


Dietary Polysaccharide-Rich Extract from Noni (Morinda citrifolia L.) Fruit Modified Ruminal Fermentation, Ruminal Bacterial Community and Nutrient Digestion in Cashmere Goats


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Polysaccharide-Rich Fraction of Noni Fruit
2.2. Experiment 1: In Vitro Batch Fermentation
2.2.1. Experimental Design and Diets
2.2.2. Inoculum and Substrates
2.2.3. In Vitro Incubation and Sampling
2.2.4. Calculation
2.3. Experiment 2: Rearing Experiment
2.3.1. Animals, Diets and Experimental Setup
2.3.2. Sampling and Measurements
2.4. Determination of Chemical Composition and Calculation of Apparent Nutrient Digestibility
nutrient content (%) × fecal output (kg)]/diet nutrient content (%) × DMI (kg)
2.5. Determination of Ruminal Fermentation Variables
2.6. DNA Extraction and Sequencing
2.7. Sequence Processing and Data Analysis
2.8. Statistical Analysis
3. Results
3.1. Experiment 1: In Vitro Ruminal Fermentation
3.2. Experiment 2: Ruminal Fermentation
3.3. Growth Performance and Nutrient Digestion
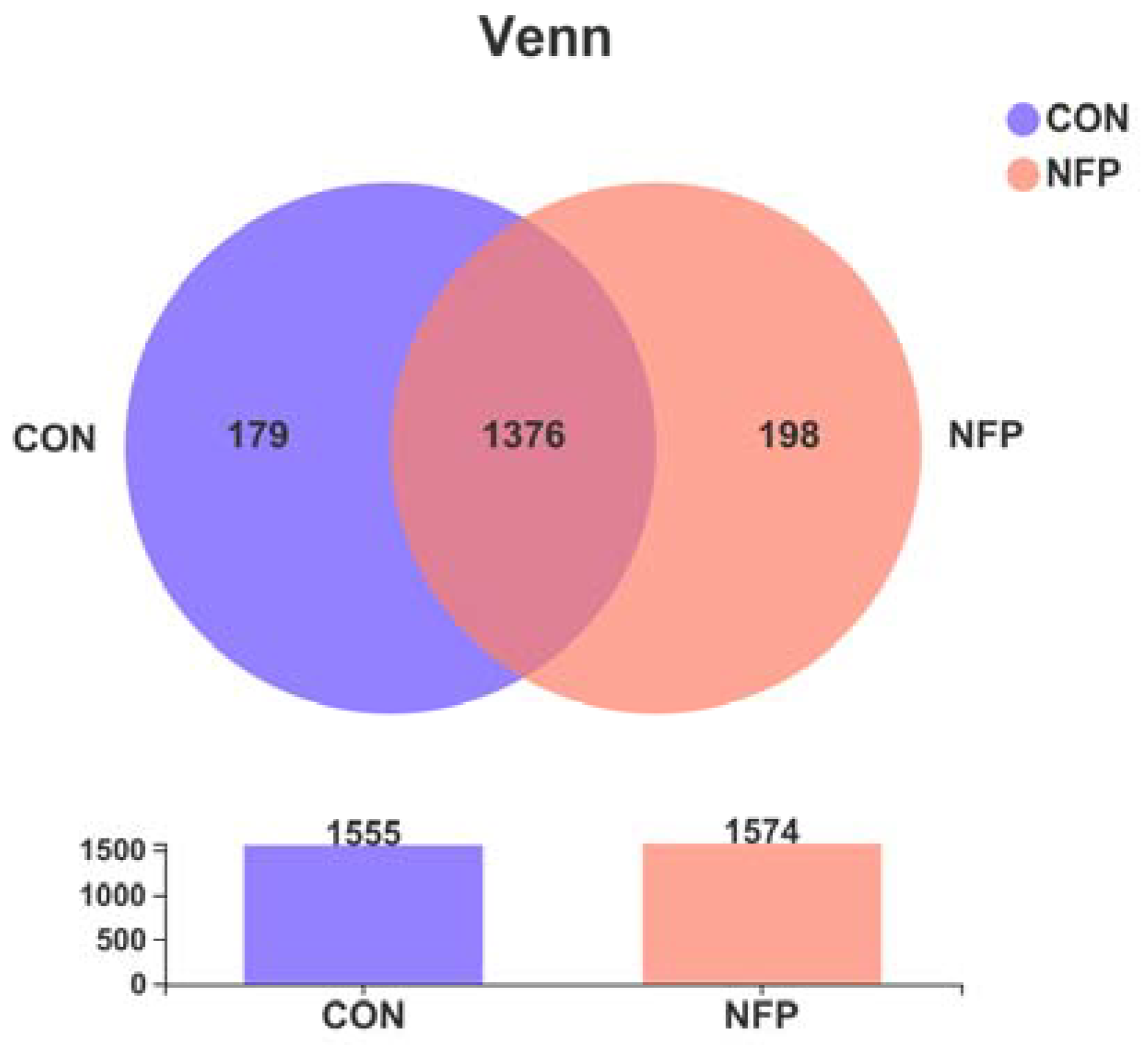
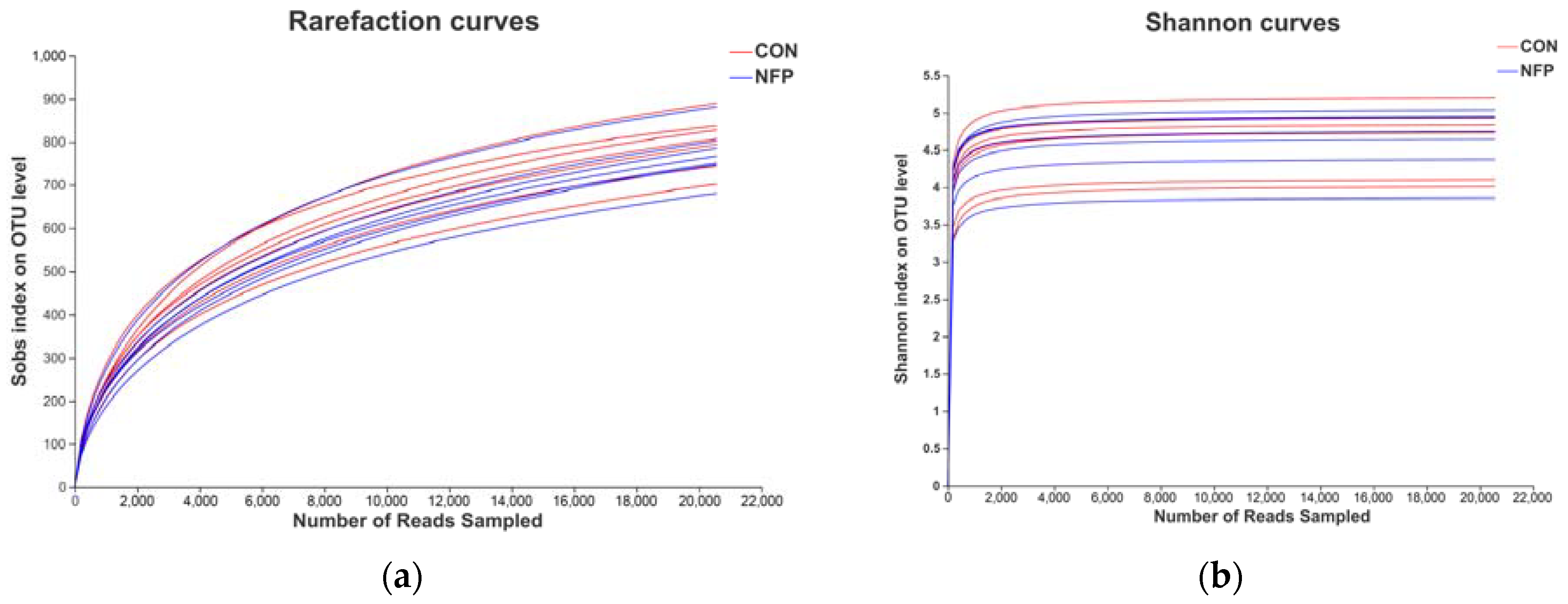
3.4. Sequencing Coverage and Bacterial Diversity
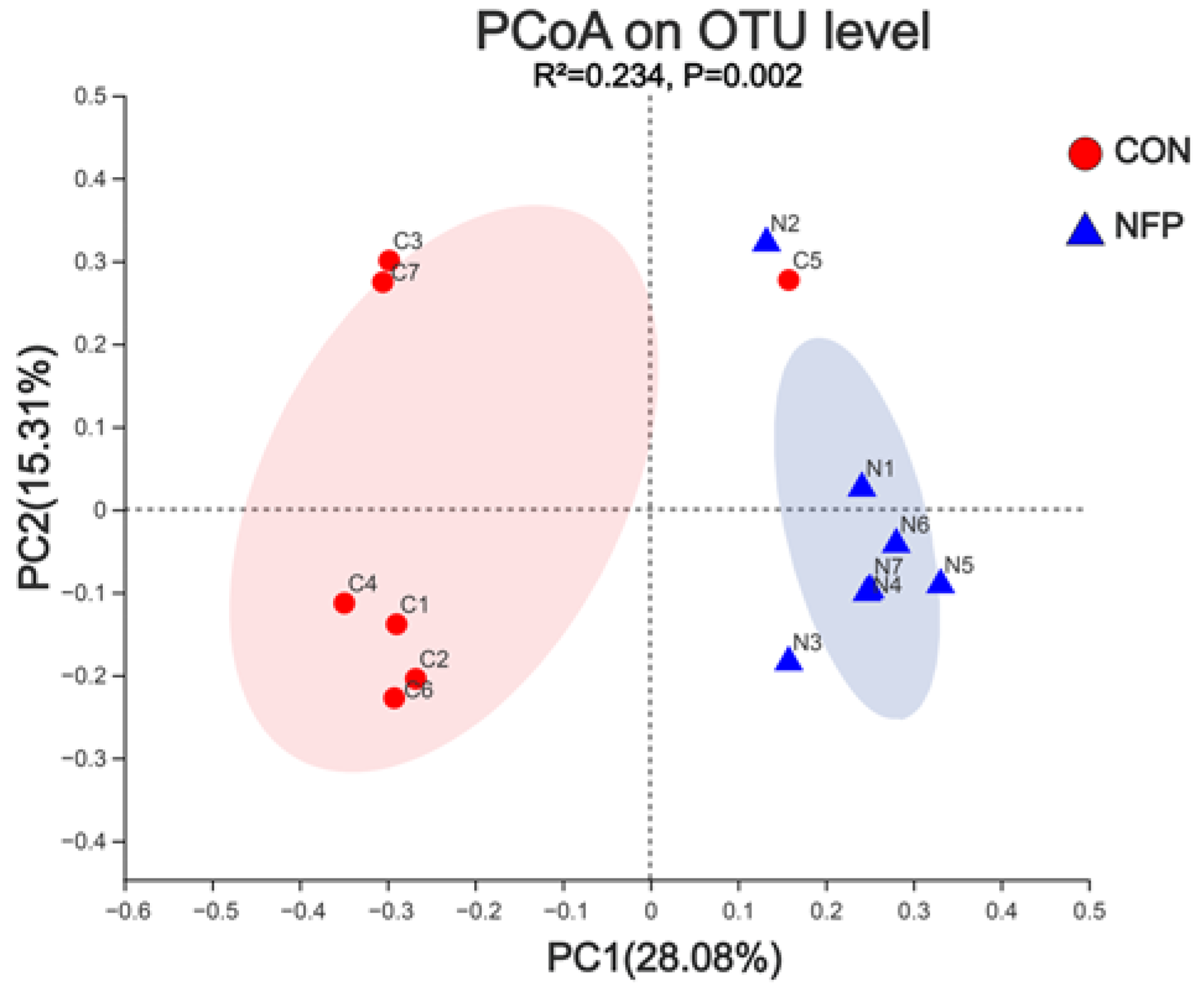
3.5. Principal Coordinate Analysis
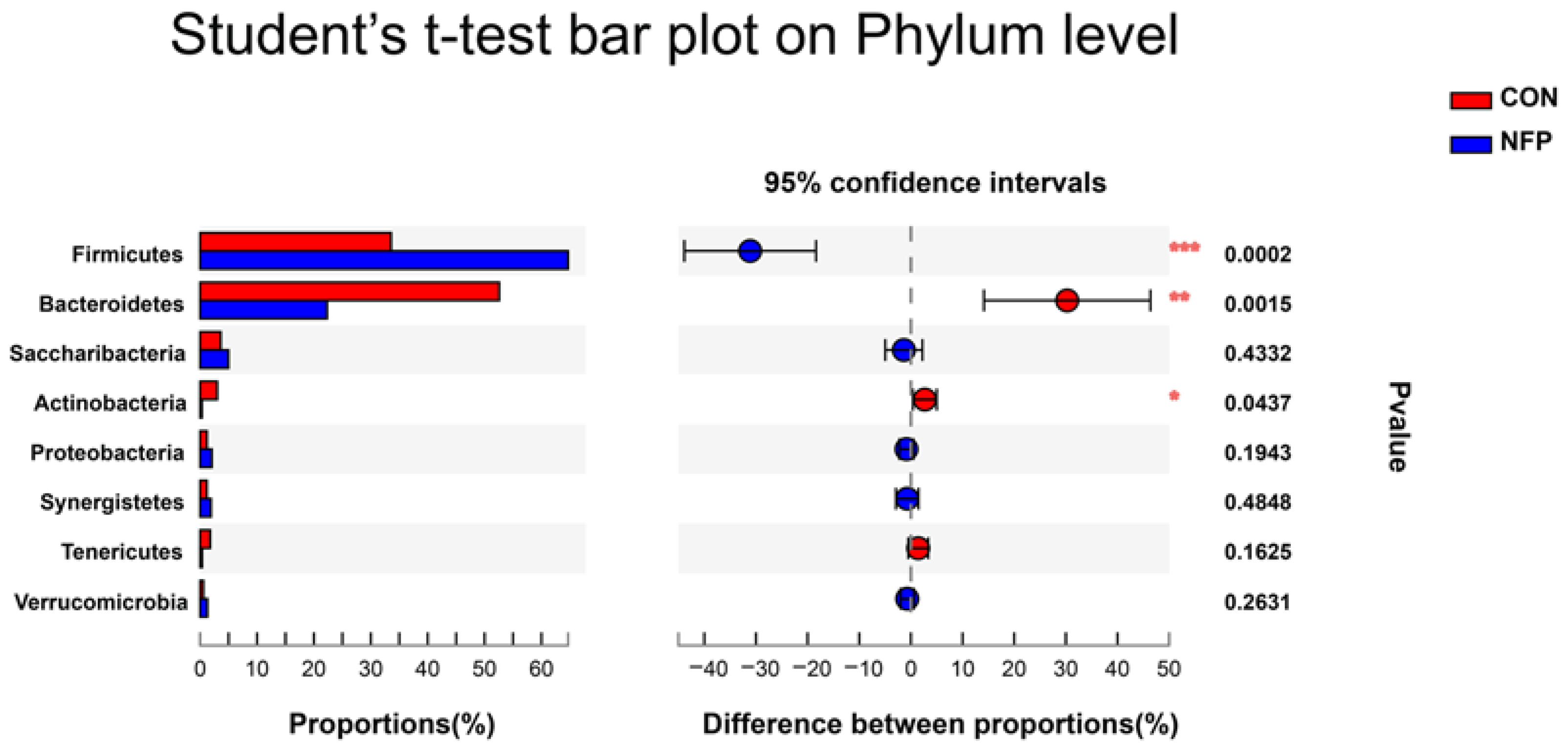
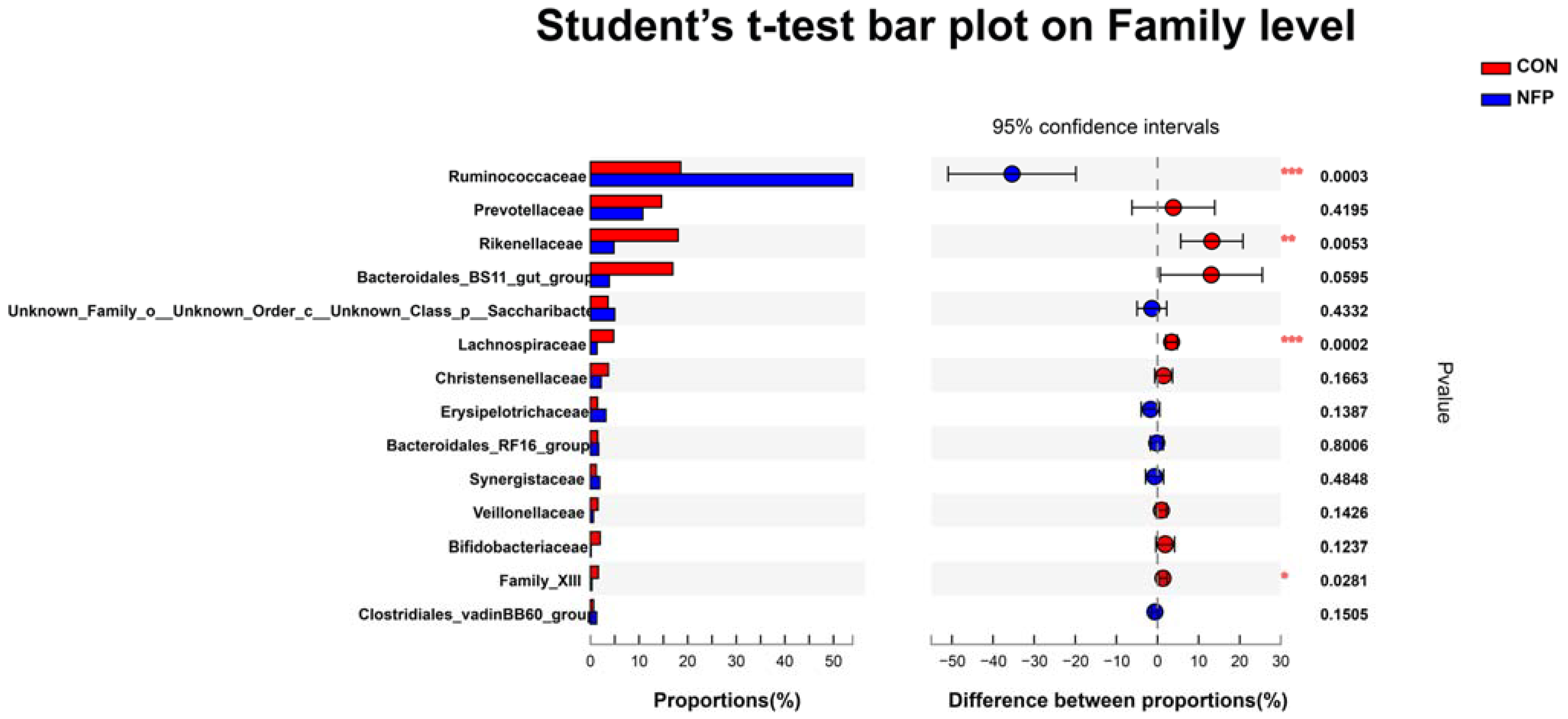
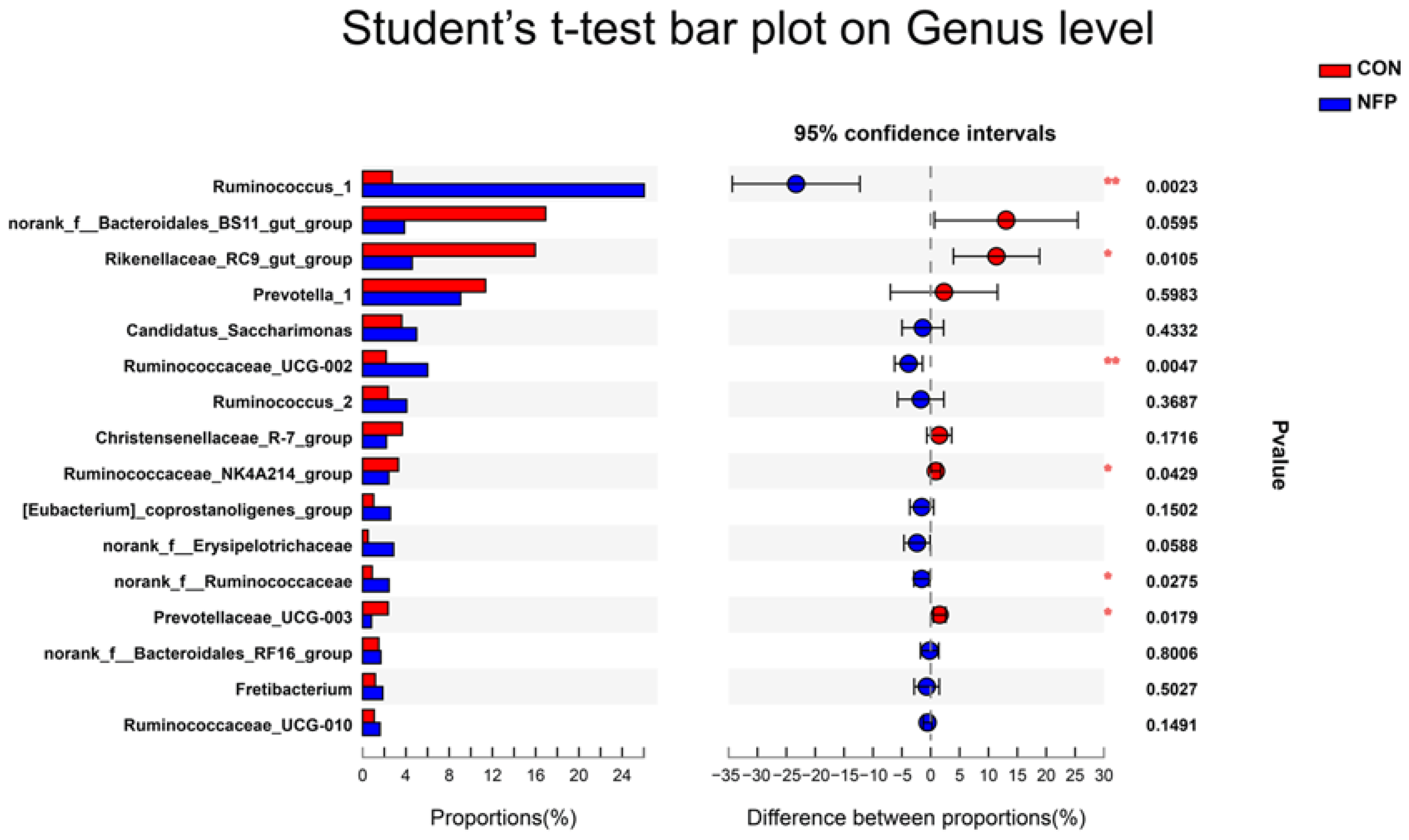
3.6. Microbial Composition Analysis
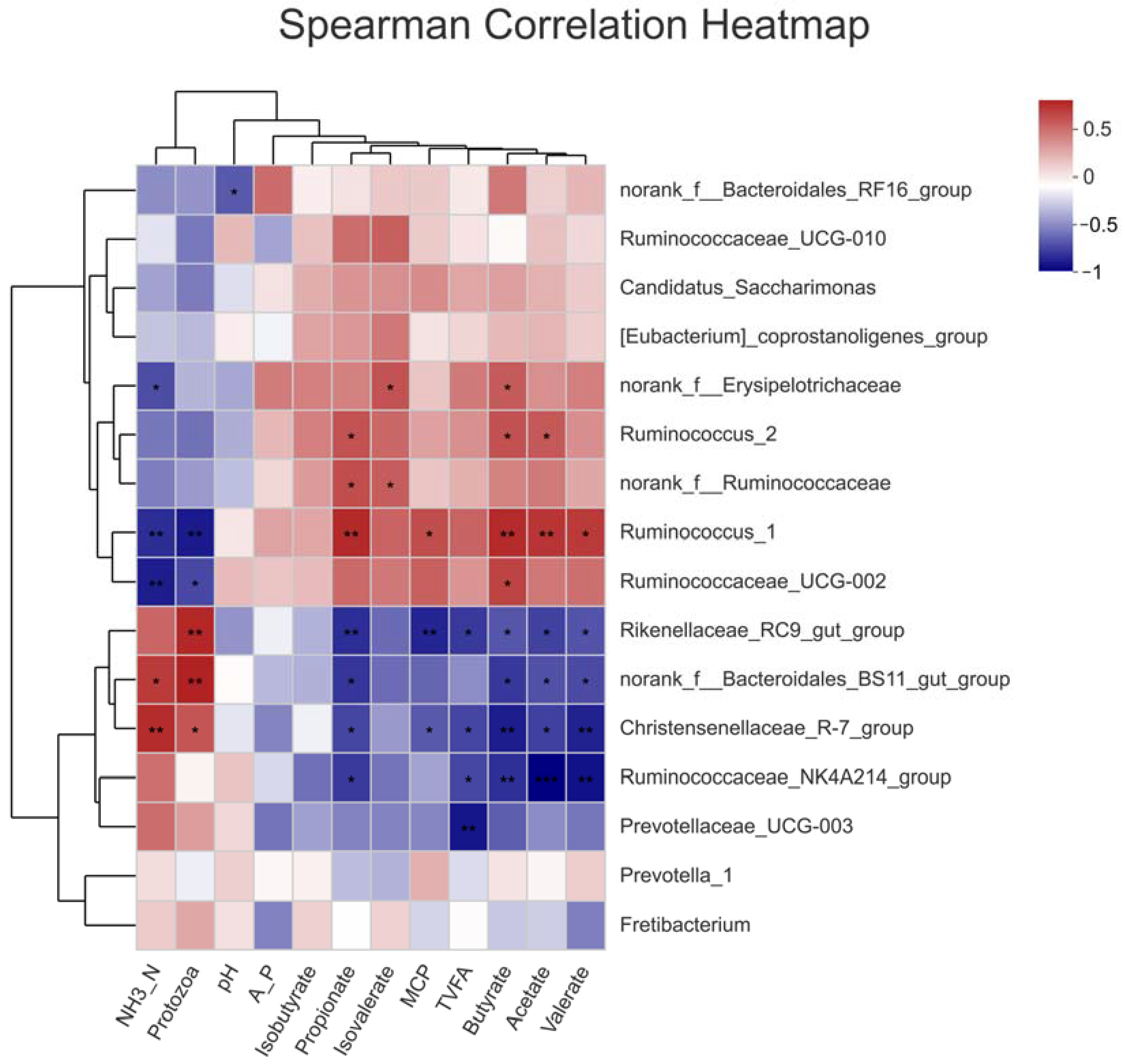
3.7. Spearman Correlation Analysis between Ruminal Bacteria Abundance and Ruminal Fermentation Variables
4. Discussion
4.1. In Vitro Ruminal Fermentation of NFP Diets
4.2. Ruminal Fermentation Variables In Vivo
4.3. Apparent Digestibility of Nutrients, VFA and Ruminal Bacterial Community
4.4. Growth Performance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Q.; Li, Y.; Yin, G.; Li, Y.; Zhao, Y.; Guo, X.; Guo, Y.; Yan, S. The effects of supplementation of noni (Morinda citrifolia L.) fruit polysaccharides-rich extract on antioxidant status and immune function in cashmere goats. J. Anim. Sci. 2022, 23, skac276. [Google Scholar] [CrossRef]
