



Tongue of the Egyptian Endemic Bridled Skink (Heremites vittatus; Olivier, 1804): Gross, Electron Microscopy, Histochemistry, and Immunohistochemical Analysis
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Sample Collection
2.2. Morphological Examinations
2.3. Scanning Electron Microscopy (SEM) Analysis
2.4. Histological and Mucin Histochemical Examinations
2.5. Immunohistochemical Analysis of the Dorsal Mucosal Lingual Keratin
2.6. Negative Image Analysis Using CMEIAS Color Segmentation (Supplementary Image)
2.7. Digital Coloring of Scanning and Transmission Electron Microscopic Images
3. Results
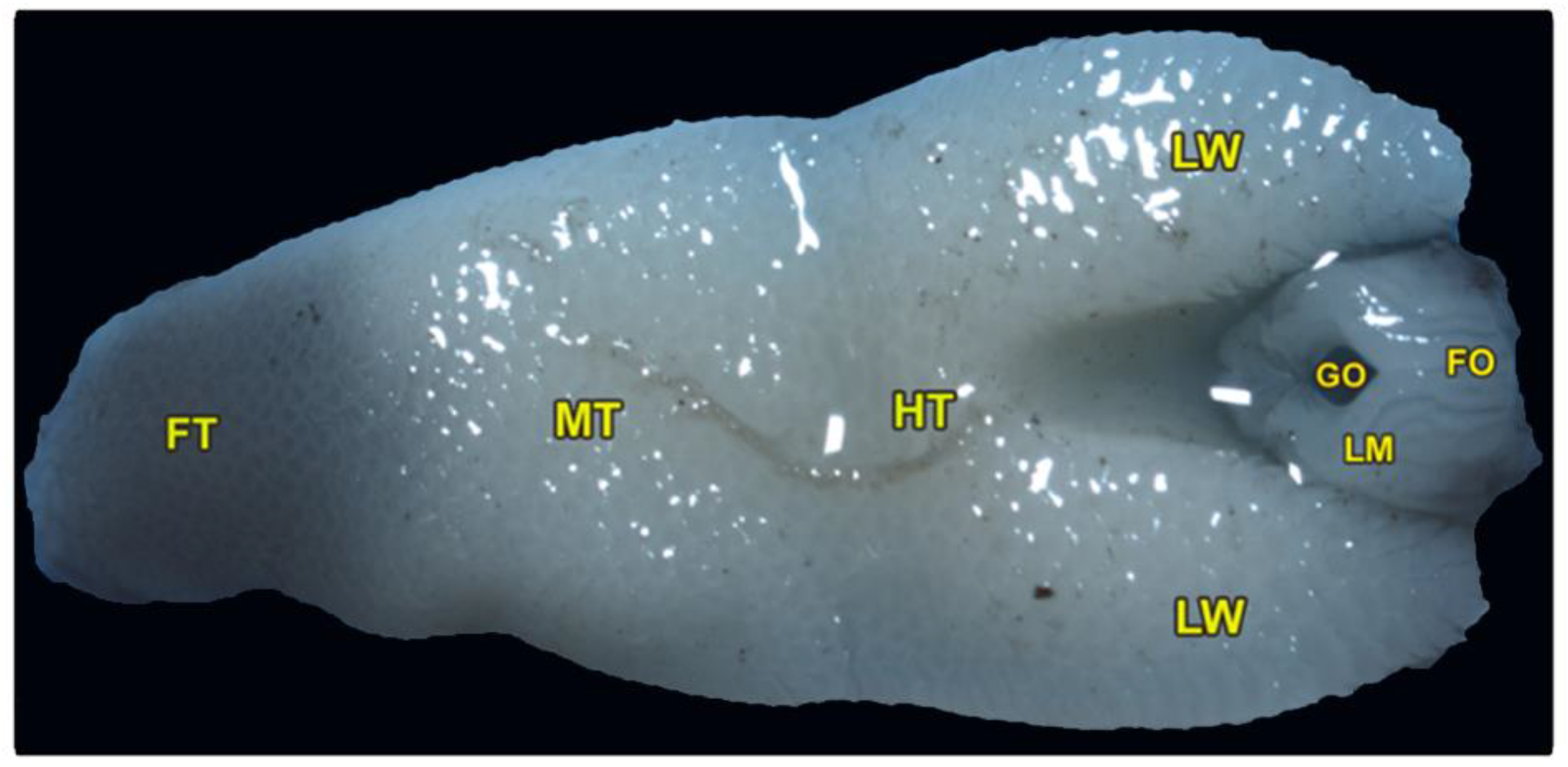
3.1. Gross Morphological Observations of the Tongue and Laryngeal Entrance of the Heremites Vittatus
3.2. Scanning Electron Microscopy Observations of the Tongue and Laryngeal Entrance
3.3. Histological, Histochemical, and Immunohistochemical Studies Examinations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Montuelle, S.J.; Herrel, A.; Libourel, P.A.; Reveret, L.; Bels, V.L. Locomotor–feeding coupling during prey capture in a lizard (Gerrhosaurus major): Effects of prehension mode. J. Exp. Biol. 2009, 6, 768–777. [Google Scholar] [CrossRef]
- Bels, V.; Paindavoine, A.S.; Zghikh, L.; Paulet, E.; Pallandre, J.; Montuelle, S. Feeding in Lizards: Form-Function and Complex Multifunctional System. In Feeding in Vertebrates. Fascinating Life Sciences; Bels, V., Whishaw, I., Eds.; Springer: Cham, Switzerland, 2019; pp. 469–525. [Google Scholar]
- Iwasaki, S. Evolution of the structure and function of the vertebrate tongue. J. Anat. 2002, 201, 1–13. [Google Scholar] [CrossRef]
- Herrel, A.; Canbek, M.; Ozelmas, U.; Uyanoglu, M.; Karakaya, M. Comparative functional analysis of the hyolingual anatomy in lacertid lizards. Anat. Rec. 2005, 284, 561–573. [Google Scholar] [CrossRef]
- Schwenk, K. Of tongues and noses: Chemoreception in lizards and snakes. Trends Ecol. Evol. 1995, 10, 7–12. [Google Scholar] [CrossRef]
- Herrel, A.; Timmermans, J.P.; De Vree, F. Tongue flicking in agamid lizards: Morphology, kinematics, and muscle activity patterns. Anat. Rec. 1998, 252, 102–116. [Google Scholar] [CrossRef]
- El-Mansi, A.A.; Al-Kahtani, M.A.; Abumandour, M.M.A.; Ahmed, A.E. Structural and Functional Characterization of the Tongue and Digestive Tract of Psammophis sibilans (Squamata, Lamprophiidae): Adaptive Strategies for Foraging and Feeding Behaviors. Microsc. Microanal. 2020, 26, 524–541. [Google Scholar] [CrossRef]
- Abumandour, M.M.A.; Farrag, F.A.; El-Mansi, A.; Lashen, S.E.; Shukry, M.; Kassab, M.A.; Hamoda, H.S. Posthatching developmental studies on the tongue and laryngeal entrance of the common quail (Coturnix coturnix, Linnaeus, 1758) in different five age-stages. Microsc. Res. Tech. 2021, 84, 1649–1672. [Google Scholar] [CrossRef]
