BLV-miR-B1-5p Promotes Staphylococcus aureus Adhesion to Mammary Epithelial Cells by Targeting MUC1

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Transfection (MAC-T)
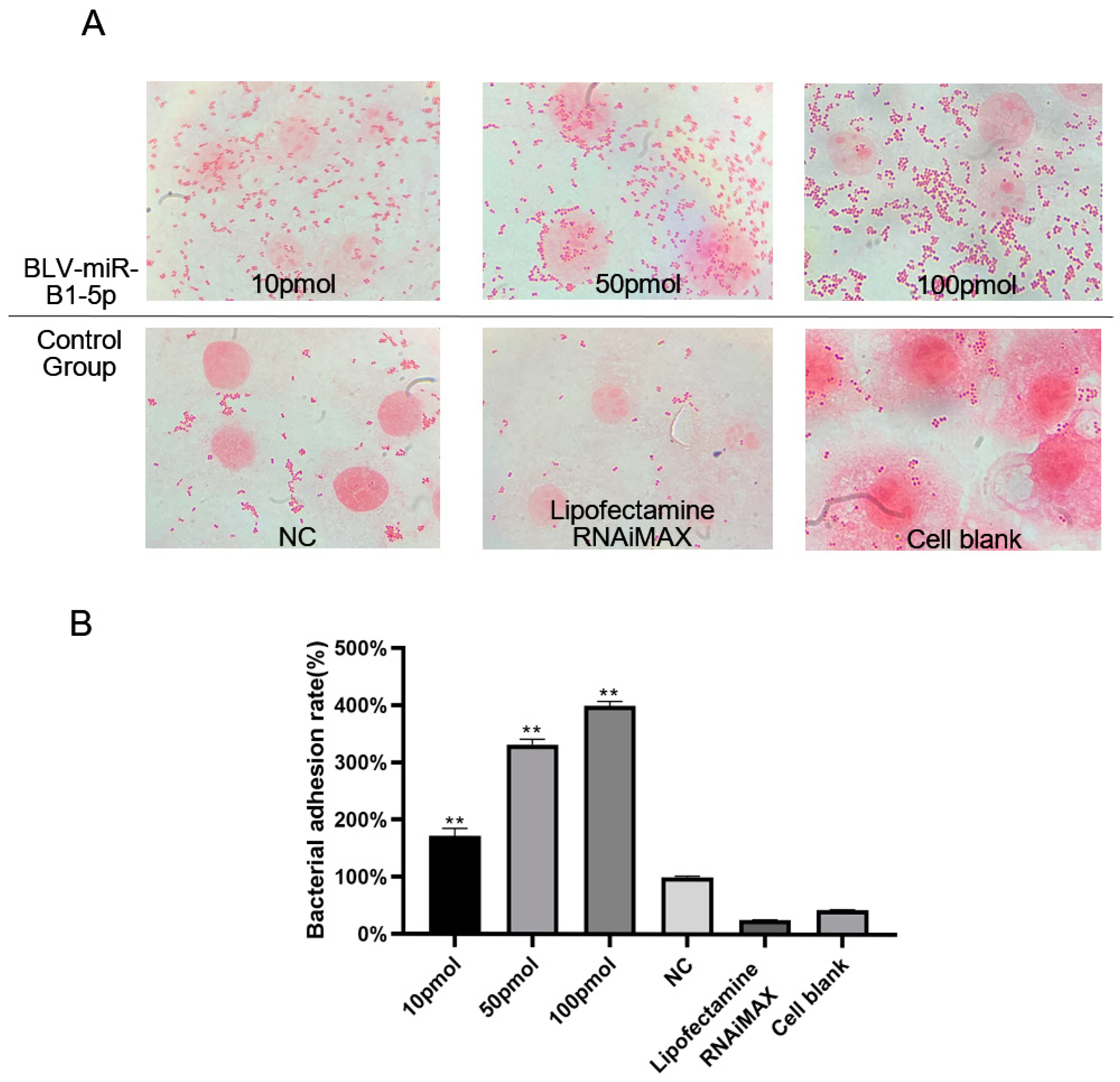
2.3. Bacterial Adhesion
2.4. Target Gene Binding Site Prediction
2.5. Luciferase Reporter Gene Assay
2.6. Analysis of Relative Gene Expression
2.7. Immunofluorescence of MAC-T Cells
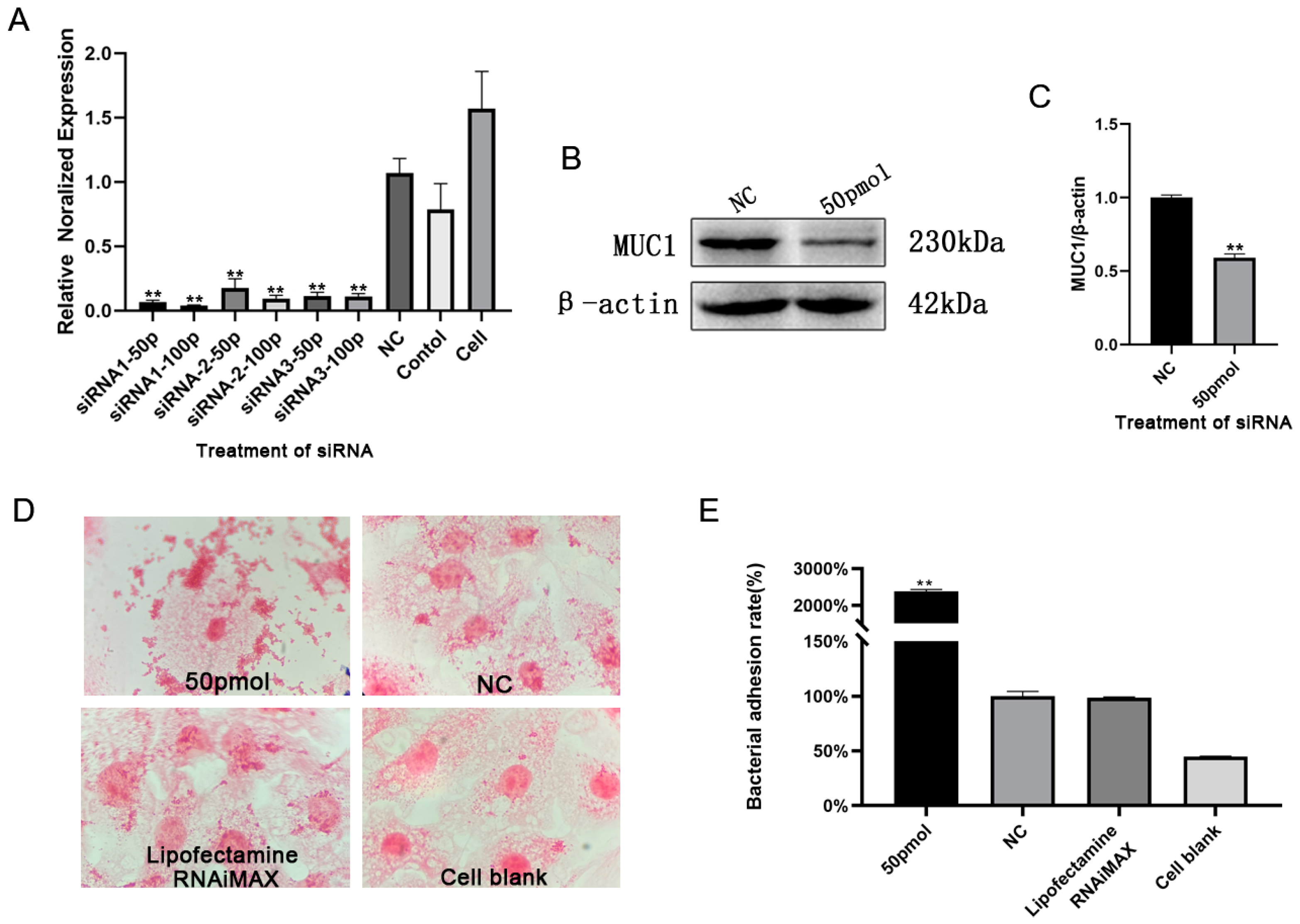
2.8. MUC1 Interference
2.9. Statistical Analysis
3. Results
3.1. BLV-miR-B1-5p Promoted S. aureus Adhesion to Mammary Epithelial Cells
3.2. MUC1 as a Target of BLV-miR-B1-5p
3.2.1. Target Site Prediction
3.2.2. Dual Luciferase Assay
3.3. BLV-miR-B1-5p Repressed MUC1 Gene Expression
3.4. BLV-miR-B1-5p Repressed MUC1 Protein Expression
3.5. Silencing of MUC1 Promoted S. aureus Adhesion to Mammary Epithelial Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Appendix A

References
- Benitez, O.J.; LaDronka, R.M.; Norby, B.; Grooms, D.L.; Bartlett, P.C. The effect of bovine leukemia virus on dairy cow longevity. JDS Commun. 2022, 3, 185–188. [Google Scholar] [CrossRef]
- Kuczewski, A.; Orsel, K.; Barkema, H.W.; Mason, S.; Erskine, R.; van der Meer, F. Invited review: Bovine leukemia virus-Transmission, control, and eradication. J. Dairy Sci. 2021, 104, 6358–6375. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Fan, W.; Mao, Y.; Yang, Z.; Lu, G.; Zhang, R.; Zhang, H.; Szeto, C.; Wang, C. Bovine leukemia virus infection in cattle of China: Association with reduced milk production and increased somatic cell score. J. Dairy Sci. 2016, 99, 3688–3697. [Google Scholar] [CrossRef] [PubMed]
- Blagitz, M.G.; Souza, F.N.; Batista, C.F.; Azevedo, L.F.F.; Sanchez, E.M.R.; Diniz, S.A.; Silva, M.X.; Haddad, J.P.; Della Libera, A. Immunological implications of bovine leukemia virus infection. Res. Vet. Sci. 2017, 114, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Lima, E.S.; Blagitz, M.G.; Batista, C.F.; Alves, A.J.; Fernandes, A.C.C.; Ramos Sanchez, E.M.; Frias Torres, H.; Diniz, S.A.; Silva, M.X.; Della Libera, A.; et al. Milk Macrophage Function in Bovine Leukemia Virus-Infected Dairy Cows. Front. Vet. Sci. 2021, 8, 650021. [Google Scholar] [CrossRef] [PubMed]
