathol. 2001, 30, 641–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adkins, H.B.; Brojatsch, J.; Naughton, J.; Rolls, M.M.; Pesola, J.M.; Young, J.A.T. Identification of a Cellular Receptor for Subgroup E Avian Leukosis Virus. Proc. Natl. Acad. Sci. USA 1997, 94, 11617–11622. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.T.; Chen, S.Y.; Chen, W.G.; Liu, Y.; Sun, B.L.; Li, H.X.; Zhang, H.M.; Qu, H.; Wang, J.; Shu, D.M.; et al. Single Nucleotide Polymorphism Variants within Tva and Tvb Receptor Genes in Chinese Chickens. Poult. Sci. 2014, 93, 2482–2489. [Google Scholar] [CrossRef]
- Guan, X.; Zhang, Y.; Yu, M.; Ren, C.; Gao, Y.; Yun, B.; Liu, Y.; Wang, Y.; Qi, X.; Liu, C.; et al. Residues 28 to 39 of the Extracellular Loop 1 of Chicken Na+/H+ Exchanger Type I Mediate Cell Binding and Entry of Subgroup J Avian Leukosis Virus. J. Virol. 2018, 92, e01627-17. [Google Scholar] [CrossRef] [Green Version]
- Koslová, A.; Trefil, P.; Mucksová, J.; Krchlíková, V.; Plachý, J.; Krijt, J.; Reinišová, M.; Kučerová, D.; Geryk, J.; Kalina, J.; et al. Knock-Out of Retrovirus Receptor Gene Tva in the Chicken Confers Resistance to Avian Leukosis Virus Subgroups A and K and Affects Cobalamin (Vitamin B12)-Dependent Level of Methylmalonic Acid. Viruses 2021, 13, 2504. [Google Scholar] [CrossRef]
- Koslová, A.; Kučerová, D.; Reinišová, M.; Geryk, J.; Trefil, P.; Hejnar, J. Genetic Resistance to Avian Leukosis Viruses Induced by CRISPR/Cas9 Editing of Specific Receptor Genes in Chicken Cells. Viruses 2018, 10, 605. [Google Scholar] [CrossRef] [Green Version]
- Wallny, H.-J.; Avila, D.; Hunt, L.G.; Powell, T.J.; Riegert, P.; Salomonsen, J.; Skjødt, K.; Vainio, O.; Vilbois, F.; Wiles, M.V.; et al. Peptide Motifs of the Single Dominantly Expressed Class I Molecule Explain the Striking MHC-Determined Response to Rous Sarcoma Virus in Chickens. Proc. Natl. Acad. Sci. USA 2006, 103, 1434–1439. [Google Scholar] [CrossRef]
- Zhang, Q.; Xie, T.; Mo, G.; Zhang, Z.; Lin, L.; Zhang, X. ACSL1 Inhibits ALV-J Replication by IFN-Ⅰ Signaling and PI3K/Akt Pathway. Front. Immunol. 2021, 12, 774323. [Google Scholar] [CrossRef]
- Mountford, J.; Gheyas, A.; Vervelde, L.; Smith, J. Genetic Variation in Chicken Interferon Signalling Pathway Genes in Research Lines Showing Differential Viral Resistance. Anim. Genet. 2022, 53, 640–656. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yan, Y.; Lin, W.; Li, A.; Zhang, H.; Lei, X.; Dai, Z.; Li, X.; Li, H.; Chen, W.; et al. Circular RNA Vav3 Sponges Gga-MiR-375 to Promote Epithelial-Mesenchymal Transition. RNA Biol. 2019, 16, 118–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Yan, Y.; Lei, X.; Li, A.; Zhang, H.; Dai, Z.; Li, X.; Chen, W.; Lin, W.; Chen, F.; et al. Circular RNA Alterations Are Involved in Resistance to Avian Leukosis Virus Subgroup-J-Induced Tumor Formation in Chickens. Oncotarget 2017, 8, 34961–34970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Pan, W.; Zhang, L.; Liu, J.; Ouyang, H.; Nie, Q.; Zhang, X. NHE1 Gene Associated with Avian Leukosis Virus Subgroup J Infection in Chicken. Mol. Biol. Rep. 2014, 41, 6519–6524. [Google Scholar] [CrossRef]
- Dai, M.; Feng, M.; Xie, T.; Li, Y.; Zhang, X. Fluctuations in Luteinizing Hormone, Follicle Stimulating Hormone, and Progesterone Might Affect the Disappearance of Avian Leukosis Virus Subgroup J Viremia in Chickens with Intermittent Viremia. Poult. Sci. 2019, 98, 3533–3538. [Google Scholar] [CrossRef]
- Mo, G.; Hu, B.; Wang, G.; Xie, T.; Fu, H.; Zhang, Q.; Fu, R.; Feng, M.; Luo, W.; Li, H.; et al. Prolactin Affects the Disappearance of ALV-J Viremia in Vivo and Inhibits Viral Infection. Vet. Microbiol. 2021, 261, 109205. [Google Scholar] [CrossRef]
- Mo, G.; Hu, B.; Zhang, Q.; Ruan, Z.; Li, W.; Liang, J.; Shen, Y.; Mo, Z.; Zhang, Z.; Wu, Z.; et al. DPRLR Causes Differences in Immune Responses between Early and Late Feathering Chickens after ALV-J Infection. Vet. Res. 2022, 53, 1. [Google Scholar] [CrossRef]
- Gomez-Lucia, E.; Tejerizo, G.; Domenech, A. Effect of Steroid Hormones on Retroviruses. In Oncogenic Viruses Research Trends; Johannes, L.T., Ed.; Nova Science: Hauppauge, NY, USA, 2007; pp. 163–191. ISBN 978-60021-628-2. [Google Scholar]
