
Conservation of Major Satellite DNAs in Snake Heterochromatin
 ,
,  , ,
, ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Line Establishment and Karyotype Analysis
2.2. Repetitive DNA Identification
2.3. Fluorescence In Situ Hybridization (FISH)
3. Results
3.1. Tandem Repeat Identification
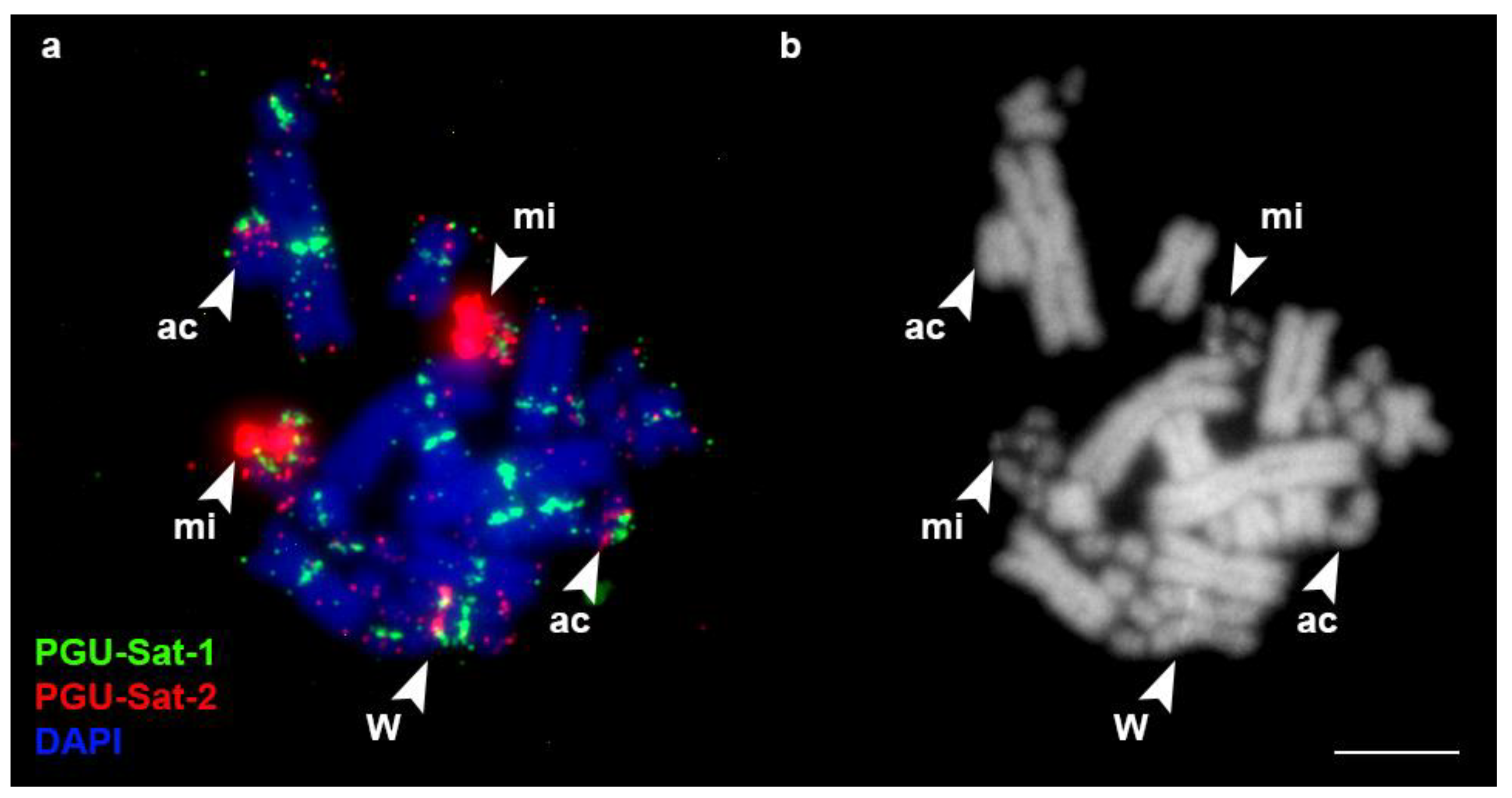
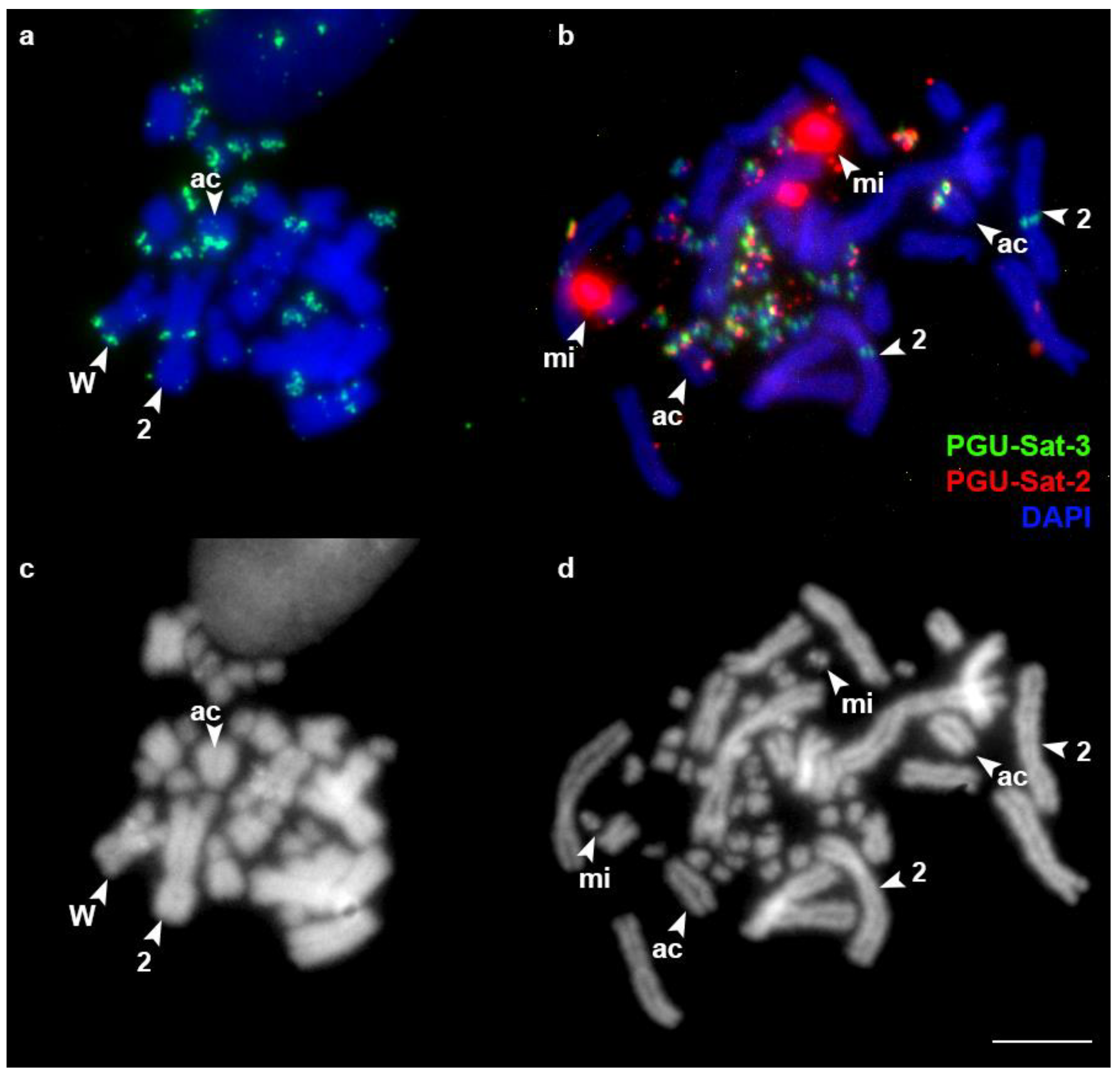
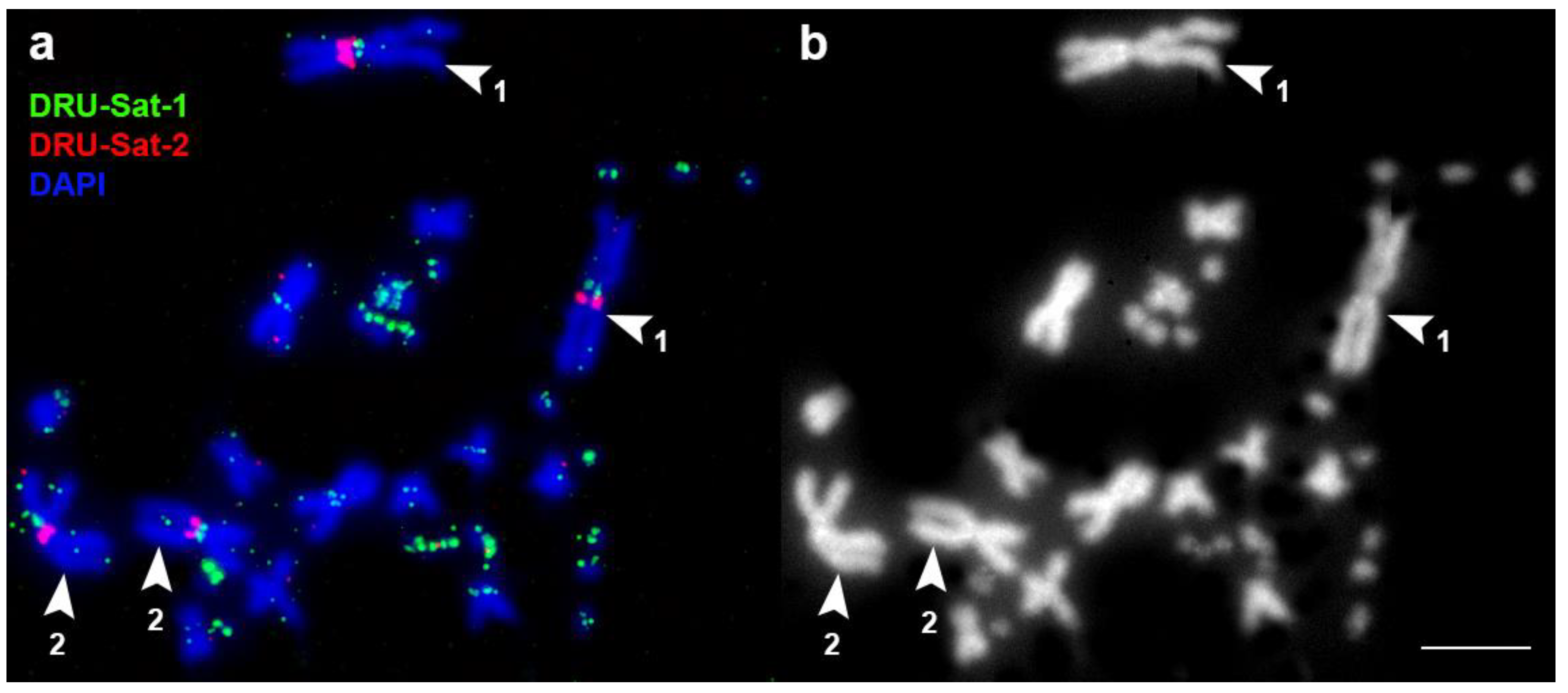
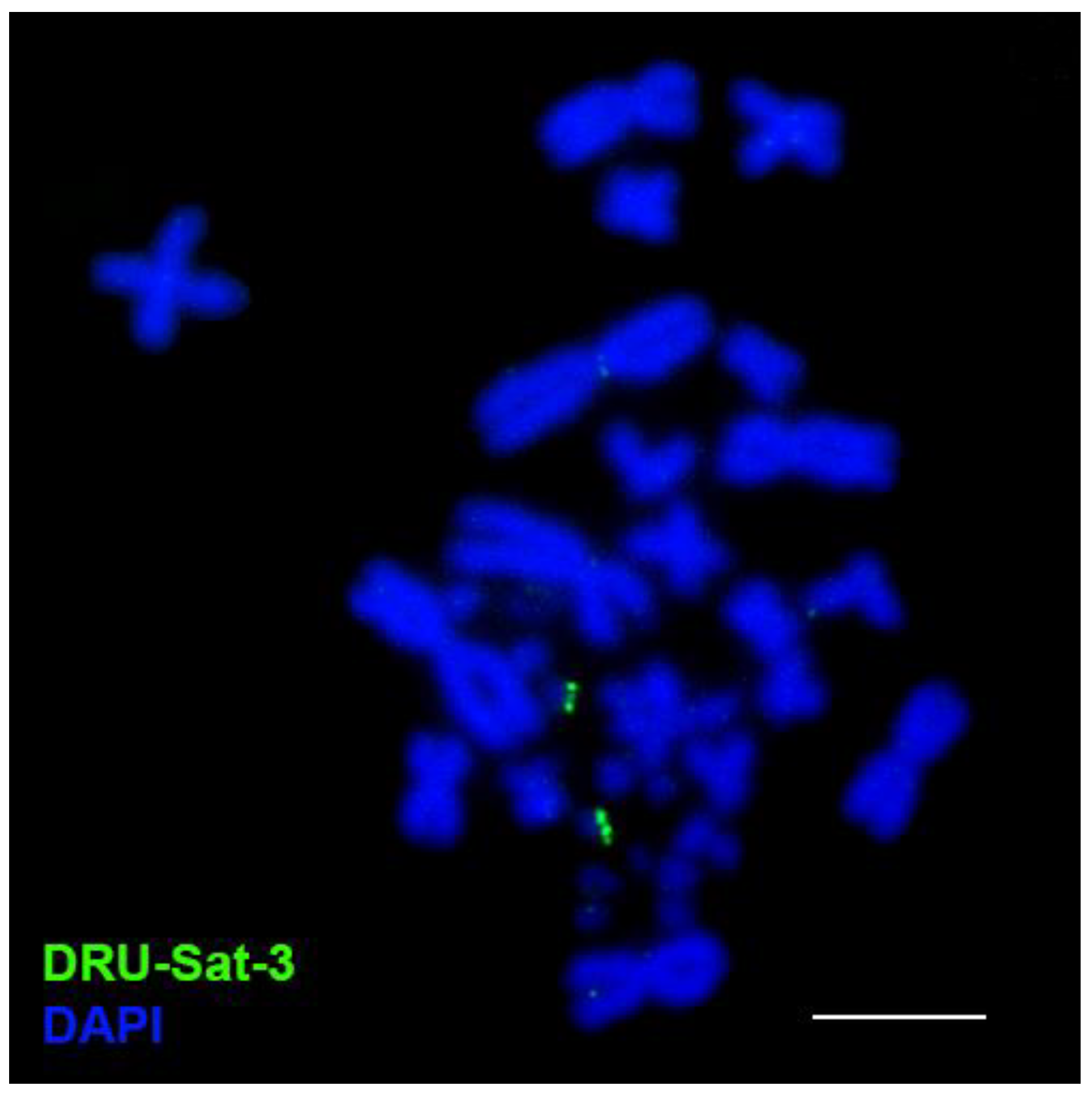
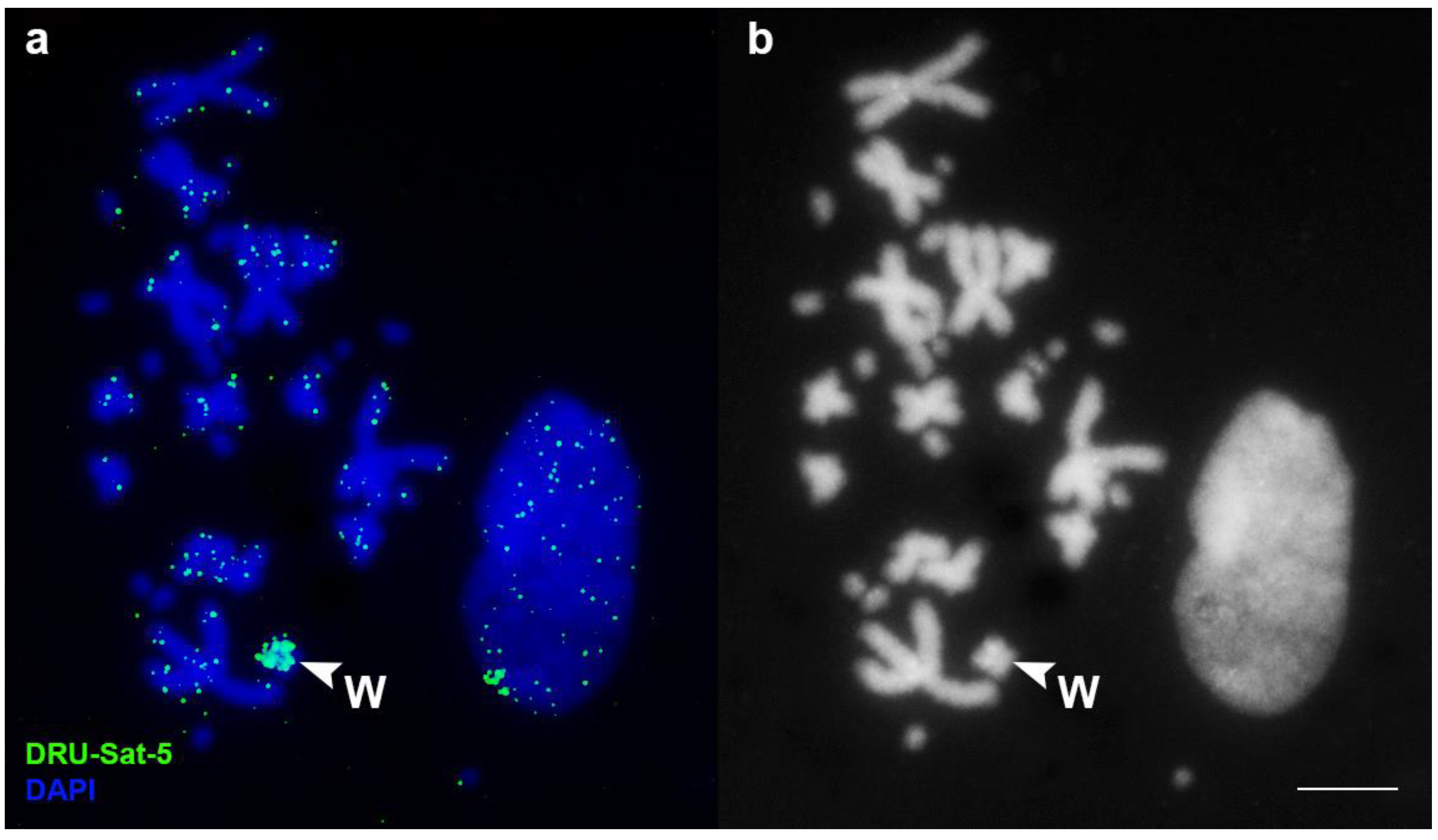
3.2. FISH Analysis
3.3. BLAST Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Biscotti, M.A.; Olmo, E.; Heslop-Harrison, J.S. Repetitive DNA in Eukaryotic Genomes. Chromosome Res. 2015, 23, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Ramos, M.A. Satellite DNA: An Evolving Topic. Genes 2017, 8, 230. [Google Scholar] [CrossRef] [PubMed]
- Plohl, M.; Luchetti, A.; Meštrović, N.; Mantovani, B. Satellite DNAs between Selfishness and Functionality: Structure, Genomics and Evolution of Tandem Repeats in Centromeric (Hetero) Chromatin. Gene 2008, 409, 72–82. [Google Scholar] [CrossRef]
- Navajas-Pérez, R.; de La Herrán, R.; Jamilena, M.; Lozano, R.; Rejón, C.R.; Rejón, M.R.; Garrido-Ramos, M.A. Reduced Rates of Sequence Evolution of Y-Linked Satellite DNA in Rumex (Polygonaceae). J. Mol. Evol. 2005, 60, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Meštrović, N.; Mravinac, B.; Pavlek, M.; Vojvoda-Zeljko, T.; Šatović, E.; Plohl, M. Structural and Functional Liaisons between Transposable Elements and Satellite DNAs. Chromosome Res. 2015, 23, 583–596. [Google Scholar] [CrossRef] [PubMed]
- Šatović, E.; Plohl, M. Tandem Repeat-Containing MITEs in the Clam Donax Trunculus. Genome Biol. Evol. 2013, 5, 2549–2559. [Google Scholar] [CrossRef] [Green Version]
