
Fatty Acid Profiles in the Gonads of Red King Crab (Paralithodes camtschaticus) from the Barents Sea
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
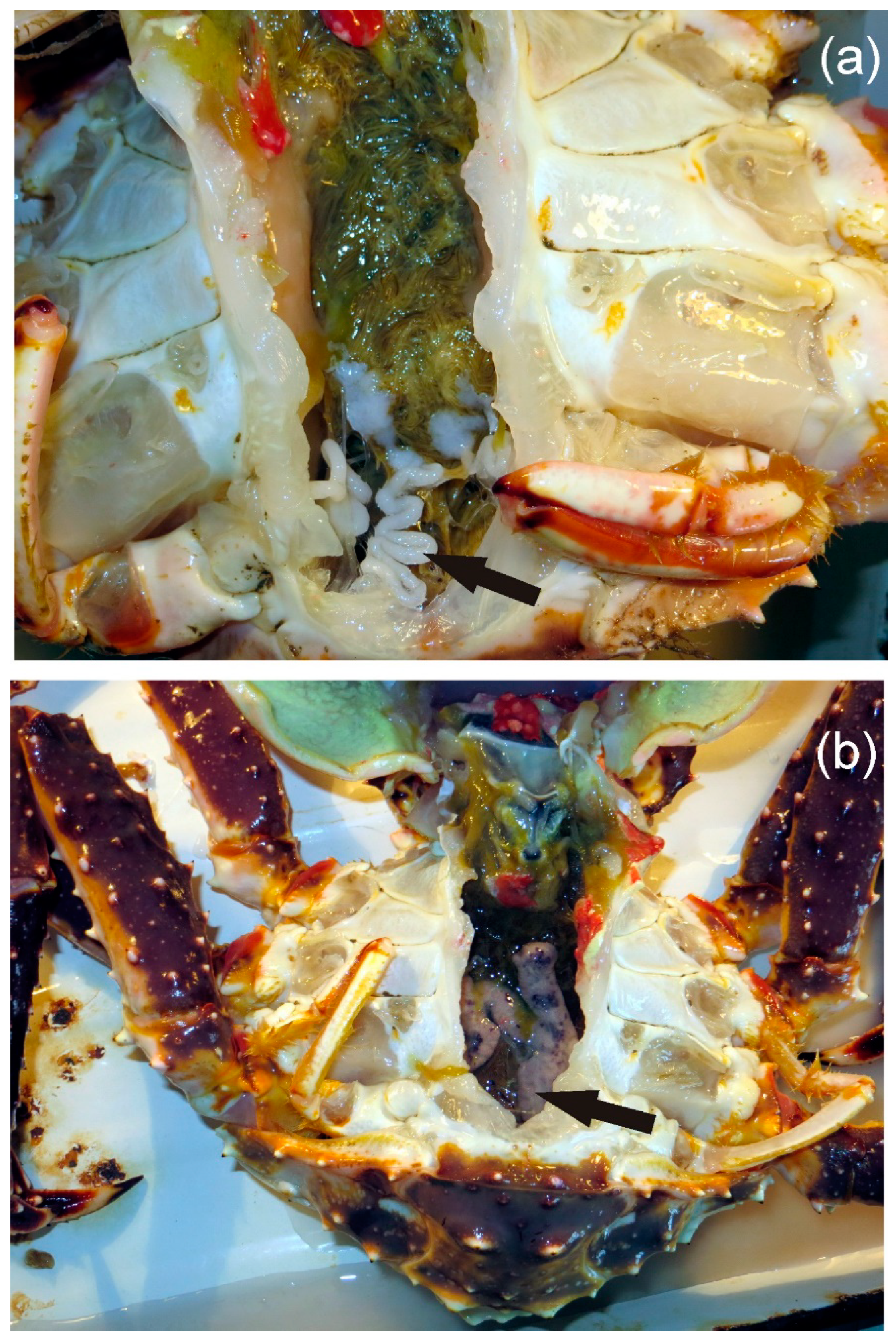
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kuzmin, S.A.; Gudimova, E.N. Introduction of the Kamchatka (Red King) Crab in the Barents Sea: Peculiarities of Biology, Perspectives of Fishery; KSC RAS Press: Apatity, Russia, 2002. (In Russian) [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Commercial fish and shellfish in the Barents Sea: Have introduced crab species affected the population trajectories of commercial fish? Rev. Fish Biol. Fish. 2015, 25, 297–322. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Red king crab (Paralithodes camtschaticus) fisheries in Russian waters: Historical review and present status. Rev. Fish Biol. Fish. 2018, 28, 331–353. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Inter-annual dynamics of the Barents Sea red king crab (Paralithodes camtschaticus) stock indices in relation to environmental factors. Polar Sci. 2016, 10, 541–552. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. New echinoderm-crab epibiotic associations from the coastal Barents Sea. Animals 2021, 11, 917. [Google Scholar] [CrossRef] [PubMed]
- Dvoretsky, A.G.; Dvoretsky, V.G. Epibiotic communities of common crab species in the coastal Barents Sea: Biodiversity and infestation patterns. Diversity 2022, 14, 6. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Ecology and distribution of red king crab larvae in the Barents Sea: A review. Water 2022, 14, 2328. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Renewal of the recreational red king crab fishery in Russian waters of the Barents Sea: Potential benefits and costs. Mar. Policy 2022, 136, 104916. [Google Scholar] [CrossRef]
- Pavlova, L.V.; Dvoretsky, A.G. Prey selectivity in juvenile red king crabs from the coastal Barents Sea. Diversity 2022, 14, 568. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Epibionts of an introduced king crab in the Barents Sea: A second five-year study. Diversity 2023, 15, 29. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Bichkaeva, F.A.; Baranova, N.F.; Dvoretsky, V.G. Fatty acids in the circulatory system of an invasive king crab from the Barents Sea. J. Food Compos. Anal. 2022, 110, 104528. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Bichkaeva, F.A.; Baranova, N.F.; Dvoretsky, V.G. Fatty acid composition of the Barents Sea red king crab (Paralithodes camtschaticus) leg meat. J. Food Compos. Anal. 2021, 98, 103826. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Tipisova, E.V.; Elfimova, A.E.; Alikina, V.A.; Dvoretsky, V.G. Sex hormones in hemolymph of red king crabs from the Barents Sea. Animals 2021, 11, 2149. [Google Scholar] [CrossRef] [PubMed]
