

Return to 1616: Multispecies Fauna Reconstruction Requires Thinking Outside the Box
 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Interspecific Interactions
3. Wild-to-Wild vs. Captive Breeding
4. Source Population Viability
5. Release Strategies
6. Stress and Animal Welfare
7. Management of Disease Risk
8. Genetic Management
9. Summary
- -
- Employ the use of decision support tools, including PVAs and DRAs, to inform translocation strategies and identify the most important risks that may need to be mitigated.
- -
- Trial novel and innovative techniques, particularly if there is a likelihood of improved translocation and animal welfare outcomes, ideally within an experimental framework to fully assess their efficacy.
- -
- Proactively communicate findings so that other projects can benefit from the collective learnings of the translocation community, including reasons for failures as well as successes, as these will be of value to those seeking to avoid similar pitfalls. The Return to 1616 project has greatly benefited from the publication of results from other translocations elsewhere.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaywood, M.J.; Stanley-Price, M. Moving Species: Reintroductions and Other Conservation Translocations. In Conservation Translocations; Gaywood, M.J., Ewen, J.G., Hollingsworth, P.M., Moehrenschlager, A., Eds.; Cambridge University Press: Cambridge, UK, 2023; pp. 3–42. [Google Scholar]
- IUCN/SSC. Guidelines for Reintroductions and Other Conservation Translocations. Version 1.0; IUCN Species Survival Commission: Gland, Switzerland, 2013; p. 57. [Google Scholar]
- Morris, S.D.; Brook, B.W.; Moseby, K.E.; Johnson, C.N. Factors affecting success of conservation translocations of terrestrial vertebrates: A global systematic review. Glob. Ecol. Conserv. 2021, 28, e01630. [Google Scholar] [CrossRef]
- Swan, K.D.; McPherson, J.M.; Seddon, P.J.; Moehrenschlager, A. Managing Marine Biodiversity: The Rising Diversity and Prevalence of Marine Conservation Translocations. Conserv. Lett. 2016, 9, 239–251. [Google Scholar] [CrossRef]
- Resende, P.S.; Viana–Junior, A.B.; Young, R.J.; de Azevedo, C.S. A global review of animal translocation programs. Anim. Biodivers. Conserv. 2020, 43, 221–232. [Google Scholar] [CrossRef]
- Doherty, T.S.; Glen, A.S.; Nimmo, D.G.; Ritchie, E.G.; Dickman, C.R. Invasive predators and global biodiversity loss. Proc. Natl. Acad. Sci. USA 2016, 113, 11261–11265. [Google Scholar] [CrossRef] [PubMed]
- Legge, S.; Woinarski, J.C.Z.; Burbidge, A.A.; Palmer, R.; Ringma, J.; Radford, J.Q.; Mitchell, N.; Bode, M.; Wintle, B.; Baseler, M.; et al. Havens for threatened Australian mammals: The contributions of fenced areas and offshore islands to the protection of mammal species susceptible to introduced predators. Wildl. Res. 2018, 45, 627–644. [Google Scholar] [CrossRef]
- Soorae, P.S. (Ed.) Global Conservation Translocation Perspectives: 2021. Case Studies from around the Globe; IUCN SSC Conservation Translocation Specialist Group, Environment Agency—Abu Dhabi and Calgary Zoo, Canada: Gland, Switzerland, 2021. [Google Scholar]
- Berger-Tal, O.; Blumstein, D.T.; Swaisgood, R.R. Conservation translocations: A review of common difficulties and promising directions. Anim. Conserv. 2020, 23, 121–131. [Google Scholar] [CrossRef]
- Morris, K.; Page, M.; Thomas, N.; Ottewell, K. A Strategic Framework for Reconstruction and Conservation of the Vertebrate Fauna of Dirk Hartog Island 2017–2030; Department of Parks and Wildlife: Perth, WA, Australia, 2017. [Google Scholar]
- Kanowski, J. Translocation program set to secure the future of 21 threatened species. Australian Wildlife Conservancy. Wildlife Matters 2019, 38–43. Available online: https://www.australianwildlife.org/translocation-program-set-to-secure-the-future-of-21-threatened-species/ (accessed on 7 July 2023).
- Johnston, G.; Menz, C. An Independent Review of the Evidence under-Pinning the Rewilding of Southern Yorke Peninsula; Northern and Yorke Natural Resource Management Board: Clare, SA, Australia, 2019. [Google Scholar]
- Kingsford, R.T.; West, R.S.; Pedler, R.D.; Keith, D.A.; Moseby, K.E.; Read, J.L.; Letnic, M.; Leggett, K.E.A.; Ryall, S.R. Strategic adaptive management planning—Restoring a desert ecosystem by managing introduced species and native herbivores and reintroducing mammals. Cons. Sci. Pract. 2020, 3, e268. [Google Scholar] [CrossRef]
- Parks Victoria. Wilsons Prom Sanctuary. Available online: https://www.parks.vic.gov.au/get-into-nature/conservation-and-science/conserving-our-parks/wilsons-prom-sanctuary (accessed on 11 May 2023).