- Satović, E.; Vojvoda Zeljko, T.; Luchetti, A.; Mantovani, B.; Plohl, M. Adjacent Sequences Disclose Potential for Intra-Genomic Dispersal of Satellite DNA Repeats and Suggest a Complex Network with Transposable Elements. BMC Genom. 2016, 17, 997. [Google Scholar] [CrossRef] [Green Version]
- Jagannathan, M.; Cummings, R.; Yamashita, Y.M. A Conserved Function for Pericentromeric Satellite DNA. Elife 2018, 7, e34122. [Google Scholar] [CrossRef]
- Spangenberg, V.; Losev, M.; Volkhin, I.; Smirnova, S.; Nikitin, P.; Kolomiets, O. DNA Environment of Centromeres and Non-Homologous Chromosomes Interactions in Mouse. Cells 2021, 10, 3375. [Google Scholar] [CrossRef] [PubMed]
- Biscotti, M.A.; Canapa, A.; Forconi, M.; Olmo, E.; Barucca, M. Transcription of Tandemly Repetitive DNA: Functional Roles. Chromosome Res. 2015, 23, 463–477. [Google Scholar] [CrossRef]
- Meštrović, N.; Plohl, M.; Mravinac, B.; Ugarković, D. Evolution of Satellite DNAs from the Genus Palorus—Experimental Evidence for the “library” Hypothesis. Mol. Biol. Evol. 1998, 15, 1062–1068. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, L.; Venanzetti, F.; Sbordoni, V. Characterization of a Species-Specific Satellite DNA Family of Dolichopoda Schiavazzii (Orthoptera, Rhaphidophoridae) Cave Crickets. J. Mol. Evol. 1994, 39, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Mravinac, B.; Plohl, M.; Ugarković, D. Preservation and High Sequence Conservation of Satellite DNAs Suggest Functional Constraints. J. Mol. Evol. 2005, 61, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, F.R.; de La Herrán, R.; Navajas-Pérez, R.; Cano-Roldán, B.; Sola-Campoy, P.J.; García-Zea, J.A.; Rejón, C.R. Centromeric Satellite DNA in Flatfish (Order Pleuronectiformes) and Its Relation to Speciation Processes. J. Hered. 