
Comparative Transcriptome Analysis Reveals the Effect of Aurantiochytrium sp. on Gonadal Development in Zebrafish

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Source and Diet Preparation
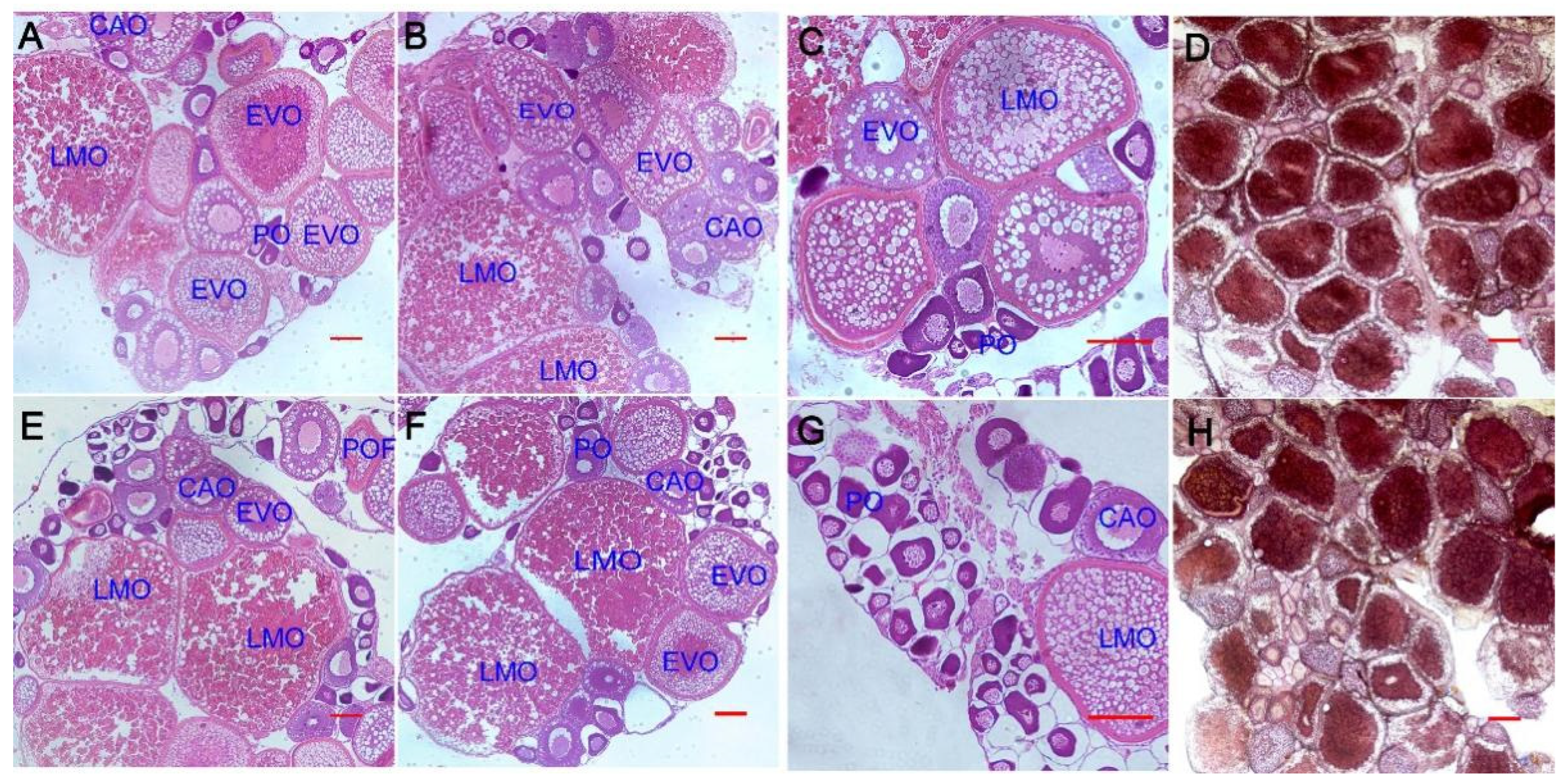
2.2. Histological Examination
2.3. The Influence of Varied Aurantiochytrium sp. Concentrations on In Vitro Oocyte Maturation
2.4. Intraperitoneal Administration to Assess the Impact of Aurantiochytrium sp. on Oocyte Maturation and Development
2.5. RNA-Seq Analysis and Bioinformatics
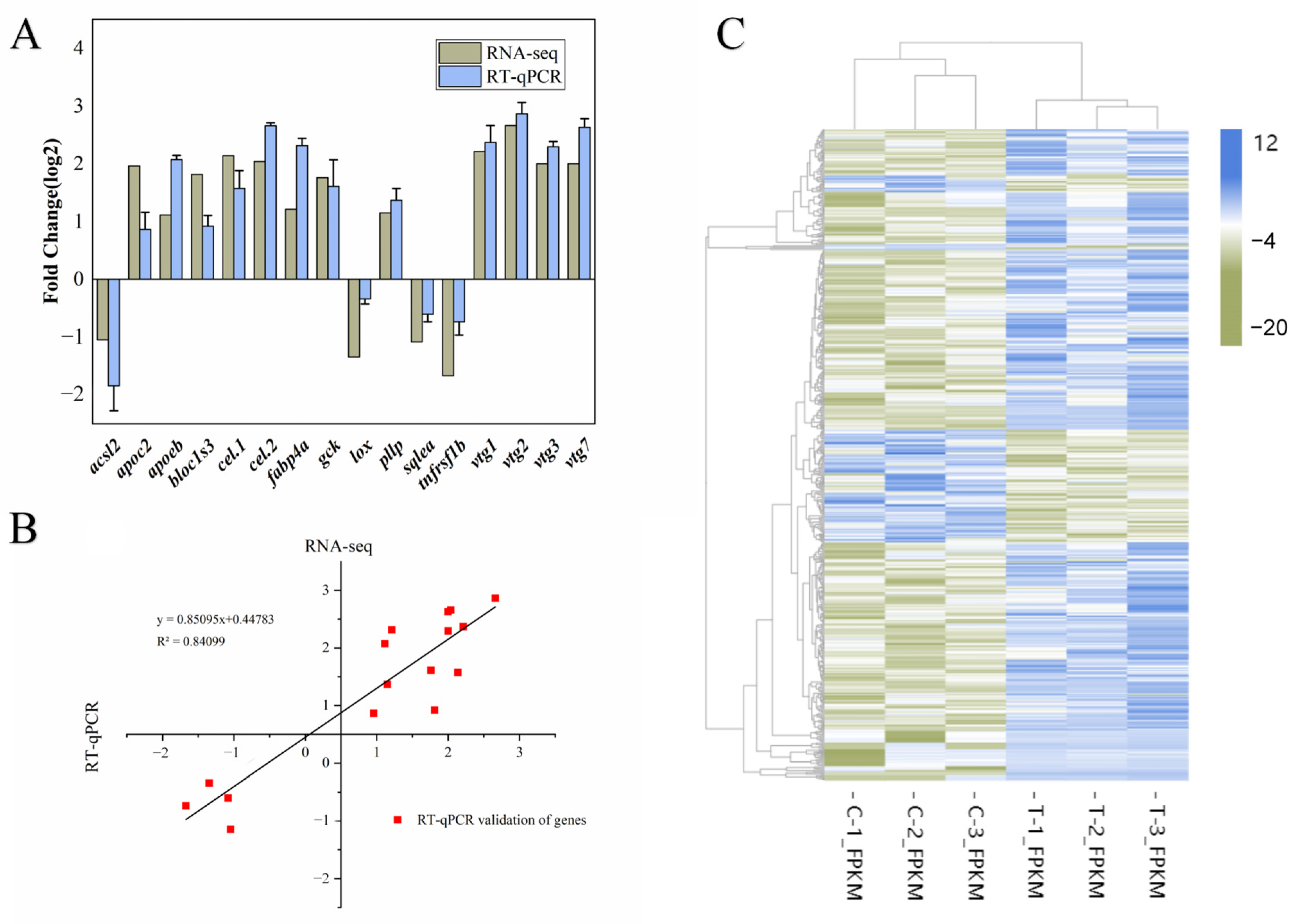
2.6. Real-Time Quantitative PCR (RT-qPCR) Validation
2.7. Analytical Statistics
3. Results
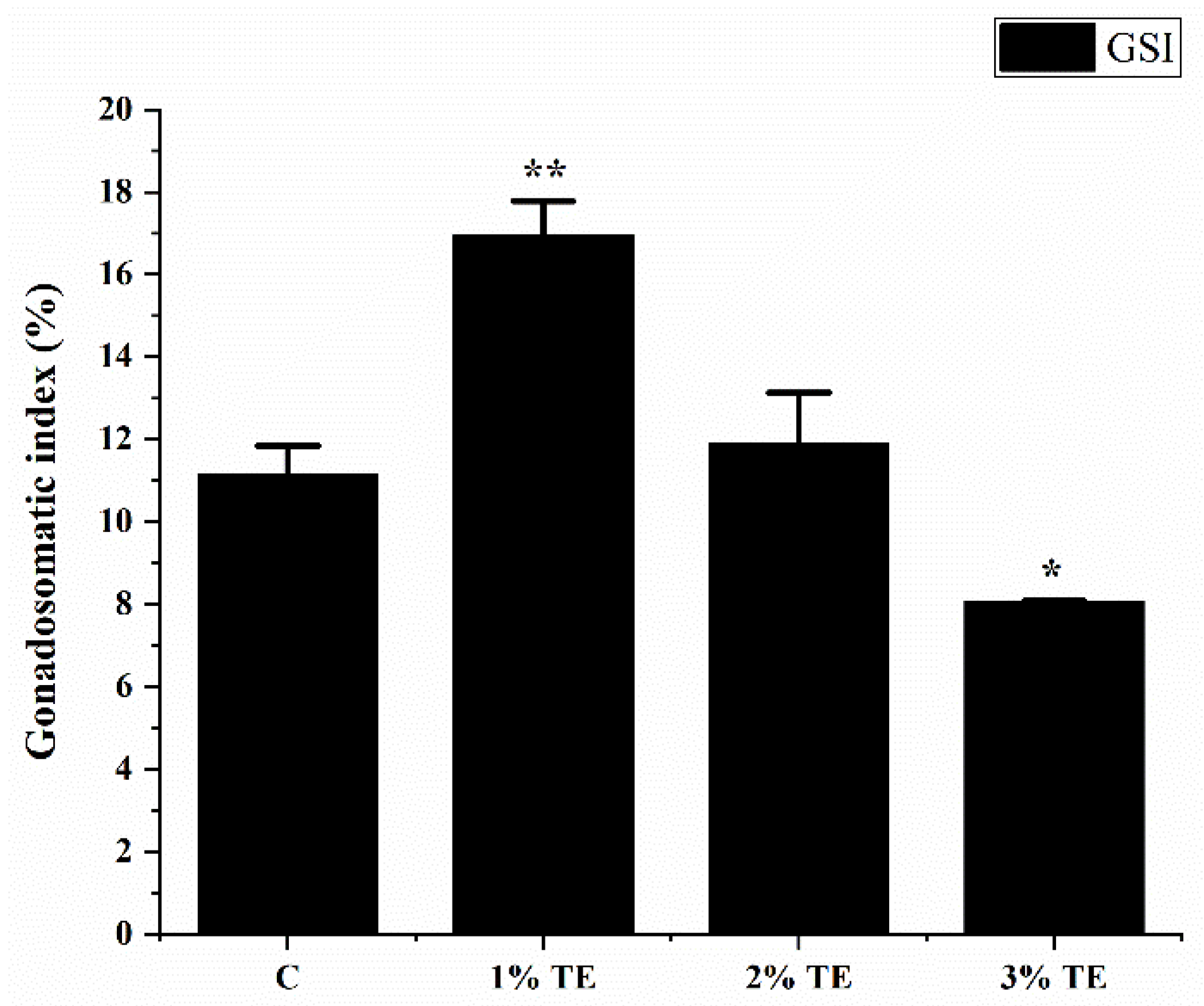
3.1. Effect of Aurantiochytrium sp. Extract on Gonadal Maturation in Zebrafish
3.2. Effects of Exposure to Different Concentrations of Active Products for 12 h on In Vitro Oocyte Maturation
3.3. Intraperitoneal Administration to Evaluate the Effect of Activity on Oocyte Maturation and Development
3.4. Transcriptome Analysis
3.5. Differential Expression Analysis
4. Discussion
4.1. Aurantiochytrium sp. Significantly Improves Gonadal Index in Zebrafish
4.2. Aurantiochytrium sp. Promotes Oocyte Maturation
4.3. Enriched GO and KEGG Terms
4.4. Transcriptome Reveals Lipid Metabolism Effects
4.5. Transcriptome Reveals Oocyte Development Effects
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-Year Retrospective Review of Global Aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Nasopoulou, C.; Zabetakis, I. Benefits of Fish Oil Replacement by Plant Originated Oils in Compounded Fish Feeds. A Review. LWT 2012, 47, 217–224. [Google Scholar] [CrossRef]
- Xu, Y.; Li, W.; Ding, Z. Polyunsaturated Fatty Acid Supplements Could Considerably Promote the Breeding Performance of Carp. Eur. J. Lipid Sci. Technol. 2017, 119, 1600183. [Google Scholar] [CrossRef]
- Lv, D.; Zhang, F.; Ding, J.; Chang, Y.; Zuo, R. Effects of Dietary N-3 LC-PUFA on the Growth Performance, Gonad Development, Fatty Acid Profile, Transcription of Related Genes and Intestinal Microflora in Adult Sea Urchin (Strongylocentrotus intermedius). Aquac. Res. 2021, 52, 1431–1441. [Google Scholar] [CrossRef]