- Wang, X.; Wu, T.; Yan, S.; Shi, B.; Zhang, Y.; Guo, X. Influence of pasture or total mixed ration on fatty acid composition and expression of lipogenic genes of longissimus thoracis and subcutaneous adipose tissues in albas white cashmere goats. Ital. J. Anim. Sci. 2019, 18, 111–123. [Google Scholar] [CrossRef]
- Tiezzi, F.; Tomassone, L.; Mancin, G.; Cornale, P.; Tarantola, M. The assessment of housing conditions, management, animal-based measure of dairy goats′ welfare and its association with productive and reproductive traits. Animals 2019, 9, 893. [Google Scholar] [CrossRef]
- Russell, J.B.; Hespell, R.B. Microbial rumen fermentation. J. Dairy Sci. 1981, 64, 1153–1169. [Google Scholar] [CrossRef]
- Kharaeva, Z.; Shokarova, A.; Shomakhova, Z.; Ibragimova, G.; Trakhtman, P.; Trakhtman, I.; Chung, J.S.; Mayer, W.; De Luca, C.; Korkina, L. Fermented Carica papaya and Morinda citrifolia as perspective food supplements for the treatment of post-COVID symptoms: Randomized placebo-controlled clinical laboratory study. Nutrients 2022, 14, 2203. [Google Scholar] [CrossRef]
- Wang, R.M.; Wang, L.; Zhang, L.; Wan, S.T.; Li, C.F.; Liu, S.X. Solvents effect on phenolics, iridoids, antioxidant activity, antibacterial activity, and pancreatic lipase inhibition activity of noni (Morinda citrifolia L.) fruit extract. Food Chem. 2022, 377, 131989. [Google Scholar] [CrossRef]
- Ma, L.D.; Lin, G.B.; Yang, L.B.; Cao, J.L.; Wang, J.; Chen, Q.D.; Li, W.Q.; Zhong, W.J. Morinda citrifolia (noni) juice suppresses a549 human lung cancer cells via inhibiting akt/nuclear factor-κ b signaling pathway. Chin. J. Integr. Med. 2020, 27, 688–695. [Google Scholar] [CrossRef]
- Ezzat, M.I.; Hassan, M.; Abdelhalim, M.A.; EL-Desoky, A.M.; Mohamed, S.O.; Ezzat, S.M. Immunomodulatory effect of noni fruit and its isolates: Insights into cell-mediated immune response and inhibition of lps-induced thp-1 macrophage inflammation. Food Funct. 2021, 12, 3170–3179. [Google Scholar] [CrossRef]