- Mohammed, M.B.H. Structure and function of the tongue and hyoid apparatus in Acanthodactylus boskianus (Lacertidae; Reptilia). J. Egypt. Ger. Soc. Zool. 1992, 7, 65–89. [Google Scholar]
- Abbate, F.; Guerrera, M.C.; Montalbano, G.; Zichichi, R.; Germana, A.; Ciriaco, E. Morphology of the lingual dorsal surface and oral taste buds in Italian lizard (Podarcis sicula). Anat. Histol. Embryol. 2010, 39, 167–171. [Google Scholar] [CrossRef]
- Bayoumi, S.S.; Abd-Elhameed, A.A.; Mohamed, E.M. Comparative studies on the dorsal lingual surface of two Egyptian squamate reptiles with two different feeding habits. Egypt. J. Exp. Biol. (Zool.) 2011, 7, 203–211. [Google Scholar]
- Abumandour, M.M.A.; Madkour, N.; Morsy, K.; Haddad, S.; Abourashed, N.M.; El Basyouny, H.A.; Kandyel, R.M. Lingual papillary system of the Egyptian mouse-tailed bat (Rhinopoma hardwickii): New insights to its feeding strategies using scanning electron microscope, and immunohistochemical analysis of keratin. Microsc. Res. Tech. 2022, 85, 2192–2205. [Google Scholar] [CrossRef]
- Bahmani, Z.; Rastegar-Pouyani, N.; Rastegar-Pouyani, E. Systematics of the Genus Trachylepis Fitzinger, 1843 (Sauria: Scincidae) in Iran: A review. Iran. J. Anim. Biosyst. 2016, 12, 211–224. [Google Scholar]
- Verderame, M.; Chianese, T.; Rosati, L.; Scudiero, R. Molecular and Histological Effects of Glyphosate on Testicular Tissue of the Lizard Podarcis siculus. Int. J. Mol. Sci. 2022, 23, 4850. [Google Scholar] [CrossRef]
- International Committee on Veterinary Gross Anatomical Nomenclature. Nomina Anatomica Veterinaria, 6th ed.; Revised Version; Editorial Committee: Hanover, Germany; Ghent, Belgium; Sapporo, Japan; Columbia, MO, USA; Rio de Janeiro, Brazil, 2017; Available online: https://www.vetmed.uni-leipzig.de/fileadmin/Fakult%C3%A4t_VMF/Institut_Veterin%C3%A4r-Anatomisches/Dokumente/NAV_6th-Edition-2017.pdf (accessed on 20 July 2023).
- Abumandour, M.M.; Kandyel, R.M. Age-related ultrastructural features of the tongue of the rock pigeon Columba livia dakhlae in different three age stages (young, mature, and adult) captured from Egypt. Microsc. Res. Tech. 2020, 83, 118–132. [Google Scholar] [CrossRef]
- Suvarna, K.S.; Layton, C.; Bancroft, J.D. Bancroft’s Theory and Practice of Histological Techniques, 8th ed.; Elsevier Health Sciences: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Bancroft, J.D.; Cook, H.; Turner, D. Manual of Histological Techniques and Their Diagnostic Application, 2nd ed.; Churchill Livingstone: London, UK, 1996. [Google Scholar]