- Olsen, R.L.; Hasan, M.R. A Limited Supply of Fishmeal: Impact on Future Increases in Global Aquaculture Production. Trends Food Sci. Technol. 2012, 27, 120–128. [Google Scholar] [CrossRef]
- Shepherd, C.J.; Monroig, O.; Tocher, D.R. Future Availability of Raw Materials for Salmon Feeds and Supply Chain Implications: The Case of Scottish Farmed Salmon. Aquaculture 2017, 467, 49–62. [Google Scholar] [CrossRef]
- Beal, C.M.; Gerber, L.N.; Thongrod, S.; Phromkunthong, W.; Kiron, V.; Granados, J.; Archibald, I.; Greene, C.H.; Huntley, M.E. Marine Microalgae Commercial Production Improves Sustainability of Global Fisheries and Aquaculture. Sci. Rep. 2018, 8, 15064. [Google Scholar] [CrossRef] [Green Version]
- Chi, G.; Xu, Y.; Cao, X.; Li, Z.; Cao, M.; Chisti, Y.; He, N. Production of Polyunsaturated Fatty Acids by Schizochytrium (Aurantiochytrium) Spp. Biotechnol. Adv. 2022, 55, 107897. [Google Scholar] [CrossRef]
- Iwasaka, H.; Koyanagi, R.; Satoh, R.; Nagano, A.; Watanabe, K.; Hisata, K.; Satoh, N.; Aki, T. A Possible Trifunctional β-Carotene Synthase Gene Identified in the Draft Genome of Aurantiochytrium Sp. Strain KH105. Genes 2018, 9, 200. [Google Scholar] [CrossRef] [Green Version]
- Aasen, I.M.; Ertesvåg, H.; Heggeset, T.M.B.; Liu, B.; Brautaset, T.; Vadstein, O.; Ellingsen, T.E. Thraustochytrids as Production Organisms for Docosahexaenoic Acid (DHA), Squalene, and Carotenoids. Appl. Microbiol. Biotechnol. 2016, 100, 4309–4321. [Google Scholar] [CrossRef]
- Jiang, Y.; Fan, K.-W.; Tsz-Yeung Wong, R.; Chen, F. Fatty Acid Composition and Squalene Content of the Marine Microalga Schizochytrium Mangrovei. J. Agric. Food Chem. 2004, 52, 1196–1200. [Google Scholar] [CrossRef]
- Raghukumar, S. Thraustochytrid Marine Protists: Production of PUFAs and Other Emerging Technologies. Mar. Biotechnol. 2008, 10, 631–640. [Google Scholar] [CrossRef]
- Tocher, D.R. Omega-3 Long-Chain Polyunsaturated Fatty Acids and Aquaculture in Perspective. Aquaculture 2015, 449, 94–107. [Google Scholar] [CrossRef]
- Russo, G.L.; Langellotti, A.L.; Oliviero, M.; Sacchi, R.; Masi, P. Sustainable Production of Food Grade Omega-3 Oil Using Aquatic Protists: Reliability and Future Horizons. New Biotechnol. 2021, 62, 32–39. [Google Scholar] [CrossRef]