- Evvyernie, D.; Handayani, M.; Permana, I.G.; Toharmat, T. In vitro fermentability and digestibility of seedless noni waste (Morinda citrifolia L.) as a concentrate substitute in lactating dairy goat diet. IOP Conf. Ser.: Earth Environ. Sci. 2019, 230, 012028. [Google Scholar] [CrossRef]
- Keum, S.H.; Kim, W.S.; Ghassemi Nejad, J.; Lee, J.S.; Jo, Y.H.; Park, K.Y.; Kim, Y.R.; Jo, J.H.; Lee, H.G. Evaluation of the Feed Nutritional Value of Noni (Morinda citrifolia) Meal for Holstein Dairy Cows. Animals 2022, 12, 2196. [Google Scholar] [CrossRef]
- Yancey, J.W.S.; Apple, J.K.; Kegley, E.B.; Godbee, R.G. Effects of Morinda citrifolia (Noni) pulp on growth performance and stress responses of growing cattle. Prof. Anim. Sci. 2013, 29, 420–425. [Google Scholar] [CrossRef]
- Sunder, J.; Jeyakumar, S.; Sujatha, T.; Kundu, A. Grommune: Morinda citrifolia-based herbal tonic for growth and immunity for commercial broilers. J. Appl. Anim. Res. 2015, 43, 137–140. [Google Scholar] [CrossRef]
- Choi, S.I.; La, I.J.; Han, X.; Men, X.; Lee, S.J.; Oh, G.; Kwon, H.Y.; Kim, Y.D.; Seong, G.S.; Kim, S.H.; et al. Immunomodulatory Effect of Polysaccharide from Fermented Morinda citrifolia L. (Noni) on RAW 264.7 Macrophage and Balb/c Mice. Foods 2022, 11, 1925. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.G.; Zhang, B.; Li, L.; Li, X.B.; Zhang, J.Q.; Chen, G.Y. Fermented noni (Morinda citrifolia L.) fruit juice improved oxidative stress and insulin resistance under the synergistic effect of Nrf2/ARE pathway and gut flora in db/db mice and HepG2 cells. Food Funct. 2022, 13, 8254–8273. [Google Scholar] [CrossRef]
- Zhang, B.; Zhou, X.M.; Yu, Z.X.; Li, X.B.; Yi, J.L.; Huang, K.L.; Tang, C.M.; Liu, Q.; Chen, G.Y. Two new phenolic derivatives with α-glucosidase inhibitory activities from Noni Juice. Chinese J. Org. Chem. 2021, 41, 2910–2913. [Google Scholar] [CrossRef]