- Domenech, A.; Tejerizo, G.; Illera, J.C.; Gomez-Lucia, E. Effect of Male Steroid Hormones on Retroviral Infections. Curr. Top. Steroid Res. 2009, 6, 69–81. [Google Scholar]
- Greenig, M. HERVs, Immunity, and Autoimmunity: Understanding the Connection. PeerJ 2019, 7, e6711. [Google Scholar] [CrossRef] [Green Version]
- Collado, V.M.; Domenech, A.; Miró, G.; Martin, S.; Escolar, E.; Gomez-Lucia, E. Epidemiological Aspects and Clinicopathological Findings in Cats Naturally Infected with Feline Leukemia Virus (FeLV) and/or Feline Immunodeficiency Virus (FIV). Open J. Vet. Med. 2012, 2, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Erbeck, K.; Gagne, R.B.; Kraberger, S.; Chiu, E.S.; Roelke-Parker, M.; VandeWoude, S. Feline Leukemia Virus (FeLV) Endogenous and Exogenous Recombination Events Result in Multiple FeLV-B Subtypes during Natural Infection. J. Virol. 2021, 95, e0035321. [Google Scholar] [CrossRef] [PubMed]
- Denner, J. The Origin of Porcine Endogenous Retroviruses (PERVs). Arch. Virol. 2021, 166, 1007–1013. [Google Scholar] [CrossRef] [PubMed]
- Mason, A.S.; Fulton, J.E.; Smith, J. Endogenous Avian Leukosis Virus Subgroup E Elements of the Chicken Reference Genome. Genet. Mol. Biol. 2020, 99, 2911–2915. [Google Scholar] [CrossRef]
- Chen, W.; Qu, H.; Li, C.; Luo, C.; Wang, J.; Yang, C.; Shu, D. Polymorphism of Avian Leukosis Virus Subgroup E Loci Showing Selective Footprints in Chicken. Biochem. Genet. 2014, 52, 524–537. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, K.; Benkel, B.F. Characterization of Insertion Sites and Development of Locus-Specific Assays for Three Broiler-Derived Subgroup E Avian Leukosis Virus Proviruses. Avian Pathol. 2013, 42, 373–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Yuan, Y.; Zheng, G.; Sun, M.; Wang, Q.; Wu, J.; Li, J.; Sun, C.; Wang, Y.; Yang, N.; et al. Short Communication: Diversity of Endogenous Avian Leukosis Virus Subgroup E Elements in 11 Chicken Breeds. J. Anim. Sci. 2023, 101, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rutherford, K.; McLean, N.; Benkel, B.F. A Rapid Profiling Assay for Avian Leukosis Virus Subgroup e Proviruses in Chickens. Avian Dis. 2014, 58, 34–38. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alv Subgroup | En/ex 1 | Cytophaticity | Recombination 2 | Cell Receptor | Hosts |
|---|---|---|---|---|---|
| ALV-A | EX | NO | ALV-J | Tva protein | Galliformes (Gallus gallus; Meleagris gallopavo; Pavo muticus) Anseriformes (Sibirionetta formosa; Anas carolinensis) |
| ALV-B | EX | YES | ALV-J ALV-E | Tvb protein | Galliformes (Gallus gallus; Meleagris gallopavo) Anseriformes (Sibirionetta formosa; Anas carolinensis) |
| ALV-C | EX | NO | ALV-J | Tvc protein | Galliformes (Meleagris gallopavo) |
| ALV-D | EX | YES | Tvb protein | Galliformes (Meleagris gallopavo) | |
| ALV-E | EN | - | ALV-A ALV-B ALV-C ALV-J | Tvb protein | Galliformes (Gallus gallus; G. sonneratii (EAV-HP); Meleagris gallopavo; Lophortyx gambelii, Pavo muticus) Anseriformes (Anas falcata; A. carolinensis; Sibirionetta formosa) |
| ALV-F | EN | Galliformes (Phasianus colchicus; P. versicolor; Chrysolophus pictus) | |||
| ALV-G | EN | Galliformes (Chrysolophus pictus) | |||
| ALV-H | EX | Galliformes (Perdix perdix) | |||
| ALV-I | EX | Galliformes (Callipepla gambelii) | |||
| ALV-J | EX | ALV-A ALV-B ALV-C | chNHE1 | Galliformes (Gallus gallus; Meleagris gallopavo; Perdix perdix) Anseriformes (Anas acuta; A. poecilorhyncha; A. penelope; A. carolinensis; A. crecca; A. clypeata; A. formosa; Sibirionetta formosa; Mareca strepera) Passeriformes (Tarsiger cyanurus; Emberiza elegans; Phylloscopus inornatus; Poecile palustris) | |
| ALV-K | EX | ALV-E | Tva protein | Galliformes (Chinese indigenous chickens: Gallus gallus) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fandiño, S.; Gomez-Lucia, E.; Benítez, L.; Doménech, A. Avian Leukosis: Will We Be Able to Get Rid of It? Animals 2023, 13, 2358. https://doi.org/10.3390/ani13142358
Fandiño S, Gomez-Lucia E, Benítez L, Doménech A. Avian Leukosis: Will We Be Able to Get Rid of It? Animals. 2023; 13(14):2358. https://doi.org/10.3390/ani13142358
Chicago/Turabian StyleFandiño, Sergio, Esperanza Gomez-Lucia, Laura Benítez, and Ana Doménech. 2023. "Avian Leukosis: Will We Be Able to Get Rid of It?" Animals 13, no. 14: 2358. https://doi.org/10.3390/ani13142358