- Dvoretsky, A.G.; Tipisova, E.V.; Alikina, V.A.; Elfimova, A.E.; Dvoretsky, V.G. Thyroid hormones in hemolymph of red king crabs from the Barents Sea. Animals 2022, 12, 379. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Bichkaeva, F.A.; Baranova, N.F.; Dvoretsky, V.G. Fatty acid composition in the hepatopancreas of the Barents Sea red king crab. Biol. Bull. 2020, 47, 332–338. [Google Scholar] [CrossRef]
- Lian, F.; Siikavuopio, S.I.; Harrison, S.M.; Vang, B.; Brunton, N.P.; Esaiassen, M.; Lorentzen, G. Fatty acid profile of cooked leg meat and raw hepatopancreas of red king crab (Paralithodes camtschaticus) during three-month live holding without feeding at 5 and 10 °C. J. Food Compos. Anal. 2022, 105, 104206. [Google Scholar] [CrossRef]
- Donaldson, W.E.; Byersdorfer, S.E. Biological Field Techniques for Lithodid Crabs. Fairbanks, Alaska Sea Grant College Program; University of Alaska: Fairbanks, AK, USA, 2005. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Effects of environmental factors on the abundance, biomass, and individual weight of juvenile red king crabs in the Barents Sea. Front. Mar. Sci. 2020, 7, 726. [Google Scholar] [CrossRef]
- Evseeva, O.Y.; Ishkulova, T.G.; Dvoretsky, A.G. Environmental drivers of an intertidal bryozoan community in the Barents Sea: A case study. Animals 2022, 12, 552. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Bichkaeva, F.A.; Vlasova, O.S.; Andronov, S.V.; Dvoretsky, V.G. Fatty acid content of four salmonid fish consumed by indigenous peoples from the Yamal-Nenets Autonomous Okrug (Northwestern Siberia, Russia). Animals 2022, 12, 1643. [Google Scholar] [CrossRef]
- Folch, J.; Less, M.; Sloane-Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Barrento, S.; Marques, A.; Teixeira, B.; Vaz-Pires, P.; Nunes, M.L. Nutritional quality of the edible tissues of European lobster Homarus gammarus and American lobster Homarus americanus. J. Agric. Food Chem. 2009, 57, 3645–3652. [Google Scholar] [CrossRef]
- Barrento, S.; Marques, A.; Teixeira, B.; Anacleto, P.; Vaz-Pires, P.; Nunes, M.L. Effect of season on the chemical composition and nutritional quality of the edible crab Cancer pagurus. J. Agric. Food Chem. 2009, 57, 10814–10824. [Google Scholar] [CrossRef] [PubMed]
- Taufik, M.; Shahrul, I.; Nordin, A.R.M.; Ikhwanuddin, M.; Abol-Munafi, A.B. Fatty acid composition of hepatopancreas and gonads in both sexes of orange mud crab, Scylla olivacea cultured at various water flow velocities. Trop. Life Sci. Res. 2020, 31, 79. [Google Scholar] [CrossRef] [PubMed]