- Willer, D.F.; Furse, S.; Aldridge, D.C. Microencapsulated Algal Feeds as a Sustainable Replacement Diet for Broodstock in Commercial Bivalve Aquaculture. Sci. Rep. 2020, 10, 12577. [Google Scholar] [CrossRef]
- Jaseera, K.V.; Ebeneezar, S.; Sayooj, P.; Nair, A.V.; Kaladharan, P. Dietary Supplementation of Microalgae, Aurantiochytrium Sp. and Co-Feeding with Artemia Enhances the Growth, Stress Tolerance and Survival in Penaeus Monodon (Fabricius, 1798) Post Larvae. Aquaculture 2021, 533, 736176. [Google Scholar] [CrossRef]
- Chang, Y.-H.; Lin, Y.-H.; Hsieh, C.; Chen, Y.-M. Aurantiochytrium Dietary Supplements Reduce Intra-Cohort Cannibalism among Orange-Spotted Groupers (Epinephelus coioides) by Modulating Brain 5-HT and Serum Cortisol. Aquaculture 2019, 502, 202–211. [Google Scholar] [CrossRef]
- Humaidah, N.; Nakai, S.; Nishijima, W.; Gotoh, T.; Furuta, M. Application of Aurantiochytrium Sp. L3W for Food-Processing Wastewater Treatment in Combination with Polyunsaturated Fatty Acids Production for Fish Aquaculture. Sci. Total Environ. 2020, 743, 140735. [Google Scholar] [CrossRef]
- Guimarães, A.M.; Dias Schleder, D.; Nagata, M.; Nóbrega, R.O.; Fracalossi, D.M.; Quadros Seiffert, W.; do Nascimento Vieira, F. Aurantiochytrium Sp. Meal Can Replace Fish Oil in Practical Diets for the Juvenile Pacific White Shrimp. Aquac. Nutr. 2019, 25, 798–807. [Google Scholar] [CrossRef]
- Batista, R.O.; Nobrega, R.O.; Schleder, D.D.; Pettigrew, J.E.; Fracalossi, D.M. Aurantiochytrium Sp. Meal Improved Body Fatty Acid Profile and Morphophysiology in Nile Tilapia Reared at Low Temperature. Fishes 2021, 6, 45. [Google Scholar] [CrossRef]
- Nobrega, R.O.; Batista, R.O.; Corrêa, C.F.; Mattioni, B.; Filer, K.; Pettigrew, J.E.; Fracalossi, D.M. Dietary Supplementation of Aurantiochytrium Sp. Meal, a Docosahexaenoic-Acid Source, Promotes Growth of Nile Tilapia at a Suboptimal Low Temperature. Aquaculture 2019, 507, 500–509. [Google Scholar] [CrossRef]
- Soudant, P.; Ventura, M.; Chauchat, L.; Guerreiro, M.; Mathieu-Resuge, M.; Le Grand, F.; Simon, V.; Collet, S.; Zambonino-Infante, J.-L.; Le Goïc, N. Evaluation of Aurantiochytrium Mangrovei Biomass Grown on Digestate as a Sustainable Feed Ingredient of Sea Bass, Dicentrarchus Labrax, Juveniles and Larvae. Sustainability 2022, 14, 14573. [Google Scholar] [CrossRef]