- DEC. Dirk Hartog Island National Park Ecological Restoration Strategic Plan; Department of Environment and Conservation: Perth, WA, Australia, 2011. [Google Scholar]
- Heriot, S.; Asher, J.; Williams, M.R.; Moro, D. The eradication of ungulates (sheep and goats) from Dirk Hartog Island, Shark Bay World Heritage Area, Australia. Biol. Invasions 2019, 21, 1789–1805. [Google Scholar] [CrossRef]
- Algar, D.; Johnston, M.; Tiller, C.; Onus, M.; Fletcher, J.; Desmond, G.; Hamilton, N.; Speldewinde, P. Feral cat eradication on Dirk Hartog Island, Western Australia. Biol. Invasions 2019, 22, 1037–1054. [Google Scholar] [CrossRef]
- Astron, E.S. Dirk Hartog Island National Park Weed Management and Action Plan (2013–2018); Prepared for Department of Environment and Conservation: Perth, WA, Australia, 2012. [Google Scholar]
- Algar, D.; Morris, K.; Asher, J.; Cowen, S. Dirk Hartog Island ‘Return to 1616′ Project–The first six years (2014 to 2019). Ecol. Manag. Restor. 2020, 21, 173–183. [Google Scholar] [CrossRef]
- van Dongen, R.; Huntley, B.; Keighery, G.; Brundrett, M. Monitoring vegetation recovery in the early stages of the Dirk Hartog Island Restoration Programme using high temporal frequency Landsat imagery. Ecol. Manag. Restor. 2019, 20, 250–261. [Google Scholar] [CrossRef]
- Burbidge, A.A.; George, A.S. The flora and fauna of Dirk Hartog Island, Western Australia. J. R. Soc. West. 1978, 60, 71–90. [Google Scholar]
- Cowan, P.E.; Tyndale-Biscoe, C.H. Australian and New Zealand mammal species considered to be pests or problems. Reprod. Fertil. Dev. 1997, 9, 27–36. [Google Scholar] [CrossRef]
- Moro, D. Evaluation and cost-benefits of controlling house mice (Mus domesticus) on islands: An example from Thevenard Island, Western Australia. Biol. Conserv. 2001, 99, 355–364. [Google Scholar] [CrossRef]
- Cowen, S.; Rayner, K.; Sims, C.; Van der Weyde, L. Dirk Hartog Island National Park Ecological Restoration Project: Stage Two–Year Four Translocation and Monitoring Report; Department of Biodiversity, Conservation and Attractions: Perth, WA, Australia, 2022. [Google Scholar]
- Cowen, S.; Rayner, K.; Sims, C. Dirk Hartog Island National Park Ecological Restoration Project: Stage Two–Year Three Translocation and Monitoring Report; Department of Biodiversity, Conservation and Attractions: Perth, WA, Australia, 2021. [Google Scholar]
- Cowen, S.; Rayner, K.; Sims, C.; Morris, K. Dirk Hartog Island National Park Ecological Restoration Project: Stage One–Trial Hare-Wallaby Translocations and Monitoring; Department of Biodiversity, Conservation and Attractions: Perth, WA, Australia, 2018. [Google Scholar]
- Baynes, A. The mammals of Shark Bay, Western Australia. In Research in Shark Bay: Report of the France-Australe Bicentenary Expedition Committee; Berry, P.F., Bradshaw, S.D., Wilson, B.R., Eds.; Western Australian Museum: Perth, WA, Australia, 1990. [Google Scholar]
- Baynes, A. The Original Mammal Fauna of Dirk Hartog Island; Department of Environment and Conservation: Perth, WA, Australia, 2008. [Google Scholar]
- Shortridge, G.C. An Account of the Geographical Distribution of the Marsupials and Monotremes of South-West Australia, having special reference to the specimens collected during the Balston Expedition of 1904–1907. Proc. Zool. Soc. Lond. 1909, 79, 803–848. [Google Scholar] [CrossRef]
- Orchard, A.; Orchard, T. The Australian Botanical Journals of Allan Cunningham. The Later King Expeditions February 1819–September; Privately Published: Western Creek, ACT, Australia, 2018. [Google Scholar]
- Whitlock, F.L. Note on Dirk Hartog Island and Peron Peninsula, Shark Bay, Western Australia. Emu 1920, 20, 168–186. [Google Scholar] [CrossRef]
- Black, A.B.; Burbidge, A.H.; Garnett, S.T. Dirk Hartog Island Western Grasswren Amytornis textilis carteri. In The Action Plan for Australian Birds 2020; Garnett, S.T., Baker, G.B., Eds.; CSIRO Publishing: Melbourne, VIC, Australia, 2021; pp. 562–563. [Google Scholar]
- Davies, S.J.J.F.; Chapman, G.S. The Status of Birds on Peron Peninsula and Dirk Hartog Island, Shark Bay, Wa. Emu 2016, 75, 55–61. [Google Scholar] [CrossRef]