- Mourente, G.; Medina, A.; Gonzalez, S.; Rodriguez, A. Changes in lipid class and fatty acid contents in the ovary and midgut gland of the female fiddler crab Uca tangeri (Decapoda, Ocypodiadae) during maturation. Mar. Biol. 1994, 121, 187–197. [Google Scholar] [CrossRef]
- Dinçer, M.T.; Aydin, İ. Proximate composition and mineral and fatty acid profiles of male and female jinga shrimps (Metapenaeus affinis, H. Milne Edwards, 1837). Turk. J. Vet. Anim. Sci. 2014, 38, 445–451. [Google Scholar] [CrossRef]
- Wen, X.; Chen, L.; Ai, C.; Zhou, Z.; Jiang, H. Variation in lipid composition of Chinese mitten-handed crab, Eriocheir sinensis during ovarian maturation. Compar. Biochem. Physiol. B 2001, 130, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, H.; Ma, T.; Sato, M.; Komiya, M.; Tadaki, D.; Hirano-Iwata, A. New aspects of bilayer lipid membranes for the analysis of ion channel functions. Membranes 2022, 12, 863. [Google Scholar] [CrossRef]
- Wathes, D.C.; Cheng, Z. Polyunsaturated fatty biosynthesis and metabolism in reproductive tissues. In Polyunsaturated Fatty Acid Metabolism; Burdge, G.C., Ed.; Academic Press: London, UK, 2018; pp. 157–180. [Google Scholar]
- Subramoniam, T. Sexual Biology and Reproduction in Crustaceans; Academic Press: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Silkovsky, J.; Chayoth, R.; Sagi, A. Comparative study of effects of Prostaglandin E 2 on cAMP levels in gonads of the prawn, Macrobrachium rosenbergii and the crayfish Cherax quadricarinatus. J. Crust. Biol. 1998, 18, 643–649. [Google Scholar] [CrossRef]
- Peixoto, S.; Wasielesky, W.; Martino, R.C.; Milach, A.; Soares, R.; Cavalli, R.O. Comparison of reproductive output, offspring quality, ovarian histology and fatty acid composition between similarly-sized wild and domesticated Farfantepenaeus paulensis. Aquaculture 2008, 285, 201–206. [Google Scholar] [CrossRef]
- Reznick, D. Costs of reproduction: An evaluation of the empirical evidence. Oikos 1985, 44, 257–267. [Google Scholar] [CrossRef]
- Allen, W.V. Amino acid and fatty acid composition of tissues of the Dungeness crab (Cancer magister). J. Fish. Res. Bd. Can. 1971, 28, 1191–1195. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Population dynamics of the invasive lithodid crab, Paralithodes camtschaticus, in a typical bay of the Barents Sea. ICES J. Mar. Sci. 2013, 70, 1255–1262. [Google Scholar] [CrossRef]
- Dvoretsky, A.G.; Dvoretsky, V.G. Size-at-age of juvenile red king crab (Paralithodes camtschaticus) in the coastal Barents Sea. Cah. Biol. Mar. 2014, 55, 43–48. [Google Scholar]