- Della Libera, A.M.; de Souza, F.N.; Batista, C.F.; Santos, B.P.; de Azevedo, L.F.; Sanchez, E.M.; Diniz, S.A.; Silva, M.X.; Haddad, J.P.; Blagitz, M.G. Effects of bovine leukemia virus infection on milk neutrophil function and the milk lymphocyte profile. Vet. Res. 2015, 46, 2. [Google Scholar] [CrossRef]
- Frie, M.C.; Sporer, K.R.; Wallace, J.C.; Maes, R.K.; Sordillo, L.M.; Bartlett, P.C.; Coussens, P.M. Reduced humoral immunity and atypical cell-mediated immunity in response to vaccination in cows naturally infected with bovine leukemia virus. Vet. Immunol. Immunopathol. 2016, 182, 125–135. [Google Scholar] [CrossRef]
- Erskine, R.J.; Bartlett, P.C.; Sabo, K.M.; Sordillo, L.M. Bovine Leukemia Virus Infection in Dairy Cattle: Effect on Serological Response to Immunization against J5 Escherichia coli Bacterin. Vet. Med. Int. 2011, 2011, 915747. [Google Scholar] [CrossRef]
- Martinez Cuesta, L.; Nieto Farias, M.V.; Lendez, P.A.; Barone, L.; Pérez, S.E.; Dolcini, G.L.; Ceriani, M.C. Stable infection of a bovine mammary epithelial cell line (MAC-T) with bovine leukemia virus (BLV). Virus Res. 2018, 256, 11–16. [Google Scholar] [CrossRef]
- Watanabe, A.; Murakami, H.; Kakinuma, S.; Murao, K.; Ohmae, K.; Isobe, N.; Akamatsu, H.; Seto, T.; Hashimura, S.; Konda, K.; et al. Association between bovine leukemia virus proviral load and severity of clinical mastitis. J. Vet. Med. Sci. 2019, 81, 1431–1437. [Google Scholar] [CrossRef]
- Rosewick, N.; Momont, M.; Durkin, K.; Takeda, H.; Caiment, F.; Cleuter, Y.; Vernin, C.; Mortreux, F.; Wattel, E.; Burny, A.; et al. Deep sequencing reveals abundant noncanonical retroviral microRNAs in B-cell leukemia/lymphoma. Proc. Natl. Acad. Sci. USA 2013, 110, 2306–2311. [Google Scholar] [CrossRef]
- Kincaid, R.P.; Burke, J.M.; Sullivan, C.S. RNA virus microRNA that mimics a B-cell oncomiR. Proc. Natl. Acad. Sci. USA 2012, 109, 3077–3082. [Google Scholar] [CrossRef]