- Short, J. The Characteristics and Success of Vertebrate Translocations within Australia; Department of Agriculture, Fisheries and Forestry: Canberra, Australia, 2009. [Google Scholar]
- Palmer, B.J.; Valentine, L.E.; Page, M.; Hobbs, R.J. Translocations of digging mammals and their potential for ecosystem restoration: A review of goals and monitoring programmes. Mammal Rev. 2020, 50, 382–398. [Google Scholar] [CrossRef]
- Short, J.; Bradshaw, S.D.; Giles, J.; Prince, R.I.T.; Wilson, G.R. Reintroduction of macropods (Marsupialia: Macropodoidea) in Australia—A review. Biol. Conserv. 1992, 62, 189–204. [Google Scholar] [CrossRef]
- Fischer, J.; Lindenmayer, D.B. An assessment of the published results of animal relocations. Biol. Conserv. 2000, 96, 1–11. [Google Scholar] [CrossRef]
- Peterson, K.A.; Barnes, M.D.; Jeynes-Smith, C.; Cowen, S.; Gibson, L.; Sims, C.; Baker, C.M.; Bode, M. Reconstructing lost ecosystems: A risk analysis framework for planning multispecies reintroductions under severe uncertainty. J. Appl. Ecol. 2021, 58, 2171–2184. [Google Scholar] [CrossRef]
- Cowan, M. Reptiles of Dirk Hartog Island; Department of Biodiversity, Conservation and Attractions: Perth, WA, Australia, 2018. [Google Scholar]
- Seddon, P.J.; Armstrong, D.P.; Maloney, R.F. Developing the science of reintroduction biology. Conserv. Biol. 2007, 21, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Griffith, B.; Scott, J.M.; Carpenter, J.W.; Reed, C. Translocation as a Species Conservation Tool: Status and Strategy. Science 1989, 245, 477–480. [Google Scholar] [CrossRef]
- Robert, A. Captive breeding genetics and reintroduction success. Biol. Conserv. 2009, 142, 2915–2922. [Google Scholar] [CrossRef]
- Witzenberger, K.A.; Hochkirch, A. Ex situ conservation genetics: A review of molecular studies on the genetic consequences of captive breeding programmes for endangered animal species. Biodivers. Conserv. 2011, 20, 1843–1861. [Google Scholar] [CrossRef]
- Snyder, N.F.R.; Derrickson, S.R.; Beissinger, S.R.; Wiley, J.W.; Smith, T.B.; Toone, W.D.; Miller, B. Limitations of Captive Breeding in Endangered Species Recovery. Conserv. Biol. 1996, 10, 338–348. [Google Scholar] [CrossRef]
- Carrete, M.; Tella, J.L. Rapid loss of antipredatory behaviour in captive-bred birds is linked to current avian invasions. Sci. Rep. 2015, 5, 18274. [Google Scholar] [CrossRef]
- Ballou, J.D. Assessing the risks of infectious diseases in captive breeding and reintroduction programs. J. Zoo Wildl. Med. 1993, 24, 327–335. [Google Scholar]
- Cunningham, A.A. Disease Risks of Wildlife Translocations. Conserv. Biol. 1996, 10, 349–353. [Google Scholar] [CrossRef]
- Griffith, B.; Scott, J.M.; Carpenter, J.W.; Reed, C. Translocations of captive-reared terrestrial vertebrates 1973–1986. Endanger. Species Update 1990, 8, 10–14. [Google Scholar]
- Palmer, N.; Smith, M.J.; Ruykys, L.; Jackson, C.; Volck, G.; Riessen, N.; Thomasz, A.; Moir, C.; Palmer, B. Wild-born versus captive-bred: A comparison of survival and refuge selection by translocated numbats (Myrmecobius fasciatus). Wildl. Res. 2020, 47, 217–223. [Google Scholar] [CrossRef]
- Richards, J.D. Western Barred Bandicoot Perameles Bougainville, Burrowing Bettong Bettongia Lesueur and Banded Hare-Wallaby Lagostrophus Fasciatus National Recovery Plan, Wildlife Management Program No. 49; Department of Environment and Conservation: Perth, WA, Australia, 2012. [Google Scholar]
- Lambert, C.; Power, V.; Gaikhorst, G. Captive breeding of the Shark Bay mouse Pseudomys fieldi to facilitate species recovery in the wild. J. Zoo Aquar. Res. 2016, 4, 164–168. [Google Scholar]
- Lambert, C.; Mills, H. Husbandry and breeding of the Dibbler. Int. Zoo Yearb. 2006, 40, 290–301. [Google Scholar] [CrossRef]
- Short, J.; Copley, P.; Ruykys, L.; Morris, K.; Read, J.; Moseby, K. Review of translocations of the greater stick-nest rat (Leporillus conditor): Lessons learnt to facilitate ongoing recovery. Wildl. Res. 2019, 46, 455–475. [Google Scholar] [CrossRef]
- Hardman, B.; Moro, D. Optimising reintroduction success by delayed dispersal: Is the release protocol important for hare-wallabies? Biol. Conserv. 2006, 128, 403–411. [Google Scholar] [CrossRef]
