1. Introduction
The species–environment relationships determining the current distribution of endangered species within their geographic range are a key ecological process; therefore, examining and understanding these species–environment relationships may be essential for the development of effective conservation strategies devoted to recovering endangered species [
1,
2,
3]. However, the study of the distribution patterns of large long-lived species presents exceptional challenges, as it requires the integration of spatial and temporal shifts in abundances [
4,
5]. Furthermore, species–environment relationships depend greatly on the scale at which they are studied [
6,
7,
8], and the neglect of appropriate spatial and temporal considerations can lead to inaccuracies in forecasts of species distribution [
9]. Species distribution is a dynamic phenomenon, characterized by spatial contractions and expansions over time, that is influenced by the interplay of biological, ecological, and biogeographic factors. In this context, the application of species distribution models (SDMs; see review [
10]) has been widely used to study species occupancy and abundance patterns.
SDMs empirically examine species occupancy or abundance using grid-cells and the species–environment relationship in terms of intrinsic and extrinsic factors. Despite the advantages of using such methods (e.g., identifying important areas for species conservation; [
11]), the consideration of variability in the temporal dimension is rarely addressed. The incorporation of the continuous temporal dimension (i.e., neither a static nor specific time interval; [
10,
12]) is a novel approach that allows us to use distinct ecological processes and time-dependent factors governing fluctuations in occupancy and abundance [
13,
14,
15]. In addition, since species distribution patterns are also sensitive to factors operating in the local environment such as microclimate or food availability [
16] that differ greatly from those at work at larger scales [
6], SDMs require a specific spatial scale or scales under scrutiny. Additionally, landscape heterogeneity in terms of the availability of suitable breeding sites may also influence occupancy and abundance patterns [
17], thereby promoting spatial aggregation and uneven distribution across a landscape [
18,
19].
Studying the occupancy and abundance distribution of large long-lived vertebrate species presents numerous challenges due to their wide range of different behaviors that require large interconnected habitats [
20,
21,
22]. In this context, vultures are no exception, and their spatial and temporal distribution is often influenced by multiple, often environmental [
23] and human-related [
24,
25] factors whose impact varies depending on scale. As long-lived birds, they exhibit late maturity and low reproductive rates, which leads to slow natural changes in population numbers over time [
26].
Here, we use a novel approach to analyze the factors that influence temporal and spatial variation in the abundance distribution of breeding pairs of the long-lived Egyptian vulture (
Neophron percnopterus), a species threatened worldwide at different local (i.e., specific 100 km
2 areas within a landscape) and regional (i.e., larger geographic regions such as countries) spatial scales. Despite the crucial role that Egyptian vultures play in ecosystem health, they face threats such as habitat loss, persecution, electrocution, and poisoning [
27]. In Spain, human activities have resulted in local extinctions [
28] but, interestingly, in some regions the number of breeding territories is now increasing [
29,
30]. We used long-term Egyptian vulture monitoring information in one of this vulture’s main breeding areas. We aimed (1) to test whether or not Egyptian vulture occupancy and abundance has changed over time in continental Spain; (2) to determine the spatial patterns, i.e., the spatial heterogeneity, of the abundance of breeding territories in the study region; (3) to identify the factors contributing to spatial variation at the local scale; and (4) to investigate the factors responsible for changes in abundance over both time and space at the regional scale. Based on the hypothesis that both temporal and spatial factors influence species distribution, we predicted that the abundance of breeding pairs of Egyptian vultures would vary over time (i.e., a non-stationary distribution) and space (i.e., an aggregated distribution). Furthermore, we anticipated that the factors driving this species’ distribution would differ depending on the spatial scale employed [
6]. The findings of this study will help develop targeted conservation plans for declining vulture populations and facilitate efforts to increase the occupancy rate of their breeding populations.
4. Discussion
This study presents an analysis of the temporal and spatial variation, as well as the distribution patterns at different scales, of breeding pairs of the endangered Egyptian vulture in one of its global strongholds. Our findings revealed stability in its breeding population in continental Spain after years of continuous decline [
31,
37]. However, despite this regional stability, significant spatiotemporal variation occurred. The distribution of Egyptian vultures exhibited an aggregated pattern, with the highest abundances concentrated in locations with specific environmental characteristics. This aggregation is a result of scale-dependent factors that shape the population trend. In addition, we identified a hierarchical structure of factors affecting the distribution patterns at two different local and regional scales.