- Dvoretsky, A.G.; Dvoretsky, V.G. Size at maturity of female red king crab, Paralithodes camtschaticus, from the costal zone of Kola Peninsula (southern Barents Sea). Cah. Biol. Mar. 2015, 56, 49–54. [Google Scholar]
- Balta, I.; Stef, L.; Pet, I.; Iancu, T.; Stef, D.; Corcionivoschi, N. Essential fatty acids as biomedicines in cardiac health. Biomedicines 2021, 9, 1466. [Google Scholar] [CrossRef] [PubMed]
- Tsoupras, A.; Brummell, C.; Kealy, C.; Vitkaitis, K.; Redfern, S.; Zabetakis, I. Cardio-protective properties and health benefits of fish lipid bioactives; the effects of thermal processing. Mar. Drugs 2022, 20, 187. [Google Scholar] [CrossRef]
- Cordingley, D.M.; Cornish, S.M. Omega-3 fatty acids for the management of osteoarthritis: A narrative review. Nutrients 2022, 14, 3362. [Google Scholar] [CrossRef] [PubMed]
- Xiong, R.-G.; Zhou, D.-D.; Wu, S.-X.; Huang, S.-Y.; Saimaiti, A.; Yang, Z.-J.; Shang, A.; Zhao, C.-N.; Gan, R.-Y.; Li, H.-B. Health benefits and side effects of short-chain fatty acids. Foods 2022, 11, 2863. [Google Scholar] [CrossRef]
- Gerber, M. Omega-3 fatty acids and cancers: A systematic update of epidemiological studies. Brit. J. Nutr. 2012, 107 (Suppl. S2), S228–S239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobin-Dubigeon, C.; Nazih, H.; Croyal, M.; Bard, J.-M. Link between omega 3 fatty acids carried by lipoproteins and breast cancer severity. Nutrients 2022, 14, 2461. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, C.; Sansone, A.; Chatgilialoglu, C.; Ferreri, R.; Amézaga, J.; Burgos, M.C.; Arranz, S.; Tueros, I. Critical review on fatty acid-based food and nutraceuticals as supporting therapy in cancer. Int. J. Mol. Sci. 2022, 23, 6030. [Google Scholar] [CrossRef] [PubMed]
- Dembitsky, V.M. Hydrobiological aspects of fatty acids: Unique, rare, and unusual fatty acids incorporated into linear and cyclic lipopeptides and their biological activity. Hydrobiology 2022, 1, 331–432. [Google Scholar] [CrossRef]
- Panse, M.L.; Phalke, S.D. World market of omega-3 fatty acids. In Omega-3 Fatty Acids; Hegde, M., Zanwar, A., Adekar, S., Eds.; Springer: Cham, The Netherlands, 2016; pp. 79–88. [Google Scholar]
- Valero-Vello, M.; Peris-Martínez, C.; García-Medina, J.J.; Sanz-González, S.M.; Ramírez, A.I.; Fernández-Albarral, J.A.; Galarreta-Mira, D.; Zanón-Moreno, V.; Casaroli-Marano, R.P.; Pinazo-Duran, M.D. Searching for the antioxidant, anti-inflammatory, and neuroprotective potential of natural food and nutritional supplements for ocular health in the Mediterranean population. Foods 2021, 10, 1231. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Sex | Carapace Length, mm | Weight, g | |||||||
|---|---|---|---|---|---|---|---|---|---|
| N | X | SE | Min | Max | X | SE | Min | Max | |