2017, 108, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Capriglione, T.; Cardone, A.; Odierna, G.; Olmo, E. Evolution of a Centromeric Satellite DNA and Phylogeny of Lacertid Lizards. Comp. Biochem. Physiol. Part B Comp. Biochem. 1991, 100, 641–645. [Google Scholar] [CrossRef]
- Ciobanu, D.; Grechko, V.v.; Darevsky, I.S.; Kramerov, D.A. New Satellite DNA in Lacerta s. str. Lizards (Sauria: Lacertidae): Evolutionary Pathways and Phylogenetic Impact. J. Exp. Zool. B Mol. Dev. Evol. 2004, 302B, 505–516. [Google Scholar] [CrossRef]
- Giovannotti, M.; Nisi Cerioni, P.; Rojo, V.; Olmo, E.; Slimani, T.; Splendiani, A.; Caputo Barucchi, V. Characterization of a Satellite DNA in the Genera Lacerta and Timon (Reptilia, Lacertidae) and Its Role in the Differentiation of the W Chromosome. J. Exp. Zool. B Mol. Dev. Evol. 2018, 330, 83–95. [Google Scholar] [CrossRef]
- Giovannotti, M.; S’Khifa, A.; Nisi Cerioni, P.; Splendiani, A.; Slimani, T.; Fioravanti, T.; Olmo, E.; Caputo Barucchi, V. Isolation and Characterization of Two Satellite DNAs in Atlantolacerta andreanskyi (Werner, 1929) (Reptilia, Lacertidae). J. Exp. Zool. B Mol. Dev. Evol. 2020, 334, 178–191. [Google Scholar] [CrossRef]
- Rojo, V.; Martínez-Lage, A.; Giovannotti, M.; González-Tizón, A.M.; Cerioni, P.N.; Barucchi, V.C.; Galán, P.; Olmo, E.; Naveira, H. Evolutionary Dynamics of Two Satellite DNA Families in Rock Lizards of the Genus Iberolacerta (Squamata, Lacertidae): Different Histories but Common Traits. Chromosome Res. 2015, 23, 441–461. [Google Scholar] [CrossRef]