Contrary to our initial expectations, we only found limited significant changes in the regional distribution of the species over time. Both the occupancy and abundance distribution patterns, assessed using ROAPs and D* statistics, exhibited a relatively stable trend during the study period. This stability can be attributed to the intrinsic and consistent fidelity of Egyptian vultures to their breeding territories [
60], a characteristic observed in other raptors [
61], which ensures that individuals remain in their territories for many years regardless of environmental changes. Moreover, the combination of territorial fidelity and the conspecific attraction of raptor species [
62], resulting in the selection of territories near successful conspecific settlements (see
habitat-copying hypothesis in [
63]), probably confers great population stability at a regional scale. However, despite this regional stability, we did detect temporal variability expressed as a large number of cells with low abundance values (e.g., with only one breeding pair) with discontinuous occupancy over time (
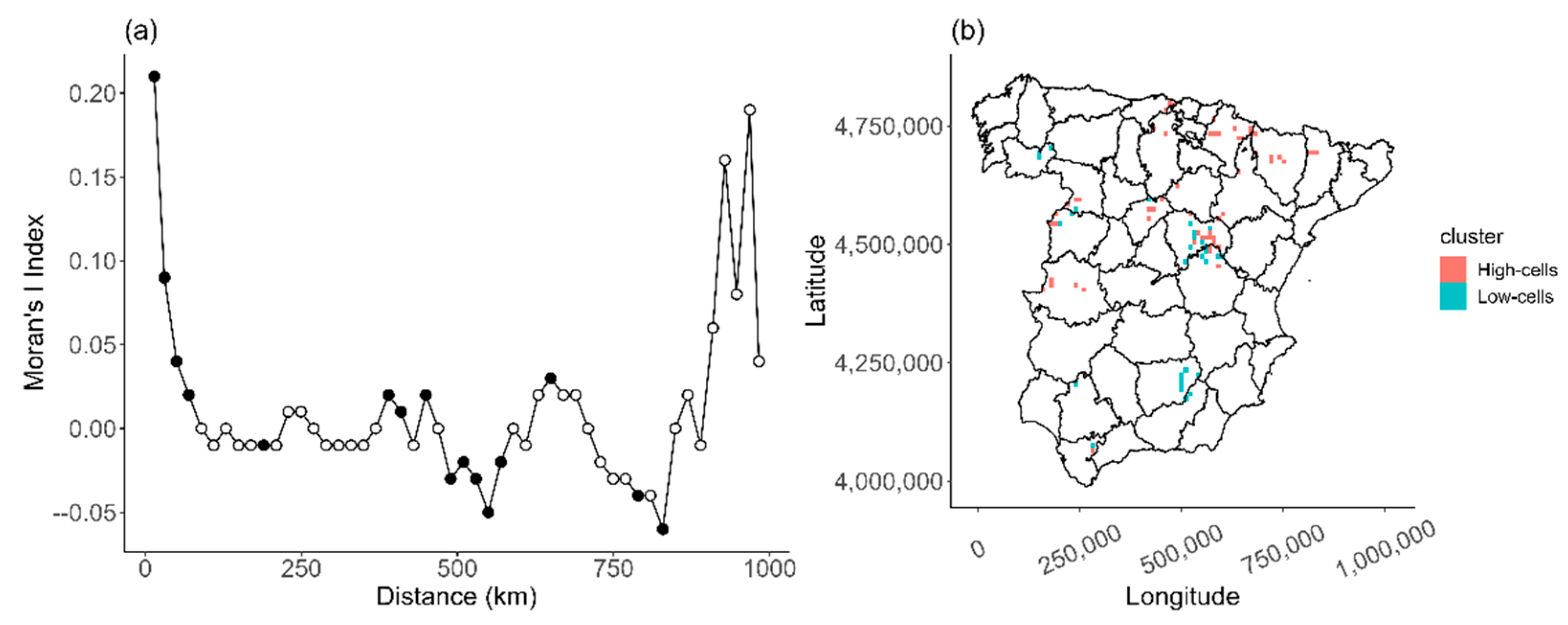
Figure 1). This observed variability in temporal abundance can be explained by human-related factors (e.g., illegal poisoning; [
32,
64]) or by demographic stochasticity (i.e., if there are few individuals, the grid cell is more likely to empty).
Additionally, the shape of the ROAP suggested a spatial aggregation of breeding territories. The steep curves indicated that breeding pairs tend to cluster in specific areas, which was confirmed by the LISA analysis that identified cells with a large number of breeding pairs. We observed more cells with low abundances (one breeding pair per cell) than cells with high abundances (five or more breeding pairs per cell), resulting in a heterogeneous distribution pattern. Moreover, this heterogeneity was also supported by our autocorrelation analyses, which revealed clear spatial autocorrelation in the census data over short distances (i.e., 20 km), consistent with patterns observed in other populations (e.g., in Turkey, [
65]) and other raptors (e.g., lesser kestrel; [
66]). The observed spatial aggregation was found to be a result of scale-dependent factors that shape the abundance distribution. Certain local-level factors such as the presence of wind turbines were associated with lower values of abundance, suggesting they acted as drivers of these patterns. The higher cover of
dehesas and presence of supplementary feeding stations were associated with more breeding territories, which indicates that these factors favor the study species. However, it is worth noting that these factors only act in specific marginal areas and not throughout continental Spain. For instance, an Egyptian vulture population in southern Spain was affected by wind farm-related mortality during the breeding [
34] and migration [
67] periods. Additionally, the
dehesas and agroforestry areas located only in western Spain serve as important foraging habitats for other vulture species due to the higher availability of food compared to other agricultural systems or landscapes [
68,
69] and support a high relative abundance of livestock grazing and other species (e.g., rabbits) that scavenging birds can exploit. The authors of [
70] reported that supplementary feeding stations used as a conservation measure help both the maintenance of the closest breeding territories and breeding success. Nevertheless, these supplementary feeding sites that act as local attractors for high densities of vultures and other scavengers may have detrimental consequences. For instance, supplementary feeding stations can adversely affect the productivity of Pyrenean Bearded vultures (
Gypaetus barbatus) due to the congregation of non-breeding individuals, leading to a decline in the quality of the reproductive habitat [
71].