| Female | 40 | 147.6 | 2.0 | 101.0 | 173.5 | 1810.6 | 63.2 | 591 | 2633 |
| Male | 8 | 174.4 | 11.9 | 122.5 | 230.0 | 3090.1 | 631.6 | 980 | 6320 |
| Female + Male | 48 | 152.0 | 2.9 | 101.0 | 230.0 | 2023.9 | 132.3 | 591 | 6320 |
| Fatty Acid | Concentration, μg·g−1 | % of Total Fatty Acid | p | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Female | Male | Female | Male | ||||||
| X | SD | X | SD | X | SD | X | SD | ||
| C6:0 | 1.9 | 1.2 | 1.0 | 0.2 | 0.009 | 0.008 | 0.032 | 0.012 | 0.115 |
| C8:0 | 2.1 | 0.7 | 2.5 | 1.0 | 0.013 | 0.017 | 0.084 | 0.041 | 0.185 |
| C9:0 | 2.2 | 0.8 | 2.5 | 1.1 | 0.012 | 0.011 | 0.081 | 0.041 | 0.379 |
| C10:0 | 2.9 | 1.6 | 2.4 | 1.1 | 0.017 | 0.016 | 0.079 | 0.040 | 0.342 |
| C11:0 | 1.8 | 1.0 | 2.1 | 1.3 | 0.010 | 0.008 | 0.067 | 0.042 | 0.460 |
| C12:0 | 49.2 | 14.6 | 15.0 | 6.8 | 0.248 | 0.100 | 0.463 | 0.253 | <0.001 |
| C13:0 | 7.3 | 2.1 | 2.3 | 0.7 | 0.036 | 0.012 | 0.073 | 0.032 | <0.001 |
| C14:0 | 348.8 | 154.9 | 45.7 | 16.0 | 1.536 | 0.295 | 1.316 | 0.233 | <0.001 |
| C15:0 | 169.5 | 68.0 | 23.3 | 10.5 | 0.767 | 0.181 | 0.652 | 0.102 | <0.001 |
| C16:0 | 3463.8 | 1254.4 | 672.3 | 339.5 | 15.776 | 1.249 | 18.445 | 2.626 | <0.001 |
| C17:0 | 132.2 | 40.8 | 33.8 | 20.9 | 0.628 | 0.128 | 0.898 | 0.113 | <0.001 |
| C18:0 | 1172.7 | 355.7 | 406.1 | 160.9 | 5.648 | 1.392 | 11.621 | 2.366 | <0.001 |
| C20:0 | 53.9 | 23.6 | 15.0 | 4.7 | 0.245 | 0.092 | 0.438 | 0.082 | <0.001 |
| C21:0 | 12.7 | 10.0 | 2.9 | 0.8 | 0.056 | 0.034 | 0.092 | 0.031 | <0.001 |
| C22:0 | 16.5 | 8.1 | 6.0 | 2.0 | 0.079 | 0.040 | 0.187 | 0.071 | <0.001 |
| C23:0 | 83.2 | 59.9 | 9.3 | 7.6 | 0.356 | 0.220 | 0.228 | 0.118 | <0.001 |
| C24:0 | 94.5 | 57.8 | 15.0 | 11.0 | 0.424 | 0.208 | 0.383 | 0.082 | <0.001 |
| C14:1n5t | – | – | 2.9 | 0.0 | – | – | 0.005 | 0.012 | – |
| C14:1n5c | 4.3 | 1.8 | 2.4 | 1.8 | 0.020 | 0.008 | 0.071 | 0.059 | <0.001 |
| C15:1 | 13.6 | 18.1 | 5.1 | 5.3 | 0.039 | 0.067 | 0.068 | 0.121 | 0.411 |
| C16:1n7t | 34.4 | 18.2 | 8.9 | 8.1 | 0.159 | 0.079 | 0.209 | 0.092 | <0.001 |
| C16:1n7c | 1538.8 | 752.9 | 91.7 | 44.1 | 6.614 | 1.793 | 2.533 | 0.727 | <0.001 |
| C17:1 | 3.0 | 2.2 | 1.6 | 1.3 | 0.014 | 0.012 | 0.051 | 0.055 | <0.001 |
| C18:1n9t | 90.0 | 32.9 | 18.9 | 5.7 | 0.431 | 0.139 | 0.564 | 0.151 | <0.001 |
| C18:1n9c | 2709.1 | 979.7 | 490.1 | 313.7 | 12.453 | 1.577 | 12.922 | 1.280 | <0.001 |
| C20:1 | 552.2 | 260.1 | 93.6 | 83.7 | 2.436 | 0.611 | 2.291 | 0.851 | <0.001 |
| C22:1 | 50.0 | 26.2 | 8.5 | 5.7 | 0.223 | 0.073 | 0.234 | 0.086 | <0.001 |
| C24:1 | 29.1 | 15.1 | 4.2 | 1.8 | 0.130 | 0.045 | 0.122 | 0.028 | <0.001 |
| C18:2n6t | 36.0 | 33.7 | 15.2 | 9.4 | 0.164 | 0.154 | 0.400 | 0.123 | <0.001 |