- Ribas, L.; Piferrer, F. The Zebrafish (Danio Rerio) as a Model Organism, with Emphasis on Applications for Finfish Aquaculture Research. Rev. Aquac. 2014, 6, 209–240. [Google Scholar] [CrossRef]
- Jørgensen, L.V.G. Zebrafish as a Model for Fish Diseases in Aquaculture. Pathogens 2020, 9, 609. [Google Scholar] [CrossRef]
- Lee-Estevez, M.; Figueroa, E.; Cosson, J.; Short, S.E.; Valdebenito, I.; Ulloa-Rodríguez, P.; Farias, J.G. Zebrafish as a Useful Model for Immunological Research with Potential Applications in Aquaculture. Rev. Aquac. 2018, 10, 213–223. [Google Scholar] [CrossRef]
- Ulloa, P.E.; Medrano, J.F.; Feijoo, C.G. Zebrafish as Animal Model for Aquaculture Nutrition Research. Front. Genet. 2014, 5, 313. [Google Scholar] [CrossRef] [Green Version]
- Aleström, P.; Winther-Larsen, H.C. Zebrafish Offer Aquaculture Research Their Services. Genomics Aquac. 2016, 165–194. [Google Scholar] [CrossRef]
- Yue, G.H.; Wang, L. Current Status of Genome Sequencing and Its Applications in Aquaculture. Aquaculture 2017, 468, 337–347. [Google Scholar] [CrossRef]
- Inbakandan, D. Transcriptomics in Aquaculture. Encycl. Mar. Biotechnol. 2020, 3, 1919–1936. [Google Scholar] [CrossRef]
- Chandhini, S.; Rejish Kumar, V.J. Transcriptomics in Aquaculture: Current Status and Applications. Rev. Aquac. 2019, 11, 1379–1397. [Google Scholar] [CrossRef]
- Liu, L.; Hu, Z.; Li, S.; Yang, H.; Li, S.; Lv, C.; Zaynab, M.; Cheng, C.H.K.; Chen, H.; Yang, X. Comparative Transcriptomic Analysis Uncovers Genes Responsible for the DHA Enhancement in the Mutant Aurantiochytrium Sp. Microorganisms 2020, 8, 529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Huang, Y.; Li, Z.; Guo, Y.; Li, S.; Huang, H.; Yang, X.; Li, G.; Chen, H. Effects of Dietary Supplementation with Aurantiochytrium Sp. on Zebrafish Growth as Determined by Transcriptomics. Animals 2022, 12, 2794. [Google Scholar] [CrossRef] [PubMed]
- Katti, P.A.; Goundadkar, B.B. Azole Pharmaceuticals Induce Germinal Vesicle Breakdown (GVBD) in Preovulatory Oocytes of Zebrafish (Danio Rerio): An in Vitro Study. Environ. Sci. Pollut. Res. 2021, 28, 3694–3702. [Google Scholar] [CrossRef] [PubMed]