- Mills, H.R.; Moro, D.; Spencer, P.B.S. Conservation significance of island versus mainland populations: A case study of dibblers (Parantechinus apicalis) in Western Australia. Anim. Conserv. 2004, 7, 387–395. [Google Scholar] [CrossRef]
- Northover, A.; Friend, T.; Cowen, S.; Ottewell, K.; Mills, H.; Moro, D.; Gibson, L. Recovery of the Dibbler Parantechinus Apicalis—Review of Progress and Future Research Directions; Department of Biodiversity, Conservation and Attractions: Perth, WA, Australia, 2022. [Google Scholar]
- Aisya, Z.; White, D.J.; Thavornkanlapachai, R.; Friend, J.A.; Rick, K.; Mitchell, N.J. Using PVA and captive breeding to balance trade-offs in the rescue of the island dibbler onto a new island ark. Sci. Rep. 2022, 12, 11913. [Google Scholar] [CrossRef] [PubMed]
- Parrott, M.L.; Ward, S.J.; Temple-Smith, P.D.; Selwood, L. Females Choose Mates Based on Genetic Relatedness in a Small Dasyurid Marsupial, the Agile Antechinus (Antechinus agilis). PLoS ONE 2015, 10, e0122381. [Google Scholar] [CrossRef]
- Bright, P.W.; Morris, P.A. Animal Translocation for Conservation: Performance of Dormice in Relation to Release Methods, Origin and Season. J. Appl. Ecol. 1994, 31, 699–708. [Google Scholar] [CrossRef]
- McClelland, P.; Gummer, H. Reintroduction of the critically endangered Campbell Island teal Anas nesiotis to Campbell Island, New Zealand. Conserv. Evid. 2006, 3, 61–63. [Google Scholar]
- Friend, T. Medium-Term Radio-Tracking of Dibblers, Fitzgerald River NP-Bankwest Landscope Conservation Visacard, Final Report; Department of Conservation and Land Management: Perth, WA, Australia, 2001. [Google Scholar]
- Mitchell, W.F.; Boulton, R.L.; Sunnucks, P.; Clarke, R.H. Are we adequately assessing the demographic impacts of harvesting for wild-sourced conservation translocations? Conserv. Sci. Pract. 2021, 4, e569. [Google Scholar] [CrossRef]
- Sims, C.; Cowen, S.; Friend, J.A. Monitoring Source Populations of Fauna for the Dirk Hartog Island National Park Ecological Restoration Project–2018; Department of Biodiversity Conservation and Attractions: Perth, WA, Australia, 2019. [Google Scholar]
- Prieto Gonzalez, R.; Thomas, L.; Marques, T.A. Estimation bias under model selection for distance sampling detection functions. Environ. Ecol. Stat. 2017, 24, 399–414. [Google Scholar] [CrossRef]
- Buckland, S.T.; Anderson, D.R.; Burnham, K.P.; Laake, J.L.; Borchers, D.L.; Thomas, L. Introduction to Distance Sampling: Estimating Abundance of Biological Populations; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Short, J.; Turner, B. The distribution and abundance of the banded and rufous hare-wallabies, Lagostrophus fasciatus and Lagorchestes hirsutus. Biol. Conserv. 1992, 60, 157–166. [Google Scholar] [CrossRef]
- Sims, C.; Cowen, S. Monitoring Fauna Source Populations for the Dirk Hartog Island National Park Ecological Restoration Project 2017/18; Department of Biodiversity Conservation and Attractions: Perth, WA, Australia, 2018. [Google Scholar]
- Sims, C.; Cowen, S.; Friend, J.A.; Comer, S.; Gibson Vega, A.; Garretson, S.; Gibson, L. Monitoring Source Populations of Fauna for the Dirk Hartog Island National Park Ecological Restoration Project–2021; Department of Biodiversity Conservation and Attractions: Perth, WA, Australia, 2022. [Google Scholar]
- Sims, C.; Cowen, S.; Garretson, S.; Friend, J.A. Monitoring Source Populations of Fauna for the Dirk Hartog Island National Park Ecological Restoration Project–2019; Department of Biodiversity Conservation and Attractions: Perth, WA, Australia, 2020. [Google Scholar]
- Sims, C.; Cowen, S.; Garretson, S.; Friend, J.A. Monitoring Source Populations of Fauna for the Dirk Hartog Island National Park Ecological Restoration Project–2020; Department of Biodiversity Conservation and Attractions: Perth, WA, Australia, 2021. [Google Scholar]
- Lacy, R.C.; Pollak, J.P. Vortex: A Stochastic Simulation of the Extinction Process; Version 10.5.5; Chicago Zoological Society: Brookfield, IL, USA, 2021. [Google Scholar]