- Nayak, S.; Mengi., S. Immunostimulant activity of noni (Morinda citrifolia) on T and B lymphocytes. Pharm. Biol. 2010, 48, 724–731. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.Y.; Wu, X.Y.; Li, M.Y.; Li, X.T.; Huang, R.M.; Sun, Y.M.; Xu, Z.L. Noni (Morinda citrifolia L.) fruit polysaccharides regulated ibd mice via targeting gut microbiota: Association of jnk/erk/nf-κb signaling pathways. J. Agric. Food Chem. 2021, 69, 10151–10162. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, E.; Hirazumi, A.; Story, S.; Jensen, J. Antitumour potential of a polysaccharide-rich substance from the fruit juice of Morinda citrifolia (Noni) on sarcoma 180 ascites tumour in mice. Phytother. Res. 2003, 17, 1158–1164. [Google Scholar] [CrossRef]
- Zhong, R.Z.; Yu, M.; Liu, H.W.; Sun, H.X.; Cao, Y.; Zhou, D.W. Effects of dietary Astragalus polysaccharide and Astragalus membranaceus root supplementation on growth performance, rumen fermentation, immune responses, and antioxidant status of lambs. Anim. Feed Sci. Tech. 2012, 174, 60–67. [Google Scholar] [CrossRef]
- Li, Z.J.; Bai, H.X.; Zheng, L.X.; Jiang, H.; Cui, H.Y.; Cao, Y.C.; Yao, J.H. Bioactive polysaccharides and oligosaccharides as possible feed additives to manipulate rumen fermentation in Rusitec fermenters. Int. J. Biol. Macromol. 2018, 109, 1088–1094. [Google Scholar] [CrossRef]
- Kang, J.H.; Zeng, B.; Tang, S.X.; Wang, M.; Han, X.F.; Zhou, C.S.; Yan, Q.X.; Liu, J.F.; Tan, Z.L. Effects of Momordica charantia polysaccharide on in vitro ruminal fermentation and cellulolytic bacteria. Ital. J. Anim. Sci. 2017, 16, 226–233. [Google Scholar] [CrossRef]
- Wei, H.Y.; Ding, L.M.; Wang, X.J.; Yan, Q.; Jiang, C.X.; Hu, C.S.; Wang, G.W.; Zhou, Y.Q.; Henkin, Z.; Degen, A.A. Astragalus root extract improved average daily gain, immunity, antioxidant status and ruminal microbiota of early weaned yak calves. J. Sci. Food Agric. 2021, 101, 82–90. [Google Scholar] [CrossRef]
- Chen, J.; Zhu, X.Q.; Yang, L.; Luo, Y.; Wang, M.Y.; Liu, X.T.; Liang, K.X.; Gu, X.L. Effect of Glycyrrhiza uralensis Fisch polysaccharide on growth performance and immunologic function in mice in ural city, xinjiang. Asian Pac. J. Trop. Med. 2016, 9, 1056–1061. [Google Scholar] [CrossRef] [PubMed]