- Norambuena, F.; Estévez, A.; Mañanós, E.; Bell, J.G.; Carazo, I.; Duncan, N. Effects of Graded Levels of Arachidonic Acid on the Reproductive Physiology of Senegalese Sole (Solea Senegalensis): Fatty Acid Composition, Prostaglandins and Steroid Levels in the Blood of Broodstock Bred in Captivity. Gen. Comp. Endocrinol. 2013, 191, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Henrotte, E.; Milla, S.; Mandiki, S.N.M.; Kestemont, P. Arachidonic Acid Induces Production of 17,20β-Dihydroxy-4-Pregnen-3-One (DHP) via a Putative PGE2 Receptor in Fish Follicles from the Eurasian Perch. Lipids 2011, 46, 179–187. [Google Scholar] [CrossRef]
- Xu, H.; Cao, L.; Zhang, Y.; Johnson, R.B.; Wei, Y.; Zheng, K.; Liang, M. Dietary Arachidonic Acid Differentially Regulates the Gonadal Steroidogenesis in the Marine Teleost, Tongue Sole (Cynoglossus Semilaevis), Depending on Fish Gender and Maturation Stage. Aquaculture 2017, 468, 378–385. [Google Scholar] [CrossRef]
- Flores, A.; Wiff, R.; Ganias, K.; Marshall, C.T. Accuracy of Gonadosomatic Index in Maturity Classification and Estimation of Maturity Ogive. Fish. Res. 2019, 210, 50–62. [Google Scholar] [CrossRef]
- Ge, W. Intrafollicular Paracrine Communication in the Zebrafish Ovary: The State of the Art of an Emerging Model for the Study of Vertebrate Folliculogenesis. Mol. Cell. Endocrinol. 2005, 237, 1–10. [Google Scholar] [CrossRef]
- Wang, Y.; Ge, W. Developmental Profiles of Activin ΒA, ΒB, and Follistatin Expression in the Zebrafish Ovary: Evidence for Their Differential Roles During Sexual Maturation and Ovulatory Cycle1. Biol. Reprod. 2004, 71, 2056–2064. [Google Scholar] [CrossRef]
- Zhao, C.; Zhai, Y.; Geng, R.; Wu, K.; Song, W.; Ai, N.; Ge, W. Genetic Analysis of Activin/Inhibin β Subunits in Zebrafish Development and Reproduction. PLoS Genet. 2022, 18, e1010523. [Google Scholar] [CrossRef]
- Harris, R.C.; Chung, E.; Coffey, R.J. EGF Receptor Ligands. In The EGF Receptor Family; Carpenter, G., Ed.; Academic Press: Burlington, NJ, USA, 2003; pp. 3–14. ISBN 978-0-12-160281-9. [Google Scholar]