- Chaudhary, V.; Oli, M.K. A critical appraisal of population viability analysis. Conserv. Biol. 2020, 34, 26–40. [Google Scholar] [CrossRef] [PubMed]
- White, D.J.; Ottewell, K.; Spencer, P.B.S.; Smith, M.; Short, J.; Sims, C.; Mitchell, N.J. Genetic Consequences of Multiple Translocations of the Banded Hare-Wallaby in Western Australia. Diversity 2020, 12, 448. [Google Scholar] [CrossRef]
- White, D.; Hogg, C. Shark Bay Bandicoot Management (Preliminary Report Subject to Further Sensitivity Testing); NESP Threatened Species Recovery Hub: Australian Government, ACT, Australia, 2019; Available online: https://www.nespthreatenedspecies.edu.au/publications-and-tools/shark-bay-bandicoot-management-preliminary-report-subject-to-further-sensitivity-testing (accessed on 7 July 2023).
- Quah, R.J.J.-E. Conservation Genetics and Population Modelling to Secure Wild Populations of Djoongari (Pseudomys fieldi); University of Western Australia: Perth, WA, Australia, 2021. [Google Scholar]
- Onley, I.R.; White, L.C.; Moseby, K.E.; Copley, P.; Cowen, S. Disproportionate admixture improves reintroduction outcomes despite the use of low-diversity source populations: Population viability analysis for a translocation of the greater stick-nest rat. Anim. Conserv. 2023, 26, 216–227. [Google Scholar] [CrossRef]
- White, L.C.; Thomson, V.A.; West, R.; Ruykys, L.; Ottewell, K.; Kanowski, J.; Moseby, K.E.; Byrne, M.; Donnellan, S.C.; Copley, P.; et al. Genetic monitoring of the greater stick-nest rat meta-population for strategic supplementation planning. Conserv. Genet. 2020, 21, 941–956. [Google Scholar] [CrossRef]
- Ewen, J.G.; Canessa, S.; Converse, S.J.; Parker, K.A. Decision-Making in Animal Conservation Translocations: Biological Considerations and Beyond. In Conservation Translocations; Gaywood, M.J., Ewen, J.G., Hollingsworth, P.M., Moehrenschlager, A., Eds.; Cambridge University Press: Cambridge, UK, 2023; pp. 108–148. [Google Scholar]
- Letty, J.; Marchandeau, S.; Aubineau, J. Problems encountered by individuals in animal translocations: Lessons from field studies. Ecoscience 2007, 14, 420–431. [Google Scholar] [CrossRef]
- de Milliano, J.; Di Stefano, J.; Courtney, P.; Temple-Smith, P.; Coulson, G. Soft-release versus hard-release for reintroduction of an endangered species: An experimental comparison using eastern barred bandicoots (Perameles gunnii). Wildl. Res. 2016, 43, 1–12. [Google Scholar] [CrossRef]
- Bubac, C.M.; Johnson, A.C.; Fox, J.A.; Cullingham, C.I. Conservation translocations and post-release monitoring: Identifying trends in failures, biases, and challenges from around the world. Biol. Conserv. 2019, 238, 108239. [Google Scholar] [CrossRef]
- Johnson, T.F.; Brown, T.J.; Richardson, D.S.; Dugdale, H.L. The importance of post-translocation monitoring of habitat use and population growth: Insights from a Seychelles Warbler (Acrocephalus sechellensis) translocation. J. Ornithol. 2017, 159, 439–446. [Google Scholar] [CrossRef]
- Parker, K.A.; Ewen, J.G.; Seddon, P.J.; Armstrong, D.P. Post-release monitoring of bird translocations. Notornis 2013, 60, 85–92. [Google Scholar]
- Jensen, M.A.; Paton, D.C.; Moseby, K.E. Delayed release improves site fidelity but has little effect on survival or breeding success of reintroduced western quolls (Dasyurus geoffroii). Austral Ecol. 2021, 46, 1298–1310. [Google Scholar] [CrossRef]
- Wilson, B.A.; Evans, M.J.; Batson, W.G.; Banks, S.C.; Gordon, I.J.; Fletcher, D.B.; Wimpenny, C.; Newport, J.; Belton, E.; Rypalski, A.; et al. Adapting reintroduction tactics in successive trials increases the likelihood of establishment for an endangered carnivore in a fenced sanctuary. PLoS ONE 2020, 15, e0234455. [Google Scholar] [CrossRef] [PubMed]
- Tetzlaff, S.J.; Sperry, J.H.; DeGregorio, B.A. Effects of antipredator training, environmental enrichment, and soft release on wildlife translocations: A review and meta-analysis. Biol. Conserv. 2019, 236, 324–331. [Google Scholar] [CrossRef]
- Cowen, S.; Rayner, K.; Scheelen, L.; Sims, C.; Gibson, L. Dirk Hartog Island National Park Ecological Restoration Project: Stage Two–Year One Translocation and Monitoring Report; Department of Biodiversity, Conservation and Attractions: Perth, WA, Australia, 2019. [Google Scholar]