- Giovannotti, M.; Nisi Cerioni, P.; Caputo, V.; Olmo, E. Characterisation of a GC-Rich Telomeric Satellite DNA in Eumeces schneideri Daudin (Reptilia, Scincidae). Cytogenet. Genome Res. 2009, 125, 272–278. [Google Scholar] [CrossRef]
- Giovannotti, M.; Cerioni, P.N.; Splendiani, A.; Ruggeri, P.; Olmo, E.; Barucchi, V.C. Slow Evolving Satellite DNAs: The Case of a Centromeric Satellite in Chalcides ocellatus (Forskål, 1775) (Reptilia, Scincidae). Amphib. Reptil. 2013, 34, 401–411. [Google Scholar] [CrossRef]
- Chaiprasertsri, N.; Uno, Y.; Peyachoknagul, S.; Prakhongcheep, O.; Baicharoen, S.; Charernsuk, S.; Nishida, C.; Matsuda, Y.; Koga, A.; Srikulnath, K. Highly Species-Specific Centromeric Repetitive DNA Sequences in Lizards: Molecular Cytogenetic Characterization of a Novel Family of Satellite DNA Sequences Isolated from the Water Monitor Lizard (Varanus salvator macromaculatus, Platynota). J. Hered. 2013, 104, 798–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prakhongcheep, O.; Thapana, W.; Suntronpong, A.; Singchat, W.; Pattanatanang, K.; Phatcharakullawarawat, R.; Muangmai, N.; Peyachoknagul, S.; Matsubara, K.; Ezaz, T.; et al. Lack of Satellite DNA Species-Specific Homogenization and Relationship to Chromosomal Rearrangements in Monitor Lizards (Varanidae, Squamata). BMC Evol. Biol. 2017, 17, 193. [Google Scholar] [CrossRef] [Green Version]
- Yamada, K.; Nishida-Umehara, C.; Matsuda, Y. Molecular and Cytogenetic Characterization of Site-Specific Repetitive DNA Sequences in the Chinese Soft-Shelled Turtle (Pelodiscus sinensis, Trionychidae). Chromosome Res. 2005, 13, 33–46. [Google Scholar] [CrossRef]