- Schumacher, U.; Duku, M.; Katoh, M.; Jörns, J.; Krause, W.J. Histochemical similarities of mucins produced by Brunner’s glands and pyloric glands: A comparative study. Anat. Rec. Part A Discov. Mol. Cell. Evol. Biol. Off. Publ. Am. Assoc. Anat. 2004, 278, 540–550. [Google Scholar] [CrossRef]
- Masson, P. Some histological methods: Trichrome staining and their preliminary technique. J. Tech. Methods Bull. Int. Assoc. Med. Mus. 1929, 12, 75–90. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Kandyel, R.M.; Elwan, M.M.; Abumandour, M.M.; El Nahass, E.E. Comparative ultrastructural-functional characterizations of the skin in three reptile species; Chalcides ocellatus, Uromastyx aegyptia aegyptia, and Psammophis schokari aegyptia (Forskal, 1775): Adaptive strategies to their habitat. Microsc. Res. Tech. 2021, 84, 2104–2118. [Google Scholar] [CrossRef]
- Abd-Elkareem, M.; Abou Khalil, N.S.; Sayed, A.E.-D.H. Cytoprotective effect of Nigella sativa seed on 4-nonylphenol-induced renal damage in the African catfish (Clarias gariepinus). Chemosphere 2020, 259, 127379. [Google Scholar] [CrossRef]
- Kandyel, R.M.; El Basyouny, H.A.; El Nahas, E.E.; Madkour, F.; Haddad, S.; Massoud, D.; Morsy, K.; Madkour, N.; Abumandour, M.M.A. A histological and immunohistochemical study on the parabronchial epithelium of the domestic fowl’s (Gallus gallus domesticus) lung with special reference to its scanning and transmission electron microscopic characteristics. Microsc. Res. Tech. 2022, 85, 1108–1119. [Google Scholar] [CrossRef]
- Gewily, D.I.; Mahmoud, F.A.; Saber, S.A.; ElSalkh, B.A.; El-Dahshan, A.A.; Abumandour, M.M.A.; Kandyel, R.M.; Gadel-Rab, A.G. Ultrastructural comparison between the tongue of two reptilian species endemic in Egyptian fauna; Bosc’s fringe-toed lizard Acanthodactylus boskianus and Sinai fan-fingered gecko Ptyodactylus guttatus. Microsc. Res. Tech. 2021, 84, 1977–1991. [Google Scholar] [CrossRef]
- Kandyel, R.M.; Abumandour, M.M.A.; Mahmoud, S.F.; Shukry, M.; Madkour, N.F.; El-Mansi, A.; Farrag, F.A. Structural and Functional adaptation of the lingual papillae of the Egyptian fruit bat (Rousettus aegyptiacus): Specific Adaptive feeding Strategies. Folia Morphol. 2021, 81, 400–411. [Google Scholar] [CrossRef]
- Abbate, F.; Guerrera, M.C.; Levanti, M.; Laurà, R.; Montalbano, G.; Cavallaro, M.; Germanà, A. The tongue of Leopard Gecko (Eublepharis macularius): LM, SEM and confocal laser study. Anat. Histol. Embryol. 2020, 49, 51–59. [Google Scholar] [CrossRef]
- Cizek, P.; Hamouzova, P.; Kvapil, P.; Kyllar, M. Light and scanning electron microscopy of the tongue of the sand lizard (Lacerta agilis). Folia Morphol. 2019, 78, 101–106. [Google Scholar] [CrossRef]