- Gillet, N.A.; Hamaidia, M.; de Brogniez, A.; Gutiérrez, G.; Renotte, N.; Reichert, M.; Trono, K.; Willems, L. Bovine Leukemia Virus Small Noncoding RNAs Are Functional Elements That Regulate Replication and Contribute to Oncogenesis In Vivo. PLoS Pathog. 2016, 12, e1005588. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Sato, H.; Kubo, Y.; Wada, S.; Aida, Y. CAT1/SLC7A1 acts as a cellular receptor for bovine leukemia virus infection. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 14516–14527. [Google Scholar] [CrossRef]
- Safari, R.; Hamaidia, M.; de Brogniez, A.; Gillet, N.; Willems, L. Cis-drivers and trans-drivers of bovine leukemia virus oncogenesis. Curr. Opin. Virol. 2017, 26, 15–19. [Google Scholar] [CrossRef]
- Gendler, S.J. MUC1, the renaissance molecule. J. Mammary Gland Biol. Neoplasia 2001, 6, 339–353. [Google Scholar] [CrossRef]
- Sando, L.; Pearson, R.; Gray, C.; Parker, P.; Hawken, R.; Thomson, P.C.; Meadows, J.R.; Kongsuwan, K.; Smith, S.; Tellam, R.L. Bovine Muc1 is a highly polymorphic gene encoding an extensively glycosylated mucin that binds bacteria. J. Dairy Sci. 2009, 92, 5276–5291. [Google Scholar] [CrossRef] [PubMed]
- Rennie, W.; Liu, C.; Carmack, C.S.; Wolenc, A.; Kanoria, S.; Lu, J.; Long, D.; Ding, Y. STarMir: A web server for prediction of microRNA binding sites. Nucleic Acids Res. 2014, 42, W114–W118. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, C.; Mano, M.; Eulalio, A. MicroRNAs at the Host-Bacteria Interface: Host Defense or Bacterial Offense. Trends Microbiol. 2019, 27, 206–218. [Google Scholar] [CrossRef]
- Van Hese, I.; Goossens, K.; Vandaele, L.; Opsomer, G. Invited review: MicroRNAs in bovine colostrum-Focus on their origin and potential health benefits for the calf. J. Dairy Sci. 2020, 103, 1–15. [Google Scholar] [CrossRef]
- Tsukada, F.; Takashima, S.; Wakihara, Y.; Kamatari, Y.O.; Shimizu, K.; Okada, A.; Inoshima, Y. Characterization of miRNAs in Milk Small Extracellular Vesicles from Enzootic Bovine Leukosis Cattle. Int. J. Mol. Sci. 2022, 23, 10782. [Google Scholar] [CrossRef]
- Nakanishi, R.; Takashima, S.; Wakihara, Y.; Kamatari, Y.O.; Kitamura, Y.; Shimizu, K.; Okada, A.; Inoshima, Y. Comparing microRNA in milk small extracellular vesicles among healthy cattle and cattle at high risk for bovine leukemia virus transmission. J. Dairy Sci. 2022, 105, 5370–5380. [Google Scholar] [CrossRef]