- Song, Y.; Chen, W.; Zhu, B.; Ge, W. Disruption of Epidermal Growth Factor Receptor but Not EGF Blocks Follicle Activation in Zebrafish Ovary. Front. Cell Dev. Biol. 2022, 9, 750888. [Google Scholar] [CrossRef]
- Li, J.; Chu, L.; Sun, X.; Liu, Y.; Cheng, C.H.K. IGFs Mediate the Action of LH on Oocyte Maturation in Zebrafish. Mol. Endocrinol. 2015, 29, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Tang, H.; Wang, L.; He, J.; Guo, Y.; Liu, Y.; Liu, X.; Lin, H. Fertility Enhancement but Premature Ovarian Failure in Esr1-Deficient Female Zebrafish. Front. Endocrinol. 2018, 9, 567. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, O.; Patinote, A.; Nguyen, T.; Com, E.; Pineau, C.; Bobe, J. Genome Editing Reveals Reproductive and Developmental Dependencies on Specific Types of Vitellogenin in Zebrafish (Danio Rerio). Mol. Reprod. Dev. 2019, 86, 1168–1188. [Google Scholar] [CrossRef] [Green Version]
- Leerberg, D.M.; Sano, K.; Draper, B.W. Fibroblast growth factor signaling is required for early somatic gonad development in zebrafish. PLoS Genet. 2017, 13, e1006993. [Google Scholar] [CrossRef] [Green Version]
- Tse, A.C.-K.; Ge, W. Spatial Localization of EGF Family Ligands and Receptors in the Zebrafish Ovarian Follicle and Their Expression Profiles during Folliculogenesis. Gen. Comp. Endocrinol. 2010, 167, 397–407. [Google Scholar] [CrossRef]
- Mutz, K.-O.; Heilkenbrinker, A.; Lönne, M.; Walter, J.-G.; Stahl, F. Transcriptome Analysis Using Next-Generation Sequencing. Curr. Opin. Biotechnol. 2013, 24, 22–30. [Google Scholar] [CrossRef]
- Calder, P.C.; Grimble, R.F. Polyunsaturated Fatty Acids, Inflammation and Immunity. Eur. J. Clin. Nutr. 2002, 56, S14–S19. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R.B. Lipid Deposition in Oocytes of Teleost Fish During Secondary Oocyte Growth. Rev. Fish. Sci. 2009, 17, 78–89. [Google Scholar] [CrossRef]
- Thiel, W.A.; Esposito, E.J.; Findley, A.P.; Blume, Z.I.; Mitchell, D.M. Modulation of Retinoid-X-Receptors Differentially Regulates Expression of Apolipoprotein Genes Apoc1 and Apoeb by Zebrafish Microglia. Biol. Open 2022, 11, bio058990. [Google Scholar] [CrossRef]