| C18:2n6C | 275.9 | 105.0 | 58.7 | 46.8 | 1.283 | 0.277 | 1.488 | 0.252 | <0.001 |
| C18:3n3 | 91.6 | 44.2 | 10.7 | 14.4 | 0.409 | 0.125 | 0.225 | 0.130 | <0.001 |
| C18:3n6 | 53.2 | 21.8 | 7.7 | 7.8 | 0.245 | 0.071 | 0.180 | 0.054 | <0.001 |
| C20:2n6 | 208.0 | 73.5 | 46.8 | 32.1 | 0.976 | 0.171 | 1.200 | 0.177 | <0.001 |
| C20:3n6 | 35.1 | 17.7 | 5.2 | 3.2 | 0.156 | 0.037 | 0.137 | 0.036 | <0.001 |
| C20:4n6 | 1240.4 | 464.5 | 311.2 | 184.6 | 5.824 | 1.218 | 8.179 | 1.195 | <0.001 |
| C22:2n6 | 2.4 | 1.4 | 0.7 | 0.2 | 0.011 | 0.005 | 0.021 | 0.007 | <0.001 |
| C20:5n3 | 5465.4 | 2160.6 | 721.3 | 486.9 | 24.448 | 3.067 | 18.429 | 2.005 | <0.001 |
| C22:6n3 | 3463.1 | 1255.2 | 566.0 | 497.8 | 15.911 | 2.375 | 13.833 | 2.818 | <0.001 |
| C20:3n3 | 46.5 | 25.9 | 7.2 | 5.8 | 0.211 | 0.066 | 0.181 | 0.076 | <0.001 |
| C22:4n6 | 69.7 | 30.3 | 12.6 | 5.3 | 0.315 | 0.074 | 0.362 | 0.132 | <0.001 |
| C22:3n3 | 2.2 | 1.9 | 0.4 | 0.0 | 0.010 | 0.008 | 0.003 | 0.008 | <0.001 |
| C22:5n6 | 63.6 | 23.7 | 8.5 | 6.7 | 0.292 | 0.084 | 0.212 | 0.080 | <0.001 |
| C22:5n3 | 305.8 | 130.8 | 34.7 | 19.3 | 1.364 | 0.220 | 0.939 | 0.334 | <0.001 |
| ∑SFA | 5611.7 | 1903.6 | 1257.3 | 568.3 | 25.861 | 2.006 | 35.139 | 5.667 | <0.001 |
| ∑MUFA | 5018.8 | 1940.5 | 723.4 | 458.2 | 22.520 | 2.273 | 19.070 | 1.955 | <0.001 |
| ∑PUFA | 11357.9 | 4004.0 | 1806.7 | 1297.3 | 51.620 | 2.463 | 45.791 | 4.548 | <0.001 |
| Total | 21988.4 | 7732.4 | 3787.3 | 2307.9 | 100.000 | 0.000 | 100.000 | 0.000 | <0.001 |
| ∑n-3 | 9374.5 | 3363.5 | 1340.0 | 1013.2 | 42.353 | 2.290 | 33.611 | 4.063 | <0.001 |
| ∑n-6 | 1983.4 | 703.6 | 466.6 | 287.4 | 9.266 | 1.433 | 12.180 | 1.054 | <0.001 |
| ∑n-9 | 3430.4 | 1273.6 | 615.4 | 408.0 | 15.674 | 1.903 | 16.133 | 2.142 | <0.001 |
| ∑n-7 | 1575.8 | 760.3 | 102.3 | 51.1 | 6.787 | 1.794 | 2.793 | 0.663 | <0.001 |
| n-3/n-6 | 4.7 | 0.8 | 2.8 | 0.3 | 4.682 | 0.777 | 2.767 | 0.333 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dvoretsky, A.G.; Bichkaeva, F.A.; Baranova, N.F.; Dvoretsky, V.G. Fatty Acid Profiles in the Gonads of Red King Crab (Paralithodes camtschaticus) from the Barents Sea. Animals 2023, 13, 336. https://doi.org/10.3390/ani13030336
Dvoretsky AG, Bichkaeva FA, Baranova NF, Dvoretsky VG. Fatty Acid Profiles in the Gonads of Red King Crab (Paralithodes camtschaticus) from the Barents Sea. Animals. 2023; 13(3):336. https://doi.org/10.3390/ani13030336
Chicago/Turabian StyleDvoretsky, Alexander G., Fatima A. Bichkaeva, Nina F. Baranova, and Vladimir G. Dvoretsky. 2023. "Fatty Acid Profiles in the Gonads of Red King Crab (Paralithodes camtschaticus) from the Barents Sea" Animals 13, no. 3: 336. https://doi.org/10.3390/ani13030336