- Romanenko, S.A.; Prokopov, D.Y.; Proskuryakova, A.A.; Davletshina, G.I.; Tupikin, A.E.; Kasai, F.; Ferguson-Smith, M.A.; Trifonov, V.A. The Cytogenetic Map of the Nile Crocodile (Crocodylus niloticus, Crocodylidae, Reptilia) with Fluorescence In Situ Localization of Major Repetitive DNAs. Int. J. Mol. Sci. 2022, 23, 13063. [Google Scholar] [CrossRef]
- Matsubara, K.; Uno, Y.; Srikulnath, K.; Seki, R.; Nishida, C.; Matsuda, Y. Molecular Cloning and Characterization of Satellite DNA Sequences from Constitutive Heterochromatin of the Habu Snake (Protobothrops flavoviridis, Viperidae) and the Burmese Python (Python bivittatus, Pythonidae). Chromosoma 2015, 124, 529–539. [Google Scholar] [CrossRef]
- Thongchum, R.; Singchat, W.; Laopichienpong, N.; Tawichasri, P.; Kraichak, E.; Prakhongcheep, O.; Sillapaprayoon, S.; Muangmai, N.; Baicharoen, S.; Suntrarachun, S.; et al. Diversity of PBI-DdeI Satellite DNA in Snakes Correlates with Rapid Independent Evolution and Different Functional Roles. Sci. Rep. 2019, 9, 15459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsubara, K.; Tarui, H.; Toriba, M.; Yamada, K.; Nishida-Umehara, C.; Agata, K.; Matsuda, Y. Evidence for different origin of sex chromosomes in snakes, birds, and mammals and step-wise differentiation of snake sex chromosomes. Proc. Natl. Acad. Sci. USA 2006, 103, 18190–18195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novák, P.; Robledillo, L.Á.; Koblížková, A.; Vrbová, I.; Neumann, P.; Macas, J. TAREAN: A Computational Tool for Identification and Characterization of Satellite DNA from Unassembled Short Reads. Nucleic Acids Res. 2017, 45, e111. [Google Scholar] [CrossRef] [PubMed]