- Ishikawa, H.; Rahman, M.M.; Yamauchi, M.; Takashima, S.; Wakihara, Y.; Kamatari, Y.O.; Shimizu, K.; Okada, A.; Inoshima, Y. mRNA Profile in Milk Extracellular Vesicles from Bovine Leukemia Virus-Infected Cattle. Viruses 2020, 12, 669. [Google Scholar] [CrossRef]
- Hiraoka, M.; Takashima, S.; Wakihara, Y.; Kamatari, Y.O.; Shimizu, K.; Okada, A.; Inoshima, Y. Identification of Potential mRNA Biomarkers in Milk Small Extracellular Vesicles of Enzootic Bovine Leukosis Cattle. Viruses 2022, 14, 1022. [Google Scholar] [CrossRef]
- Safari, R.; Jacques, J.R.; Brostaux, Y.; Willems, L. Ablation of non-coding RNAs affects bovine leukemia virus B lymphocyte proliferation and abrogates oncogenesis. PLoS Pathog. 2020, 16, e1008502. [Google Scholar] [CrossRef]
- Chen, J.; Xia, S.; Yang, X.; Chen, H.; Li, F.; Liu, F.; Chen, Z. Human Cytomegalovirus Encoded miR-US25-1-5p Attenuates CD147/EMMPRIN-Mediated Early Antiviral Response. Viruses 2017, 9, 365. [Google Scholar] [CrossRef]
- Liu, X.; Happel, C.; Ziegelbauer, J.M. Kaposi’s Sarcoma-Associated Herpesvirus MicroRNAs Target GADD45B To Protect Infected Cells from Cell Cycle Arrest and Apoptosis. J. Virol. 2017, 91, e02045-16. [Google Scholar] [CrossRef]
- Babu, S.G.; Pandeya, A.; Verma, N.; Shukla, N.; Kumar, R.V.; Saxena, S. Role of HCMV miR-UL70-3p and miR-UL148D in overcoming the cellular apoptosis. Mol. Cell. Biochem. 2014, 393, 89–98. [Google Scholar] [CrossRef]
- Frie, M.C.; Droscha, C.J.; Greenlick, A.E.; Coussens, P.M. MicroRNAs Encoded by Bovine Leukemia Virus (BLV) Are Associated with Reduced Expression of B Cell Transcriptional Regulators in Dairy Cattle Naturally Infected with BLV. Front. Vet. Sci. 2017, 4, 245. [Google Scholar] [CrossRef]
- Zyrianova, I.M.; Koval’chuk, S.N. Bovine leukemia virus pre-miRNA genes’ polymorphism. RNA Biol. 2018, 15, 1440–1447. [Google Scholar] [CrossRef] [PubMed]
- Casas, E.; Ma, H.; Lippolis, J.D. Expression of Viral microRNAs in Serum and White Blood Cells of Cows Exposed to Bovine Leukemia Virus. Front. Vet. Sci. 2020, 7, 536390. [Google Scholar] [CrossRef]