- Levi, L.; Pekarski, I.; Gutman, E.; Fortina, P.; Hyslop, T.; Biran, J.; Levavi-Sivan, B.; Lubzens, E. Revealing Genes Associated with Vitellogenesis in the Liver of the Zebrafish (Danio rerio) by Transcriptome Profiling. BMC Genomics 2009, 10, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, D.Y.; Howles, P.N. Carboxyl Ester Lipase. J. Lipid Res. 2002, 43, 2017–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Y.; Sun, S.; Yu, X.; Zhou, J.; Cai, W.; Qian, L. Carboxyl Ester Lipase Is Highly Conserved in Utilizing Maternal Supplied Lipids during Early Development of Zebrafish and Human. Biochim. Biophys. Acta BBA-Mol. Cell Biol. Lipids 2020, 1865, 158663. [Google Scholar] [CrossRef]
- Liu, C.; Ye, D.; Wang, H.; He, M.; Sun, Y. Elovl2 But Not Elovl5 Is Essential for the Biosynthesis of Docosahexaenoic Acid (DHA) in Zebrafish: Insight from a Comparative Gene Knockout Study. Mar. Biotechnol. 2020, 22, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, M.S.; Fernández-Palacios, H.; Tacon, A.G.J. Effect of Broodstock Nutrition on Reproductive Performance of Fish. Aquaculture 2001, 197, 25–42. [Google Scholar] [CrossRef]
- Glaser, C.; Heinrich, J.; Koletzko, B. Role of FADS1 and FADS2 Polymorphisms in Polyunsaturated Fatty Acid Metabolism. Metabolism 2010, 59, 993–999. [Google Scholar] [CrossRef]
- Oboh, A.; Kabeya, N.; Carmona-Antoñanzas, G.; Castro, L.F.C.; Dick, J.R.; Tocher, D.R.; Monroig, O. Two Alternative Pathways for Docosahexaenoic Acid (DHA, 22:6n-3) Biosynthesis Are Widespread among Teleost Fish. Sci. Rep. 2017, 7, 3889. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Tan, Q.; Zhang, L.; Yao, J.; Zhou, H.; Hu, P.; Liang, X.; Liu, H. The Migration of Docosahexenoic Acid (DHA) to the Developing Ovary of Female Zebrafish (Danio Rerio). Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 2019, 233, 97–105. [Google Scholar] [CrossRef]
- Laquérriere, A.; Maluenda, J.; Camus, A.; Fontenas, L.; Dieterich, K.; Nolent, F.; Zhou, J.; Monnier, N.; Latour, P.; Gentil, D.; et al. Mutations in CNTNAP1 and ADCY6 Are Responsible for Severe Arthrogryposis Multiplex Congenita with Axoglial Defects. Hum. Mol. Genet. 2014, 23, 2279–2289. [Google Scholar] [CrossRef]