- Cowen, S.; Rayner, K.; Sims, C.; Friend, T.; Knox, F.; Ottewell, K.; Gibson, L. Dirk Hartog Island National Park Ecological Restoration Project: Stage Two–Year Two Translocation and Monitoring Report; Department of Biodiversity, Conservation and Attractions: Perth, WA, Australia, 2020. [Google Scholar]
- Bilby, J.; Moseby, K. Review of hyperdispersal in wildlife translocations. Conserv. Biol. 2023, e14083. [Google Scholar] [CrossRef]
- Copley, P. Natural histories of Australia’s stick-nest rats, genus Leporillus (Rodentia: Muridae). Wildl. Res. 1999, 26, 513–539. [Google Scholar] [CrossRef]
- Onley, I.R. Conservation Genomics and Adaptive Management of Translocated Greater Stick-Nest Rats (Leporillus conditor) under Climate Change; University of Adelaide: Adelaide, SA, Australia, 2022. [Google Scholar]
- Lambert, C. Animal Management Guidelines Dibbler (Parantechinus apicalis); Native Species Breeding Program, Department of Animal Health and Research, Perth Zoo: Perth, WA, Australia, 2012. [Google Scholar]
- John, S. Invertebrate Abundance on Dirk Hartog Island, Western Australia: Food Availability for Dibblers Parantechinus apicalis; Curtin University: Perth, WA, Australia, 2018. [Google Scholar]
- Dickens, M.J.; Delehanty, D.J.; Michael Romero, L. Stress: An inevitable component of animal translocation. Biol. Conserv. 2010, 143, 1329–1341. [Google Scholar] [CrossRef]
- Dickens, M.J.; Delehanty, D.J.; Romero, L.M. Stress and translocation: Alterations in the stress physiology of translocated birds. Proc. Biol. Sci. 2009, 276, 2051–2056. [Google Scholar] [CrossRef]
- Green-Barber, J.M.; Stannard, H.J.; Old, J.M. A suspected case of myopathy in a free-ranging eastern grey kangaroo (Macropus giganteus). Aust. Mammal. 2018, 40, 122–126. [Google Scholar] [CrossRef]
- Cowan, M.; Blythman, M.; Angus, J.; Gibson, L. Post-Release Monitoring of Western Grey Kangaroos (Macropus fuliginosus) Relocated from an Urban Development Site. Animals 2020, 10, 1914. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.R.; Langford, D.G.; Gibson, D.F. Capture myopathy in Lagorchestes hirsutus (Marsupialia: Macropodidae). Aust. Mammal. 1994, 17, 137–138. [Google Scholar] [CrossRef]
- Langford, D.; Burbidge, A.A. Translocation of mala (Lagorchestes hirsutus) from the Tanami Desert, Northern Territory to Trimouille Island, Western Australia. Aust. Mammal. 2001, 23, 37–46. [Google Scholar] [CrossRef]
- Breed, D.; Meyer, L.C.R.; Steyl, J.C.A.; Goddard, A.; Burroughs, R.; Kohn, T.A. Conserving wildlife in a changing world: Understanding capture myopathy-a malignant outcome of stress during capture and translocation. Conserv. Physiol. 2019, 7, coz027. [Google Scholar] [CrossRef]
- Baldwin, R.A.; Meinerz, R.; Jantz, H.E.; Witmer, G.W. Impact of Capture and Transportation Methods on Survival of Small Rodents During Relocation Events. Southwest. Nat. 2016, 60, 385–389. [Google Scholar] [CrossRef]
- Narayan, E.J. Evaluation of physiological stress in Australian wildlife: Embracing pioneering and current knowledge as a guide to future research directions. Gen. Comp. Endocrinol. 2017, 244, 30–39. [Google Scholar] [CrossRef]
- Chipman, R.; Slate, D.; Rupprecht, C.; Mendoza, M. Downside Risk of Wildlife Translocation; USDA National Wildlife Research Center-Staff Publications. 1896. U.S. Department of Agriculture: Riverdale, MD, USA, 2008. [Google Scholar]
- Portas, T. Veterinary aspects of native mammal conservation translocations. In Current Therapy in Medicine of Australian Mammals; Vogelnest, L., Portas, T., Eds.; CSIRO Publishing: Clayton South, VIT, Australia, 2019; pp. 17–35. [Google Scholar]
- OIE; IUCN. Guidelines for Wildlife Disease Risk Analysis; Published in Association with the IUCN and the Species Survival Commission; OIE: Paris, France, 2014; p. 24. [Google Scholar]
- Vaughan-Higgins, R.; Vitali, S.; Reiss, A. Western Barred Bandicoot (Perameles bougainville) Proposed Translocation: Disease Risk Analysis (DRA); Department of Environment and Conservation: Perth, WA, Australia, 2018. [Google Scholar]