- Graphodatsky, A.S.; Yang, F.; Serdukova, N.; Perelman, P.; Zhdanova, N.S.; Ferguson-Smith, M.A. Dog Chromosome-Specific Paints Reveal Evolutionary Inter- and Intrachromosomal Rearrangements in the American Mink and Human. Cytogenet. Genome Res. 2000, 90, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; O’Brien, P.C.M.; Milne, B.S.; Graphodatsky, A.S.; Solanky, N.; Trifonov, V.; Rens, W.; Sargan, D.; Ferguson-Smith, M.A. A Complete Comparative Chromosome Map for the Dog, Red Fox, and Human and Its Integration with Canine Genetic Maps. Genomics 1999, 62, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A Better Web Interface. Nucleic Acids Res. 2008, 36, W5–W9. [Google Scholar] [CrossRef]
- Bao, W.; Kojima, K.K.; Kohany, O. Repbase Update, a Database of Repetitive Elements in Eukaryotic Genomes. Mob DNA 2015, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- Owczarzy, R.; Tataurov, A.v.; Wu, Y.; Manthey, J.A.; McQuisten, K.A.; Almabrazi, H.G.; Pedersen, K.F.; Lin, Y.; Garretson, J.; McEntaggart, N.O.; et al. IDT SciTools: A Suite for Analysis and Design of Nucleic Acid Oligomers. Nucleic Acids Res. 2008, 36, W163–W169. [Google Scholar] [CrossRef] [PubMed]
- Biltueva, L.S.; Prokopov, D.Y.; Makunin, A.I.; Komissarov, A.S.; Kudryavtseva, A.v.; Lemskaya, N.A.; Vorobieva, N.V.; Serdyukova, N.A.; Romanenko, S.A.; Gladkikh, O.L.; et al. Genomic Organization and Physical Mapping of Tandemly Arranged Repetitive DNAs in Sterlet (Acipenser ruthenus). Cytogenet. Genome Res. 2017, 152, 148–157. [Google Scholar] [CrossRef]
- Romanenko, S.A.; Biltueva, L.S.; Serdyukova, N.A.; Kulemzina, A.I.; Beklemisheva, V.R.; Gladkikh, O.L.; Lemskaya, N.A.; Interesova, E.A.; Korentovich, M.A.; Vorobieva, N.v.; et al. Segmental Paleotetraploidy Revealed in Sterlet (Acipenser ruthenus) Genome by Chromosome Painting. Mol. Cytogenet. 2015, 8, 90. [Google Scholar] [CrossRef] [Green Version]