- Clelland, E.; Peng, C. Endocrine/Paracrine Control of Zebrafish Ovarian Development. Mol. Cell. Endocrinol. 2009, 312, 42–52. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Clean Reads | Clean Bases | GC Content | % ≥ Q30 |
|---|---|---|---|---|
| C-1 | 19,651,945 | 5,868,622,898 | 48.21% | 94.49% |
| C-2 | 20,082,524 | 5,996,732,510 | 47.29% | 94.89% |
| C-3 | 20,863,105 | 6,234,771,674 | 47.43% | 94.40% |
| T-1 | 21,676,987 | 6,470,986,448 | 48.56% | 94.72% |
| T-2 | 20,607,217 | 6,158,537,846 | 46.87% | 94.25% |
| T-3 | 19,747,954 | 5,902,895,264 | 46.98% | 95.09% |
| Sample | Total Reads | Mapped Reads | Uniq Mapped Reads | Multiple Map Reads | Reads Map to ‘+’ | Reads Map to ‘−’ |
|---|---|---|---|---|---|---|
| C-1 | 39,303,890 | 36,104,705 (91.86%) | 33,739,060 (85.84%) | 2,365,645 (6.02%) | 19,980,679 (50.84%) | 19,979,652 (50.83%) |
| C-2 | 40,165,048 | 36,322,349 (90.43%) | 34,429,375 (85.72%) | 1,892,974 (4.71%) | 19,675,070 (48.99%) | 19,676,792 (48.99%) |
| C-3 | 41,726,210 | 38,826,693 (93.05%) | 30,444,653 (72.96%) | 8,382,040 (20.09%) | 27,825,562 (66.69%) | 27,711,937 (66.41%) |
| T-1 | 43,353,974 | 39,867,193 (91.96%) | 32,660,972 (75.34%) | 7,206,221 (16.62%) | 27,219,252 (62.78%) | 27,168,567 (62.67%) |
| T-2 | 41,214,434 | 37,442,521 (90.85%) | 34,316,396 (83.26%) | 3,126,125 (7.59%) | 21,448,304 (52.04%) | 21,472,425 (52.10%) |
| T-3 | 39,495,908 | 36,737,497 (93.02%) | 32,701,282 (82.80%) | 4,036,215 (10.22%) | 22,016,744 (55.74%) | 21,991,167 (55.68%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Yang, H.; Li, Y.; Guo, Y.; Li, G.; Chen, H. Comparative Transcriptome Analysis Reveals the Effect of Aurantiochytrium sp. on Gonadal Development in Zebrafish. Animals 2023, 13, 2482. https://doi.org/10.3390/ani13152482
Huang Y, Yang H, Li Y, Guo Y, Li G, Chen H. Comparative Transcriptome Analysis Reveals the Effect of Aurantiochytrium sp. on Gonadal Development in Zebrafish. Animals. 2023; 13(15):2482. https://doi.org/10.3390/ani13152482
Chicago/Turabian StyleHuang, Yanlin, Hao Yang, Yikai Li, Yuwen Guo, Guangli Li, and Huapu Chen. 2023. "Comparative Transcriptome Analysis Reveals the Effect of Aurantiochytrium sp. on Gonadal Development in Zebrafish" Animals 13, no. 15: 2482. https://doi.org/10.3390/ani13152482