- MacArthur Clark, J.A.; Sun, D. Guidelines for the ethical review of laboratory animal welfare People′s Republic of China National Standard GB/T 35892-2018. Anim. Model. Exp. Med. 2020, 1, 103–113. [Google Scholar]
- Li, J.; Yang, P.; Yang, Q.H.; Gong, X.W.; Ma, H.C.; Dang, K.; Chen, G.H.; Gao, X.L.; Feng, B.L. Analysis of flavonoid metabolites in buckwheat leaves using UPLC-ESI-MS/MS. Molecules 2019, 24, 1310. [Google Scholar] [CrossRef] [PubMed]
- Menke, K.H.; Raab, L.; Salewski, A.; Steingass, H.; Fritz, D.; Schneider., W. The estimation of the digestibility and metabolizable energy content of ruminant feedingstuffs from the gas production when they are incubated with rumen liquor in vitro. J. Agric. Sci. Camb. 1979, 93, 217–222. [Google Scholar] [CrossRef]
- Paz, H.A.; Anderson, C.L.; Muller, M.J.; Kononoff, P.J.; Fernando, S.C. Rumen bacterial community composition in Holstein and Jersey cows is different under same dietary condition and is not affected by sampling method. Front. Microbiol. 2016, 7, 1206. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 2000. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Erwin, E.S.; Marco, G.J.; Emery, E.M. Volatile fatty acid analyses of blood and rumen fluid by gas chromatography. J. Dairy Sci. 1961, 44, 1768–1771. [Google Scholar] [CrossRef]
- Broderick, G.A.; Kang, J.H. Automated Simultaneous Determination of Ammonia and Total Amino Acids in Ruminal Fluid and In Vitro Media1. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef]
- Makkar, H.P.S.; Sharma, O.P.; Dawra, R.K.; Negi, S.S. Simple determination of microbial protein in rumen liquor. J. Dairy Sci. 1982, 65, 2170–2173. [Google Scholar] [CrossRef] [PubMed]
- Dehority, B.A. Evaluation of subsampling and fixation procedures used for counting rumen protozoa. Appl. Environ. Microb. 1984, 48, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microb. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O′Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-7. Available online: https://CRAN.R-project.org/package=vegan (accessed on 22 December 2022).