- Vaughan-Higgins, R.J.; Vitali, S.D.; Sims, C.; Page, M.; Reiss, A. Streamlining Disease Risk Analysis for Wildlife Using the Shark Bay Bandicoot as a Model. Ecohealth 2021, 18, 13–30. [Google Scholar] [CrossRef]
- Travouillon, K.J.; Phillips, M.J. Total evidence analysis of the phylogenetic relationships of bandicoots and bilbies (Marsupialia: Peramelemorphia): Reassessment of two species and description of a new species. Zootaxa 2018, 4378, 224–256. [Google Scholar] [CrossRef]
- Woolford, L.; Rector, A.; Van Ranst, M.; Ducki, A.; Bennett, M.D.; Nicholls, P.K.; Warren, K.S.; Swan, R.A.; Wilcox, G.E.; O’Hara, A.J. A novel virus detected in papillomas and carcinomas of the endangered western barred bandicoot (Perameles bougainville) exhibits genomic features of both the Papillomaviridae and Polyomaviridae. J. Virol. 2007, 81, 13280–13290. [Google Scholar] [CrossRef]
- Woolford, L.; Bennett, M.D.; Sims, C.; Thomas, N.; Friend, J.A.; Nicholls, P.K.; Warren, K.S.; O’Hara, A.J. Prevalence, emergence, and factors associated with a viral papillomatosis and carcinomatosis syndrome in wild, reintroduced, and captive western barred bandicoots (Perameles bougainville). Ecohealth 2009, 6, 414–425. [Google Scholar] [CrossRef]
- Warren, K.; Swan, R.; Bodetti, T.; Friend, T.; Hill, S.; Timms, P. Ocular Chlamydial infections of Western Barred Bandicoot (Perameles bougainville) in Western Australia. J. Zoo Wildl. Med. 2005, 36, 100–102. [Google Scholar] [CrossRef] [PubMed]
- Moore, P. Disease Risk Analysis to Support the Translocation of Dibblers (Parantechinus apicalis) from the Jurien Bay Islands to Dirk Hartog Island National Park (WA); College of Veterinary Medicine, Murdoch University: Perth, WA, Australia, 2019. [Google Scholar]
- Atkins, M.J. A Disease Risk Analysis for the Translocation of Western Grasswrens (Amytornis textilis textilis) from Shark Bay to Dirk Hartog Island; School of Veterinary Medicine, Murdoch University: Perth, WA, Australia, 2022. [Google Scholar]
- Bowry, C. The Overlooked Diversity during Translocations: Demodex spp. of dibblers (Parantechinus apicalis). Masters’s Thesis, Murdoch University, Perth, WA, Australia, 2021. [Google Scholar]
- Sainsbury, A.W.; Carraro, C. Animal Disease and Conservation Translocations. In Conservation Translocations; Gaywood, M.J., Ewen, J.G., Hollingsworth, P.M., Moehrenschlager, A., Eds.; Cambridge University Press: Cambridge, UK, 2023; pp. 149–179. [Google Scholar]
- Vaughan-Higgins, R.J.; Masters, N.; Sainsbury, A.W. Biosecurity for conservation translocations: Fisher’s estuarine moth (Gortyna borelii lunata), short-haired bumblebee (Bombus subterraneus), pool frog (Pelophylax lessonae) and cirl bunting (Emberiza cirlus) translocations as case studies. Ecohealth 2017, 14, 84–91. [Google Scholar] [CrossRef]
- Batson, W.G.; Gordon, I.J.; Fletcher, D.B.; Portas, T.J.; Manning, A.D. The effect of pre-release captivity on the stress physiology of a reintroduced population of wild eastern bettongs. J. Zool. 2017, 303, 311–319. [Google Scholar] [CrossRef]
- Fischer, C.P.; Romero, L.M. Chronic captivity stress in wild animals is highly species-specific. Conserv. Physiol. 2019, 7, coz093. [Google Scholar] [CrossRef] [PubMed]
- Kock, R.A.; Woodford, M.H.; Rossiter, P.B. Disease risks associated with the translocation of wildlife. Rev.-Off. Int. Epizoot. 2010, 29, 329–350. [Google Scholar] [CrossRef] [PubMed]
- Frankham, R. Genetics and extinction. Biol. Conserv. 2005, 126, 131–140. [Google Scholar] [CrossRef]
- Frankham, R.; Bradshaw, C.J.A.; Brook, B.W. Genetics in conservation management: Revised recommendations for the 50/500 rules, Red List criteria and population viability analyses. Biol. Conserv. 2014, 170, 56–63. [Google Scholar] [CrossRef]
- Weeks, A.R.; Moro, D.; Thavornkanlapachai, R.; Taylor, H.R.; White, N.E.; Weiser, E.L.; Heinze, D. Conserving and enhancing genetic diversity in translocation programs. In Advances in Reintroduction Biology of Australian and New Zealand Fauna; Armstrong, D., Hayward, M., Moro, D., Seddon, P., Eds.; CSIRO Publishing, Australia: Clayton South, VIC, Australia, 2015. [Google Scholar]