- Abbate, F.; Latella, G.; Montalbano, G.; Guerrera, M.C.; Germana, G.P.; Levanti, M.B. The lingual dorsal surface of the blue-tongue skink (Tiliqua scincoides). Anat. Histol. Embryol. 2009, 38, 348–350. [Google Scholar] [CrossRef]
- Abbate, F.; Latella, G.; Montalbano, G.; Guerrera, M.C.; Levanti, M.B.; Ciriaco, E. Scanning electron microscopical study of the lingual epithelium of green iguana (Iguana iguana). Anat. Histol. Embryol. 2008, 37, 314–316. [Google Scholar] [CrossRef]
- Delheusy, V.; Toubeau, G.; Bels, V.L. Tongue structure and function in Oplurus cuvieri (Reptilia: Iguanidae). Anat. Rec. 1994, 238, 263–276. [Google Scholar] [CrossRef]
- Rabinowitz, T.; Tandler, B. Ultrastructure of lingual salivary glands in the American chameleon: Anoliscaro linensis. Anat. Rec. 1991, 229, 489–494. [Google Scholar] [CrossRef]
- Schaerlaeken, V.; Meyers, J.J.; Herrel, A. Modulation of prey capture kinematics and the role of lingual sensory feedback in the lizard Pogonavitticeps. Zoology 2007, 110, 127–138. [Google Scholar] [CrossRef]
- Iwasaki, S. Fine structure of the dorsal lingual epithelium of the lizard, Gekko japonicus (Lacertilia, Gekkonidae). Am. J. Anat. 1990, 187, 12–20. [Google Scholar] [CrossRef]
- Cooper, W.E. Chemical discrimination by tongue-flicking in lizards: A review with hypotheses on its origin and its ecological and phylogenetic relationships. J. Chem. Ecol. 1994, 20, 439–487. [Google Scholar] [CrossRef]
- Cooper, W.E. Prey chemical discrimination indicated by tongue-flicking in the eublepharid gecko Coleonyx variegates. J. Exp. Zool. 1998, 281, 21–25. [Google Scholar] [CrossRef]
- Baeckens, S.; Herrel, A.; Broeckhoven, C.; Vasilopoulou-Kampitsi, M.; Huyghe, K.; Goyens, J.; Van Damme, R. Evolutionary morphology of the lizard chemosensory system. Sci. Rep. 2017, 7, 101–141. [Google Scholar] [CrossRef]
- Filoramo, N.I.; Schwenk, K. The mechanism of chemical delivery to the vomeronasal organs in squamate reptiles: A comparative morphological approach. J. Exp. Zool. 2009, 34, 20–34. [Google Scholar] [CrossRef]
- Winokur, R.M. The buccopharyngeal mucosa of the turtles (Testudines). J. Morphol. 1988, 196, 33–52. [Google Scholar] [CrossRef]
- Jamniczky, H.A.; Russell, A.P.; Johnson, M.K.; Montuelle, S.J.; Bels, V.L. Morphology and histology of the tongue and oral chamber of Eublepharis macularius (Squamata: Gekkonidae), with special reference to the foretongue and its role in fluid uptake and transport. Evol. Biol. 2009, 36, 397–406. [Google Scholar] [CrossRef]