- Chen, J.; Bittinger, K.; Charlson, E.S.; Hoffmann, C.; Lewis, J.; Wu, G.D.; Collman, R.G.; Bushman, F.D.; Li, H.Z. Associating microbiome composition with environmental covariates using generalized UniFrac distances. Bioinformatics 2012, 28, 2106–2113. [Google Scholar] [CrossRef]
- Paradis, E.; Schliep, K. Ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 2018, 35, 526–528. [Google Scholar] [CrossRef]
- Firkins, J.L. Maximizing microbial protein synthesis in the rumen. J. Nutr. 1996, 126, 1347S–1354S. [Google Scholar] [CrossRef]
- Rajabi, M.; Rouzbehan, Y.; Rezaei, J.A. strategy to improve nitrogen utilization, reduce environmental impact, and increase performance and antioxidant capacity of fattening lambs using pomegranate peel extract. J. Anim. Sci. 2017, 95, 499–510. [Google Scholar] [CrossRef]
- Jia, P.; Cui, K.; Ma, T.; Wan, F.; Wang, W.Y.; Yang, D.; Wang, Y.F.; Guo, B.L.; Zhao, L.F.; Diao, Q.Y. Influence of dietary supplementation with Bacillus licheniformis and Saccharomyces cerevisiae as alternatives to monensin on growth performance, antioxidant, immunity, ruminal fermentation and microbial diversity of fattening lambs. Sci. Rep. 2018, 8, 16712. [Google Scholar] [CrossRef]
- Williams, A.G. Rumen holotrich ciliate protozoa. Microbiol. Rev. 1986, 50, 25–49. [Google Scholar] [CrossRef]
- Anjani, F.M.; Evvyernie, D.; Permana, I.G. In vitro evaluation of Noni juice extract waste (Morinda citrifolia L.) in lactating dairy goat diet. AIP Conf. Proc. 2019, 2120, 070010. [Google Scholar] [CrossRef]
- Busquet, M.; Calsamiglia, S.; Ferret, A.; Kamel, C. Screening for effects of plant extracts and active compounds of plants on dairy cattle rumen microbial fermentation in a continuous culture system. Anim. Feed Sci. Tech. 2005, 124, 597–613. [Google Scholar] [CrossRef]
- Cone, J.W.; Becker, P.M. Fermentation kinetics and production of volatile fatty acids and microbial protein by starchy feedstuffs. Anim. Feed Sci. Tech. 2012, 172, 34–41. [Google Scholar] [CrossRef]
- Cherdthong, A.; Wanapat, M. In vitro gas production in rumen fluid of buffalo as affected by urea-calcium mixture in high-quality feed block. Anim. Sci. J. 2014, 85, 420–426. [Google Scholar] [CrossRef]
- Kidane, A.; Overland, M.; Mydland, L.T.; Prestlokken, E. Interaction between feed use efficiency and level of dietary crude protein on enteric methane emission and apparent nitrogen use efficiency with Norwegian Red dairy cows. J. Anim. Sci. 2018, 96, 3967–3982. [Google Scholar] [CrossRef] [PubMed]
- Kidane, A.; Vhile, S.G.; Ferneborg, S.; Skeie, S.; Olsen, M.A.; Mydland, L.T.; Overland, M.; Prestlokken, E. Cyberlindnera jadinii yeast as a protein source in early- to mid-lactation dairy cow diets: Effects on feed intake, ruminal fermentation, and milk production. J. Dairy Sci. 2022, 105, 2343–2353. [Google Scholar] [CrossRef]
- Wallace, R.J.; McEwan, N.R.; McIntosh, F.M.; Teferedegne, B.; Newbold, C.J. Natural products as manipulators of rumen fermentation. Asian Austral J. Anim. 2002, 15, 1458–1468. [Google Scholar] [CrossRef]
- Jouany, J.P.; Morgavi, D.R. Use of ‘natural’ products as alternatives to antibiotic feed additives in ruminant production. Animal 2007, 1, 1443–1466. [Google Scholar] [CrossRef]
- Singh, D.R.; Singh, S. Phytochemicals in plant parts of noni (Morinda citrifolia L.) with special reference to fatty acid profiles of seeds. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2013, 83, 471–478. [Google Scholar] [CrossRef]
- Brade, W.; Distl, O. The ruminal microbiome of cattle Part 3: Eukaryotic unicellular organisms-additional organisms of the ruminal microbiome. Ber. Landwirtsch. 2015, 93, 1–12. [Google Scholar]
- Newbold, C.J.; de la Fuente, G.; Belanche, A.; Ramos-Morales, E.; McEwan, N.R. The Role of Ciliate Protozoa in the Rumen. Front. Microbiol. 2015, 6, 1313. [Google Scholar] [CrossRef] [PubMed]
- Boone, D.R.; Castenholz, R.W.; Garrity, G.M. Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Springer: New York, NY, USA, 2001. [Google Scholar] [CrossRef]
- Olson, N.D.; Morrow, J.B. DNA extract characterization process for microbial detection methods development and validation. BMC Res. Notes 2012, 5, 668. [Google Scholar] [CrossRef] [PubMed]
- Li, S.T.; Zhang, C.; Gu, Y.Y.; Chen, L.; Ou, S.Y.; Wang, Y.; Peng, X.C. Lean rats gained more body weight than obese ones from a high-fibre diet. Brit. J. Nutr. 2015, 114, 1188–1194. [Google Scholar] [CrossRef] [PubMed]
- Benchaar, C.; Romero-Perez, G.A.; Chouinard, P.Y.; Hassanat, F.; Eugene, M.; Petit, H.V.; Cortes, C. Supplementation of increasing amounts of linseed oil to dairy cows fed total mixed rations: Effects on digestion, ruminal fermentation characteristics, protozoal populations, and milk fatty acid composition. J. Dairy Sci. 2012, 95, 4578–4590. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Teather, R.; Forster, R. Composition, spatial distribution, and diversity of the bacterial communities in the rumen of cows fed different forages. FEMS Microbiol. Ecol. 2010, 74, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Joch, M.; Mrazek, J.; Skrivanova, E.; Cermak, L.; Marounek, M. Effects of pure plant secondary metabolites on methane production, rumen fermentation and rumen bacteria populations in vitro. J. Anim. Physiol. An. N. 2018, 102, 869–881. [Google Scholar] [CrossRef]