- Patton, S.; Gendler, S.J.; Spicer, A.P. The epithelial mucin, MUC1, of milk, mammary gland and other tissues. Biochim. Biophys. Acta 1995, 1241, 407–423. [Google Scholar] [CrossRef]
- DeSouza, M.M.; Surveyor, G.A.; Price, R.E.; Julian, J.; Kardon, R.; Zhou, X.; Gendler, S.; Hilkens, J.; Carson, D.D. MUC1/episialin: A critical barrier in the female reproductive tract. J. Reprod. Immunol. 1999, 45, 127–158. [Google Scholar] [CrossRef]
- Lillehoj, E.P.; Kim, H.; Chun, E.Y.; Kim, K.C. Pseudomonas aeruginosa stimulates phosphorylation of the airway epithelial membrane glycoprotein Muc1 and activates MAP kinase. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004, 287, L809–L815. [Google Scholar] [CrossRef] [PubMed]
- McGuckin, M.A.; Every, A.L.; Skene, C.D.; Linden, S.K.; Chionh, Y.T.; Swierczak, A.; McAuley, J.; Harbour, S.; Kaparakis, M.; Ferrero, R.; et al. Muc1 mucin limits both Helicobacter pylori colonization of the murine gastric mucosa and associated gastritis. Gastroenterology 2007, 133, 1210–1218. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence |
|---|---|
| siRNA1 | F: GCUGUUCCCAGUGCUUACA TT |
| R: UGUAAGCACUGGGAACAGC TT | |
| siRNA2 | F: GAAGCACACGCAGCCAGUUAU |
| R: AUAACUGGCUGCGUGUGCUUC TT | |
| siRNA3 | F: GCCACUUCCGCCAACUUGUAA TT |
| R: UUACAAGUUGGCGGAAGUGC TT | |
| siRNA NC | F: UUCUCCGAACGUGUCACGU TT |
| R: ACGUGACACGUUCGGAGAA TT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lian, S.; Liu, P.; Li, X.; Lv, G.; Song, J.; Zhang, H.; Wu, R.; Wang, D.; Wang, J. BLV-miR-B1-5p Promotes Staphylococcus aureus Adhesion to Mammary Epithelial Cells by Targeting MUC1. Animals 2023, 13, 3811. https://doi.org/10.3390/ani13243811
Lian S, Liu P, Li X, Lv G, Song J, Zhang H, Wu R, Wang D, Wang J. BLV-miR-B1-5p Promotes Staphylococcus aureus Adhesion to Mammary Epithelial Cells by Targeting MUC1. Animals. 2023; 13(24):3811. https://doi.org/10.3390/ani13243811
Chicago/Turabian StyleLian, Shuai, Pengfei Liu, Xiao Li, Guanxin Lv, Jiahe Song, Han Zhang, Rui Wu, Di Wang, and Jianfa Wang. 2023. "BLV-miR-B1-5p Promotes Staphylococcus aureus Adhesion to Mammary Epithelial Cells by Targeting MUC1" Animals 13, no. 24: 3811. https://doi.org/10.3390/ani13243811