- Singchat, W.; Ahmad, S.F.; Sillapaprayoon, S.; Muangmai, N.; Duengkae, P.; Peyachoknagul, S.; O’Connor, R.E.; Griffin, D.K.; Srikulnath, K. Partial amniote sex chromosomal linkage homologies shared on snake W sex chromosomes support the ancestral super-sex chromosome evolution in amniotes. Front. Genet. 2020, 11, 948. [Google Scholar] [CrossRef]
- Augstenová, B.; Mazzoleni, S.; Kratochvíl, L.; Rovatsos, M. Evolutionary dynamics of the W chromosome in caenophidian snakes. Genes 2017, 9, 5. [Google Scholar] [CrossRef] [Green Version]
- Zaher, H.; Murphy, R.W.; Arredondo, J.C.; Graboski, R.; Machado-Filho, P.R.; Mahlow, K.; Montingelli, G.G.; Quadros, A.B.; Orlov, N.L.; Wilkinson, M.; et al. Large-Scale Molecular Phylogeny, Morphology, Divergence-Time Estimation, and the Fossil Record of Advanced Caenophidian Snakes (Squamata: Serpentes). PLoS One 2019, 14, e0216148. [Google Scholar] [CrossRef]
- Ruiz-Ruano, F.J.; López-León, M.D.; Cabrero, J.; Camacho, J.P.M. High-Throughput Analysis of the Satellitome Illuminates Satellite DNA Evolution. Sci. Rep. 2016, 6, 28333. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Satellite | Primer Sequences |
|---|---|
| PGU-Sat-1 | F 5’–TTTCAAGTACGAGCTTTCCC–3’ R 5’–GCTGAATTGAGCCCTACTG–3’ |
| PGU-Sat-2 | F 5’–GACACCAGGATGAGTTTCAG–3’ R 5’–TCCTGACCGTGGAGTAAA–3’ |
| PGU-Sat-3 | F 5’–CTTCCTCGGGCAGCAAA–3’ R 5’–GTAACAACGGATGCTAGAATGT–3’ |
| DRU-Sat-1 | F 5’–CCCGCCTGACCGAAGACC–3’ R 5’–GAGCTCTATCTGCAACGGG–3’ |
| DRU-Sat-2 | F 5’–ACCCCGAATCTCATTCTGGC–3’ R 5’–TCCTGATGCCGGGGTCAG–3’ |
| DRU-Sat-3 | F 5’–TTGTGTTTCTGGATCAATAACC–3’ R 5’–GCCTTTCCTGTATAATCCAAA–3’ |