- Biddle, A.; Stewart, L.; Blanchard, J.; Leschine, S. Untangling the genetic basis of fibrolytic specialization by lachnospiraceae and ruminococcaceae in diverse gut communities. Diversity 2013, 5, 627–640. [Google Scholar] [CrossRef]
- Burke, M.V.; Small, D.M. Physiological mechanisms by which non-nutritive sweeteners may impact body weight and metabolism. Physiol. Behav. 2015, 152 (Pt B), 381–388. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Content, % |
|---|---|
| Saccharides | 45.47 |
| Organic acids and their derivatives | 21.37 |
| Lipids | 6.09 |
| Alkaloids and their derivatives | 6.14 |
| Amino acids and their derivatives | 7.06 |
| Coumarins | 1.36 |
| Terpenes | 1.18 |
| Cholines | 1.22 |
| Nucleotides and their derivatives | 1.21 |
| Alcohols | 1.13 |
| Phenols and their derivatives | 1.08 |
| Vitamins and their derivatives | 0.47 |
| Salts | 0.97 |
| Others | 5.27 |
| Item | Content |
|---|---|
| Ingredient, g/kg air dry basis | |
| Millet straw | 589.6 |
| Alfalfa hay | 29.6 |
| Oat grass hay | 80.8 |
| Corn grain | 145.8 |
| Soybean meal | 53.0 |
| Distillers dried grains with solubles | 33.0 |
| Flax cake | 53.0 |
| Limestone meal | 1.2 |
| Calcium bicarbonate | 1.2 |
| Premix 1 | 5.0 |
| Sodium chloride | 3.0 |
| Sodium bicarbonate | 4.8 |
| Nutrient composition | |
| Digestible energy, MJ/kg dry matter basis (DM) 2 | 9.95 |
| DM, %, air dry basis | 92.36 |
| CP, %, DM | 10.04 |
| aNDFom 3, %, DM | 54.71 |
| ADFom 4, %, DM | 30.98 |
| EE, %, DM | 2.24 |
| Calcium, %, DM | 0.63 |
| Phosphorus, %, DM | 0.29 |
| Item | pH | NH3-N, mg/dL | Protozoa, 104/mL | MCP, mg/dL | GP, mL |
|---|---|---|---|---|---|
| Treatment | |||||
| CON | 6.67 a | 10.6 a | 6.11 a | 20.4 | 56.1 ab |
| P0.1 | 6.52 b | 10.1 b | 5.48 ab | 23.3 | 60.3 ab |
| P0.2 | 6.52 b | 9.97 b | 5.27 b | 24.7 | 60.7 a |
| P0.4 | 6.53 b | 9.91 b | 5.03 b | 25.3 | 61.9 a |
| P0.55 | 6.53 b | 10.1 b | 5.25 b | 24.5 | 60.5 a |
| Incubation time, h | |||||
| 3 | 6.78 A | 9.78 C | 0.37 E | 12.5 D | 21.6 E |
| 6 | 6.80 A | 6.99 D | 1.55 D | 15.2 CD | 43.6 D |
| 9 | 6.56 B | 6.96 D | 7.89 B | 17.2 C | 64.6 C |
| 12 | 6.53 B | 15.5 A | 12.6 A | 40.9 A | 76.8 B |
| 24 | 6.10 C | 11.6 B | 4.77 C | 32.4 B | 93.0 A |
| SEM | 0.017 | 0.155 | 0.231 | 1.25 | 1.51 |
| p-value | |||||
| TRT | <0.001 | 0.011 | 0.015 | 0.085 | 0.112 |
| linear | <0.001 | 0.051 | 0.007 | 0.012 | 0.031 |
| quadratic | <0.001 | 0.004 | 0.027 | 0.039 | 0.048 |
| Hour | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| TRT × Hour | 0.015 | 0.029 | 0.700 | 0.314 | 0.112 |
| Item | Acetate, mmol/L | Propionate, mmol/L | Butyrate, mmol/L | Iso-Butyrate, mmol/L | Valerate, mmol/L | Iso-Valerate, mmol/L | TVFA, mmol/L | A/P |
|---|---|---|---|---|---|---|---|---|
| Treatment | ||||||||
| CON | 22.4 b | 8.58 b | 2.71 b | 0.31 b | 0.29 | 0.33 b | 34.6 b | 2.71 a |
| P0.1 | 23.2 ab | 9.06 a | 2.80 ab | 0.32 ab | 0.30 | 0.35 b | 35.8 ab | 2.61 a |
| P0.2 | 23.3 ab | 9.29 a | 2.84 a | 0.32 ab | 0.30 | 0.35 b | 36.2 a | 2.54 ab |
| P0.4 | 24.1 a | 9.27 a | 2.92 a | 0.33 a | 0.30 | 0.37 a | 37.3 a | 2.64 a |
| P0.55 | 23.8 a | 9.41 a | 2.89 a | 0.33 a | 0.31 | 0.36 ab | 37.1 a | 2.59 a |
| Incubation time, h | ||||||||
| 3 | 18.0 D | 5.79 D | 1.05 D | 0.16 C | 0.14 E | 0.12 D | 25.3 D | 3.15 A |
| 6 | 18.4 D | 7.52 C | 2.22 C | 0.16 C | 0.17 D | 0.13 D | 28.6 C | 2.45 C |
| 9 | 28.2 A | 10.3 B | 3.33 B | 0.33 B | 0.33 C | 0.31 C | 42.8 A | 2.75 B |
| 12 | 25.6 C | 10.1 B | 3.44 B | 0.47 A | 0.45 A | 0.58 B | 40.7 B | 2.54 C |
| 24 | 26.6B | 11.9 A | 4.13 A | 0.48 A | 0.43 B | 0.62 A | 43.8 A | 2.20 D |
| SEM | 0.422 | 0.146 | 0.045 | 0.005 | 0.005 | 0.008 | 0.518 | 0.045 |
| p-value | ||||||||
| TRT | 0.049 | 0.001 | 0.013 | 0.004 | 0.159 | 0.027 | 0.018 | 0.103 |
| linear | 0.007 | <0.001 | 0.001 | <0.001 | 0.027 | 0.004 | 0.006 | 0.283 |
| quadratic | 0.181 | 0.047 | 0.122 | 0.642 | 0.677 | 0.312 | 0.091 | 0.120 |
| Hour | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| TRT × Hour | 0.930 | 0.975 | 0.673 | 0.866 | 0.817 | 0.555 | 0.740 | 0.440 |
| Item | −1 h | 6 h | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| CON | NFP | CON | NFP | TRT | Hour | TRT × Hour | ||
| pH | 6.93 | 6.86 | 6.65 | 6.69 | 0.055 | 0.882 | <0.001 | 0.175 |
| NH3-N, mg/dL | 22.9 | 18.8 | 17.8 | 13.9 | 0.905 | 0.017 | 0.003 | 0.929 |
| MCP, mg/dL | 32.1 | 44.5 | 28.3 | 37.0 | 2.405 | 0.009 | 0.044 | 0.477 |
| Protozoa, 104/mL | 34.1 | 29.2 | 53.6 | 36.8 | 2.610 | 0.010 | 0.002 | 0.126 |
| Volatile fatty acids, mmol/L | ||||||||
| Acetate | 21.5 | 25.6 | 26.4 | 32.1 | 1.129 | 0.016 | 0.001 | 0.503 |
| Propionate | 4.13 | 5.01 | 5.44 | 6.68 | 0.184 | 0.003 | <0.001 | 0.392 |
| Butyrate | 3.64 | 5.12 | 5.78 | 6.76 | 0.375 | 0.068 | <0.001 | 0.354 |
| Iso-butyrate | 0.517 | 0.590 | 0.427 | 0.467 | 0.012 | 0.010 | <0.001 | 0.234 |
| Valerate | 0.287 | 0.346 | 0.349 | 0.406 | 0.019 | 0.075 | 0.035 | 0.968 |
| Iso-valerate | 0.674 | 0.758 | 0.427 | 0.484 | 0.018 | 0.028 | <0.001 | 0.480 |