- Weeks, A.R.; Sgrò, C.M.; Young, A.G.; Frankham, R.; Mitchell, N.J.; Miller, K.A.; Byrne, M.; Coates, D.J.; Eldridge, M.D.; Sunnucks, P.; et al. Assessing the benefits and risks of translocations in changing environments: A genetic perspective. Evol. Appl. 2011, 4, 709–725. [Google Scholar] [CrossRef]
- Eldridge, M.D.B.; Neaves, L.E.; Spencer, P.B.S. Genetic analysis of three remnant populations of the rufous hare-wallaby (Lagorchestes hirsutus) in arid Australia. Aust. Mammal. 2019, 41, 123–131. [Google Scholar] [CrossRef]
- Eldridge, M.D.B.; Kinnear, J.E.; Zenger, K.R.; McKenzie, L.M.; Spencer, P.B.S. Genetic diversity in remnant mainland and ‘‘pristine’’ island populations of three endemic Australian macropodids (Marsupialia): Macropus eugenii, Lagorchestes hirsutus and Petrogale lateralis. Conserv. Genet. 2004, 5, 325–338. [Google Scholar] [CrossRef]
- White, L.C.; Moseby, K.E.; Thomson, V.A.; Donnellan, S.C.; Austin, J.J. Long-term genetic consequences of mammal reintroductions into an Australian conservation reserve. Biol. Conserv. 2018, 219, 1–11. [Google Scholar] [CrossRef]
- Gibson Vega, A.; Hall, M.L.; Ridley, A.; Cowen, S.J.; Slender, A.L.; Burbidge, A.H.; Louter, M.; Kennington, W.J. Population genetic structure associated with a landscape barrier in the Western Grasswren (Amytornis textilis textilis). Ibis 2023. [Google Scholar] [CrossRef]
- Cowen, S.; Smith, M.; McArthur, S.; Rayner, K.; Jackson, C.; Anderson, G.; Ottewell, K. Novel microsatellites and investigation of faecal DNA as a non-invasive population monitoring tool for the banded hare-wallaby. Aust. J. Zool. 2022, 69, 55–66. [Google Scholar] [CrossRef]
- Dziminski, M.A.; Carpenter, F.M.; Morris, F. Monitoring the Abundance of Wild and Reintroduced Bilby Populations. J. Wildl. Manag. 2020, 85, 240–253. [Google Scholar] [CrossRef]
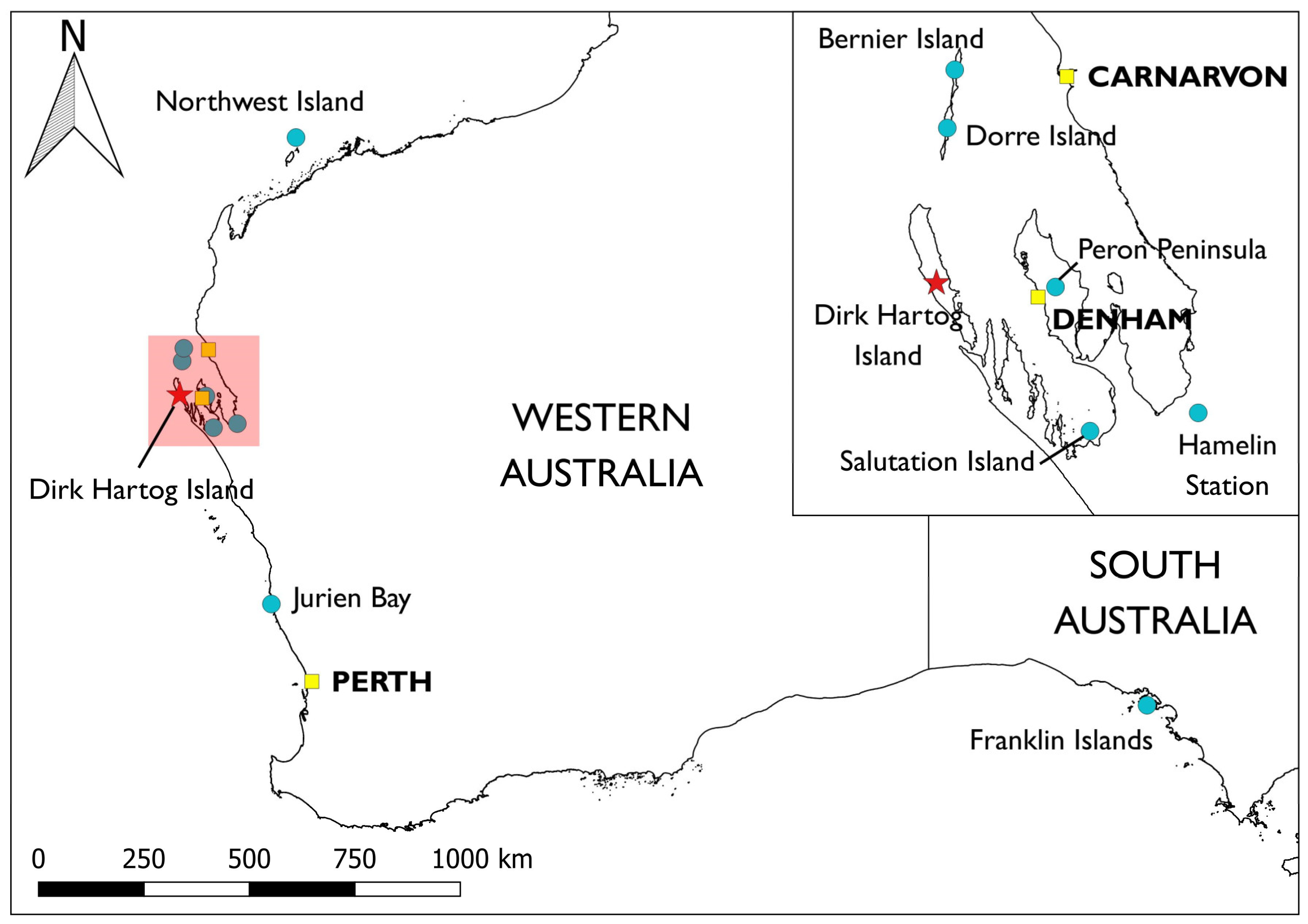

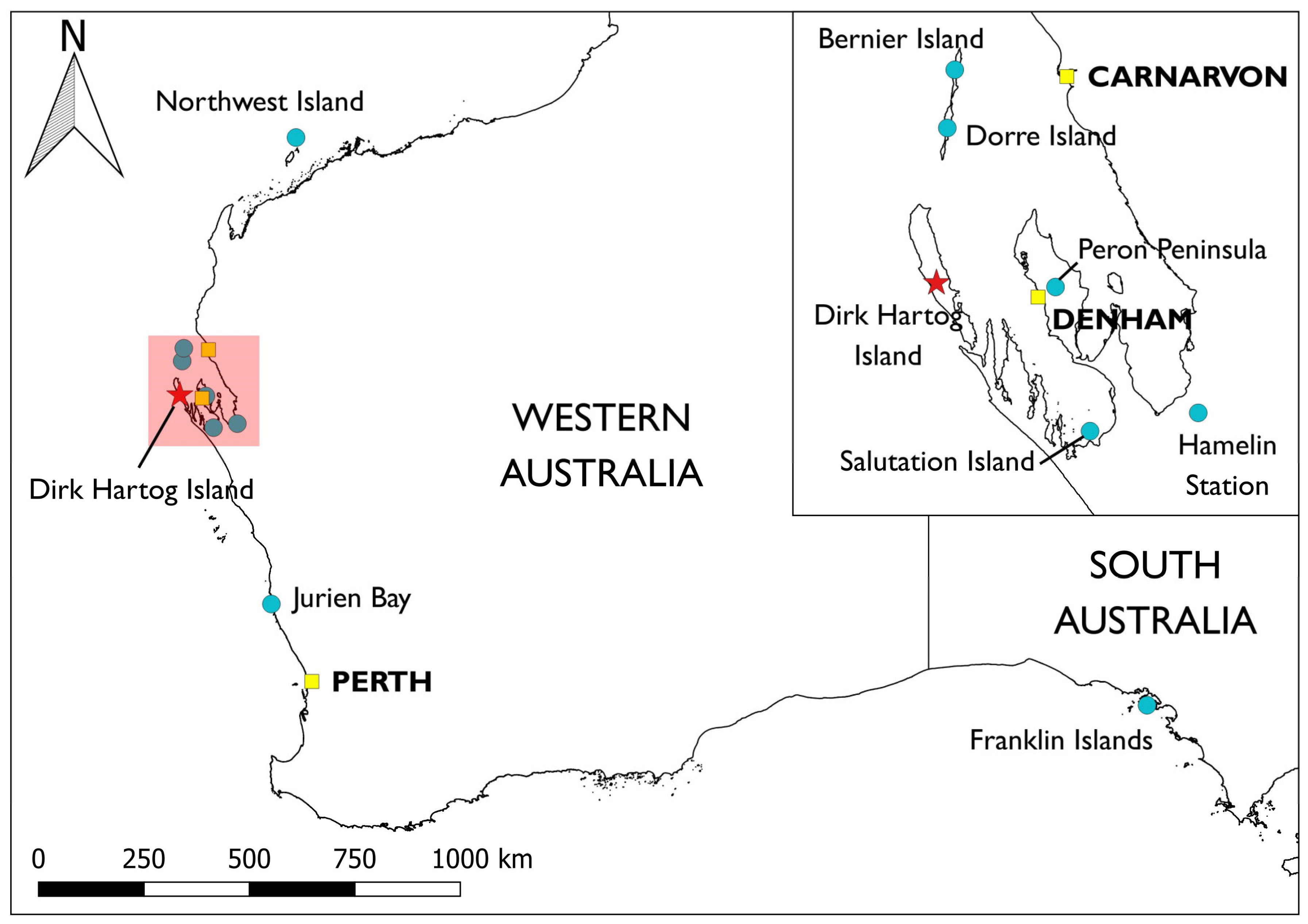{kind=link}
| Common Name | Scientific Name | Listing | Years | Released | Source(s) | PVA | DRA |
|---|---|---|---|---|---|---|---|
| Banded hare-wallaby * | Lagostrophus fasciatus | V | 2017–18 | 102 H | Bernier Is.; Dorre Is. | Y | N |
| Rufous hare-wallaby * | Lagorchestes hirsutus | V | 2017–19 | 112 H | Bernier Is.; Dorre Is. | N | N |
| Shark Bay bandicoot | Perameles bougainville | E | 2019–20 | 99 H | Bernier Is.; Dorre Is. | Y | Y |
| Dibbler | Parantechinus apicalis | E | 2019– | 137 ‡,M | (Boullanger Is.; Escape Is.; Whitlock Is.) via Perth Zoo | Y | Y |
| Shark Bay mouse | Pseudomys gouldii | V | 2021–22 | 130 A | Bernier Is.; Northwest Is. | Y | Y |
| Greater stick-nest rat | Leporillus conditor | V | 2021–22 | 122 A | East Franklin Is.; West Franklin Is.; Salutation Is. | Y | Y |
| Western grasswren † | Amytornis textilis | N | 2022 | 85 H | Hamelin Station; Peron Peninsula | Y | Y |
| Brush-tailed mulgara | Dasycercus blythi | N | 2023 | 100 M | Matuwa Kurrara Kurrara | Y | N § |
| Desert Mouse | Pseudomys desertor | N | - | - | - | - | Y |
| Heath Mouse | Pseudomys shortridgei | E | - | - | - | - | Y |
| Woylie | Bettongia penicillata | E | - | - | - | - | - |
| Boodie | Bettongia lesueur | V | - | - | - | - | - |
| Chuditch | Dasyurus geoffroii | V | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cowen, S.; Sims, C.; Ottewell, K.; Knox, F.; Friend, T.; Mills, H.; Garretson, S.; Rayner, K.; Gibson, L. Return to 1616: Multispecies Fauna Reconstruction Requires Thinking Outside the Box. Animals 2023, 13, 2762. https://doi.org/10.3390/ani13172762
Cowen S, Sims C, Ottewell K, Knox F, Friend T, Mills H, Garretson S, Rayner K, Gibson L. Return to 1616: Multispecies Fauna Reconstruction Requires Thinking Outside the Box. Animals. 2023; 13(17):2762. https://doi.org/10.3390/ani13172762
Chicago/Turabian StyleCowen, Saul, Colleen Sims, Kym Ottewell, Fiona Knox, Tony Friend, Harriet Mills, Sean Garretson, Kelly Rayner, and Lesley Gibson. 2023. "Return to 1616: Multispecies Fauna Reconstruction Requires Thinking Outside the Box" Animals 13, no. 17: 2762. https://doi.org/10.3390/ani13172762