- Taha, A.M. Comparative Anatomical, Histological and Histochemistrical study of tongue in two species of insectivorous vertebrates. Aust. J. Basic Appl. Sci. 2013, 7, 401–410. [Google Scholar]
- Wassif, E.T. The fine structure of the dorsal lingual epithelium of the scincine lizard Chalcideso cellatus Forscal (Scincidae, Sauria, Reptilia) I. Histogenesis of the lingual epithelium. Egypt. J. Biol. 2001, 3, 12–19. [Google Scholar]
- Yang, C.; Wang, L. Histological and morphological observations on tongue of Scincella tsinlingensis (Reptilia, Squamata, Scincidae). Micron 2016, 80, 24–33. [Google Scholar] [CrossRef]
- Iwasaki, S.; Kobayashi, K. Fine structure of the dorsal lingual epithelium of the Japanese lizard, Takydromus takydromoides. Acta Anat. 1992, 67, 214–225. [Google Scholar]
- Wassif, E.T. Ultrastructure of the lingual epithelium of adult scincine lizard Chalcides ocellatus. Egypt. J. Biol. 2002, 4, 76–86. [Google Scholar]
- Wassif, E.T.; El-Hawary, M.S. Scanning Electron Microscopy of the Dorsal Lingual Epithelium of the Golden Lizard; Eumeces schneideri (Reptilia: Scincidae). J. Egypt. Ger. Soc. Zool. 1998, 26, 11–30. [Google Scholar]
- Zghikh, L.N.; Vangysel, E.; Nonclercq, D.; Legrand, A.; Blairon, B.; Berri, C.; Bordeau, T.; Rémy, C.; Burtéa, C.; Montuelle, S.J. Morphology and fibre-type distribution in the tongue of the Pogona vitticeps lizard (Iguania, Agamidae). J. Anat. 2014, 225, 377–389. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kandyel, R.M.; Choudhary, O.P.; El-Nagar, S.H.; Miles, D.B.; Abumandour, M. Tongue of the Egyptian Endemic Bridled Skink (Heremites vittatus; Olivier, 1804): Gross, Electron Microscopy, Histochemistry, and Immunohistochemical Analysis. Animals 2023, 13, 3336. https://doi.org/10.3390/ani13213336
Kandyel RM, Choudhary OP, El-Nagar SH, Miles DB, Abumandour M. Tongue of the Egyptian Endemic Bridled Skink (Heremites vittatus; Olivier, 1804): Gross, Electron Microscopy, Histochemistry, and Immunohistochemical Analysis. Animals. 2023; 13(21):3336. https://doi.org/10.3390/ani13213336
Chicago/Turabian StyleKandyel, Ramadan M., Om Prakash Choudhary, Sahar H. El-Nagar, Donald B. Miles, and Mohamed Abumandour. 2023. "Tongue of the Egyptian Endemic Bridled Skink (Heremites vittatus; Olivier, 1804): Gross, Electron Microscopy, Histochemistry, and Immunohistochemical Analysis" Animals 13, no. 21: 3336. https://doi.org/10.3390/ani13213336
APA StyleKandyel, R. M., Choudhary, O. P., El-Nagar, S. H., Miles, D. B., & Abumandour, M. (2023). Tongue of the Egyptian Endemic Bridled Skink (Heremites vittatus; Olivier, 1804): Gross, Electron Microscopy, Histochemistry, and Immunohistochemical Analysis. Animals, 13(21), 3336. https://doi.org/10.3390/ani13213336