| TVFA | 30.7 | 37.5 | 38.7 | 46.9 | 1.436 | 0.006 | <0.001 | 0.652 |
| Acetate to propionate ratio | 5.24 | 5.09 | 4.77 | 4.81 | 0.200 | 0.875 | 0.022 | 0.506 |
| Item | Treatment | SEM | p-Value | |
|---|---|---|---|---|
| CON | NFP | |||
| Growth performance | ||||
| DMI, kg/d | 1.35 | 1.63 | 0.071 | 0.033 |
| BW beginning of the trial, kg | 46.5 | 46.8 | 1.291 | 0.893 |
| BW end of the trial, kg | 47.5 | 48.6 | 1.277 | 0.579 |
| ADG, kg | 0.054 b | 0.097 a | 0.011 | 0.020 |
| DMI:ADG | 25.0 a | 16.8 ab | 2.770 | 0.065 |
| Apparent nutrient digestibility, % | ||||
| Dry matter | 77.31 a | 78.73 b | 0.301 | 0.008 |
| Crude protein | 81.03 a | 83.84 b | 0.692 | 0.015 |
| Ether extract | 84.99 | 84.11 | 0.579 | 0.308 |
| Neutral detergent fiber | 52.15 | 54.12 | 1.103 | 0.236 |
| Acid detergent fiber | 43.12 | 45.11 | 0.949 | 0.176 |
| Calcium | 38.60 a | 42.93 ab | 1.320 | 0.061 |
| Phosphorus | 43.79 | 48.25 | 4.922 | 0.540 |
| Item | Treatment | SEM | p-Value | |
|---|---|---|---|---|
| CON | NFP | |||
| Clean reads | 44,208 | 43,091 | 2565 | 0.764 |
| Sobs | 800 | 774 | 23.2 | 0.436 |
| Shannon | 4.65 | 4.65 | 0.161 | 0.998 |
| Simpson | 0.047 | 0.039 | 0.016 | 0.725 |
| Ace | 971 | 980 | 21.9 | 0.778 |
| Chao | 981 | 981 | 24.2 | 0.989 |
| Good’s coverage | 0.9903 a | 0.9895 ab | 0.0003 | 0.064 |
| Item | CON | NFP | SEM | p-Value |
|---|---|---|---|---|
| phyla (proportion of total observations) | ||||
| Firmicutes | 0.336 b | 0.647 a | 0.0411 | <0.001 |
| Bacteroidetes | 0.526 a | 0.223 b | 0.0507 | 0.002 |
| Saccharibacteria | 0.036 | 0.050 | 0.0105 | 0.433 |
| Actinobacteria | 0.030 a | 0.003 b | 0.0056 | 0.044 |
| Proteobacteria | 0.012 | 0.021 | 0.0044 | 0.194 |
| Synergistetes | 0.012 | 0.019 | 0.0068 | 0.485 |
| Tenericutes | 0.018 | 0.004 | 0.0049 | 0.163 |
| Verrucomicrobia | 0.006 | 0.013 | 0.0038 | 0.263 |
| family (proportion of total observations) | ||||
| Ruminococcaceae | 0.186 b | 0.539 a | 0.0504 | <0.001 |
| Prevotellaceae | 0.146 | 0.108 | 0.0322 | 0.420 |
| Rikenellaceae | 0.180 a | 0.048 b | 0.0227 | 0.005 |
| Bacteroidales_BS11_gut_group | 0.169 a | 0.039 ab | 0.0327 | 0.060 |
| Unknown_Family_o__Unknown_Order_ c__Unknown_Class_p__Saccharibacteria | 0.036 | 0.050 | 0.0045 | 0.433 |
| Lachnospiraceae | 0.048 a | 0.013 b | 0.0044 | <0.001 |
| Christensenellaceae | 0.037 | 0.022 | 0.0066 | 0.166 |
| Erysipelotrichaceae | 0.015 | 0.032 | 0.0069 | 0.139 |
| Bacteroidales_RF16_group | 0.015 | 0.017 | 0.0031 | 0.801 |
| Synergistaceae | 0.012 | 0.019 | 0.0068 | 0.485 |
| Veillonellaceae | 0.015 | 0.006 | 0.0041 | 0.143 |
| Bifidobacteriaceae | 0.020 | 0.001 | 0.0037 | 0.124 |
| Family_XIII | 0.016 a | 0.003 b | 0.0026 | 0.028 |
| Clostridiales_vadinBB60_group | 0.006 | 0.013 | 0.0026 | 0.151 |
| genera (proportion of total observations) | ||||
| Ruminococcus_1 | 0.027 b | 0.260 a | 0.0319 | 0.002 |
| norank_f__Bacteroidales_BS11_gut_group | 0.169 a | 0.039 ab | 0.0327 | 0.060 |
| Rikenellaceae_RC9_gut_group | 0.160 a | 0.046 b | 0.0224 | 0.011 |
| Prevotella_1 | 0.114 | 0.091 | 0.0291 | 0.598 |
| Candidatus_Saccharimonas | 0.036 | 0.050 | 0.0105 | 0.433 |
| Ruminococcaceae_UCG-002 | 0.022 b | 0.060 a | 0.0074 | 0.005 |
| Ruminococcus_2 | 0.024 | 0.041 | 0.0126 | 0.369 |
| Christensenellaceae_R-7_group | 0.037 | 0.022 | 0.0066 | 0.172 |
| Ruminococcaceae_NK4A214_group | 0.033 a | 0.024 b | 0.0028 | 0.043 |
| [Eubacterium]_coprostanoligenes_group | 0.010 | 0.026 | 0.0056 | 0.150 |
| norank_f__Erysipelotrichaceae | 0.005 ab | 0.029 a | 0.0061 | 0.059 |
| norank_f__Ruminococcaceae | 0.009 b | 0.024 a | 0.0043 | 0.028 |
| Prevotellaceae_UCG-003 | 0.024 a | 0.008 b | 0.0031 | 0.018 |
| Bacteroidales_RF16_group | 0.015 | 0.017 | 0.0048 | 0.801 |
| Fretibacterium | 0.012 | 0.019 | 0.0068 | 0.503 |
| Ruminococcaceae_UCG-010 | 0.011 | 0.016 | 0.0025 | 0.149 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Zhao, Y.; Li, Y.; Guo, X.; Guo, Y.; Ma, G.; Liang, X.; Yan, S. Dietary Polysaccharide-Rich Extract from Noni (Morinda citrifolia L.) Fruit Modified Ruminal Fermentation, Ruminal Bacterial Community and Nutrient Digestion in Cashmere Goats. Animals 2023, 13, 221. https://doi.org/10.3390/ani13020221
Zhang Q, Zhao Y, Li Y, Guo X, Guo Y, Ma G, Liang X, Yan S. Dietary Polysaccharide-Rich Extract from Noni (Morinda citrifolia L.) Fruit Modified Ruminal Fermentation, Ruminal Bacterial Community and Nutrient Digestion in Cashmere Goats. Animals. 2023; 13(2):221. https://doi.org/10.3390/ani13020221
Chicago/Turabian StyleZhang, Qingyue, Yanli Zhao, Yinhao Li, Xiaoyu Guo, Yongmei Guo, Guoqiang Ma, Xiaoshuai Liang, and Sumei Yan. 2023. "Dietary Polysaccharide-Rich Extract from Noni (Morinda citrifolia L.) Fruit Modified Ruminal Fermentation, Ruminal Bacterial Community and Nutrient Digestion in Cashmere Goats" Animals 13, no. 2: 221. https://doi.org/10.3390/ani13020221
APA StyleZhang, Q., Zhao, Y., Li, Y., Guo, X., Guo, Y., Ma, G., Liang, X., & Yan, S. (2023). Dietary Polysaccharide-Rich Extract from Noni (Morinda citrifolia L.) Fruit Modified Ruminal Fermentation, Ruminal Bacterial Community and Nutrient Digestion in Cashmere Goats. Animals, 13(2), 221. https://doi.org/10.3390/ani13020221