The main factors associated with changes in abundance at the regional scale over both time and space were griffon vulture and cattle abundances. On the one hand, our results suggest that cattle are one of the main food sources of carrion and feces at local and regional levels for the Egyptian vultures breeding in continental Spain, and play an important role in its distribution [
54,
72]. In addition, the coprophagous behavior of this species also explains its close association with cows. Egyptian vultures consume cow dung to obtain lutein, a yellow carotenoid responsible for its facial coloration [
73] that also plays an important role in its immunological system as an antioxidant [
74]. On the other hand, the positive correlation between breeding Egyptian and griffon vultures suggests a heterospecific interaction between these two species that positively impacts the number of Egyptian vulture breeding pairs. Nevertheless, in other studies, the presence of griffon vultures was not associated with the territory occupancy rate of Egyptian vultures as observed in the Balkan Peninsula [
23]. In addition, both vulture species probably respond in a similar fashion to specific environmental characteristics, which means that the abundance of griffon vultures will be an indicator of the most suitable habitat for breeding pairs of Egyptian vulture [
52,
75]. Due to the spatial overlap between these two species, some authors define this interaction as commensalism [
76] because (i) both species have similar ecological requirements (e.g., they are both cliff-nesting; [
26]) and (ii) given that breeding individuals, regardless of the species, are linked to a breeding area, the abundance of breeding griffon vultures may not only indicate a suitable breeding habitat but also a habitat with food availability [
52,
76].
Despite the fact that our main aim was to assess the likely causes of changes in the abundance distribution of breeding Egyptian vultures at different spatial scales, other factors relating to human pressure that probably also play an important role in their distribution should not be neglected in future research (e.g., electrocution and/or collision against power lines; [
33,
34]). Indeed, our results underscore the importance of considering both temporal and spatial variability during the process of generating distribution models. On the one hand, we used temporal population dynamics (i.e., changes between censuses) to capture how the abundance distribution can enhance the subsequent abundance distribution in such a way that the model revealed the suitability of a breeding territory. On the other hand, we took into account spatial autocorrelation in the modelling process because ignoring spatial constraints can lead to inaccurate conclusions (see [
9]). To fully understand the changes in endangered species distribution, more research is needed using other approaches, such as Bayesian INLA models, that consider the spatiotemporal variation in species abundance [
77].
Conservation Implications
Our findings reveal the scale-dependent factors that influence the Egyptian vulture breeding population in mainland Spain. At the regional level, these factors require the implementation of global conservation strategies to ensure the species is protected across large areas and to serve as guidelines for developing conservation synergies between neighboring areas. At the local scale, the factors affecting populations or even individuals require specific actions related to the main threats affecting each population. Therefore, it is important to highlight the impact of hierarchical approaches on environmental policies. Thus, successful conservation programs aimed at preserving large vertebrate species over large areas should incorporate efficient local management actions [
7,
78]. Based on our results, we advocate the development of a national strategy promoting, at the regional level, extensive livestock farming and the abandoning of healthy carcasses (with sanitary control) as an important and unpredictable food source for not only Egyptian vultures but the whole vulture guild [
79]. Although this approach is partially implemented through the ZPAEN network (Protection Zones for the Feeding of Necrophagous Species of Community Interest), local administrations use different criteria to establish these zones, which leads to a lack of coordination at the regional scale (see [
80]). Additionally, some local actions should be taken to counteract the negative effect of the blades of wind turbines with which certain soaring birds including vultures are prone to collide ([
81,
82], personal data). Some studies have shown that the strategic placement of wind turbines and appropriate mitigation measures could help minimize the potential negative effects of wind farms on soaring birds while still allowing for the generation of renewable energy [
83]. Finally, we believe it is important to underline the importance of grids with a single or few breeding pairs, since the potential for recovery and growth of endangered populations lies in these low-density areas. Conserving small populations allows them to reproduce and expand gradually, and to serve as future sources for repopulating larger areas.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