| DRU-Sat-5 | F 5’–CAGAGCTGCTGGGAAGTG–3’ R 5’–GAGATCAATGAGGACCCCA–3’ |
| Sattelite Name | Monomer Size (bp) | Genome Proportion, % | GC | Species | Accession Number |
|---|---|---|---|---|---|
| Content, % | |||||
| DRU-Sat-1 | 168 | 0.3 | 42.3 | Daboia russelii (Russell’s viper) | OP820475 |
| DRU-Sat-2 | 170 | 0.13 | 35.9 | —//— | OP820476 |
| DRU-Sat-3 | 64 | 0.012 | 37.5 | —//— | OP820477 |
| DRU-Sat-5 | 147 | 0.025 | 44.9 | —//— | OP820478 |
| PGU-Sat-1 | 167 | 0.31 | 42.5 | Pantherophis guttatus | OP820479 |
| (Corn snake) | |||||
| PGU-Sat-2 | 187 | 0.085 | 37.4 | —//— | OP820480 |
| PGU-Sat-3 | 169 | 0.043 | 39.1 | —//— | OP820481 |
| PGU-Sat-2 | PGU-Sat-3 | DRU-Sat-2 | |
|---|---|---|---|
| PGU-Sat-2 | - | —//— | —//— |
| PGU-Sat-3 | 0.278 | - | —//— |
| DRU-Sat-2 | 0.262 | 0.226 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lisachov, A.; Rumyantsev, A.; Prokopov, D.; Ferguson-Smith, M.; Trifonov, V. Conservation of Major Satellite DNAs in Snake Heterochromatin. Animals 2023, 13, 334. https://doi.org/10.3390/ani13030334
Lisachov A, Rumyantsev A, Prokopov D, Ferguson-Smith M, Trifonov V. Conservation of Major Satellite DNAs in Snake Heterochromatin. Animals. 2023; 13(3):334. https://doi.org/10.3390/ani13030334
Chicago/Turabian StyleLisachov, Artem, Alexander Rumyantsev, Dmitry Prokopov, Malcolm Ferguson-Smith, and Vladimir Trifonov. 2023. "Conservation of Major Satellite DNAs in Snake Heterochromatin" Animals 13, no. 3: 334. https://doi.org/10.3390/ani13030334
APA StyleLisachov, A., Rumyantsev, A., Prokopov, D., Ferguson-Smith, M., & Trifonov, V. (2023). Conservation of Major Satellite DNAs in Snake Heterochromatin. Animals, 13(3), 334. https://doi.org/10.3390/ani13030334