

Identifying the Effects of Social Disruption through Translocation on African Elephants (Loxodonta africana), with Specifics on the Social and Ecological Impacts of Orphaning

Abstract
:Simple Summary
Abstract
1. Introduction
- The Orphans would be less competent in rearing their young than the A&B herd, demonstrated by a younger average age of death and a higher calf mortality rate [26,43,44], and we expected this to be reflected in rates of vigilance behaviour, with the Orphans exhibiting less awareness of danger and the appropriate response;
- Except for possibly two of the founding members, the Orphans are not thought to be kin and are instead remnants of different herds pieced together for translocation from KNP. We, therefore, expected the Orphans to be less cohesive as a unit, causing the group to fission more frequently and exhibit less affiliative behaviour with herd mates (and also because they were not exposed to such behaviours by their own mothers and allomothers);
- Despite the founding members of the Orphans all being similar in age, as noted in other orphaned elephant groups [49], we expected one individual to assume the role of matriarch. However, whilst matriarchs typically act as a vital repository of social and ecological knowledge, we assumed that the Orphans’ matriarch would lack much of this knowledge due to early-age orphaning and thus no access to older females to learn from example. We expected this to play out as a less-definitive leadership approach demonstrated through her position relative to her herd and the decisiveness of herd movement decisions.
2. Methods and Material
2.1. History of the PGR Elephants
- Vasectomising seven subadult bulls in 2008;
- GnRH contraceptive treatment of the two oldest bulls (unable to undergo vasectomies) from 2008 until their deaths in 2010 and 2013;
- Destruction of a dominant, mature A&B herd cow in 2011;
- GnRH treatment of five subadult bulls from 2016–2017 as a means of contraception
- Hunting of adult bulls in 2010 and 2013;
- Forced movement of the A&B herd from the Royal Jozini Private Game Reserve (RJ) to the Pongola Nature Reserve Eastern Shores on 11 October 2016, during which time the herd was split, and a subset of the A&B herd (n = 7) remained in RJ. This separation persisted until the end of the study period.
2.2. Data Collection
Demographic and Behavioural Monitoring
2.3. Data Analyses
3. Results
3.1. Calf-Rearing Ability
Age of Death and Infant Mortality
3.2. Vigilance Behaviour
3.3. Cohesiveness
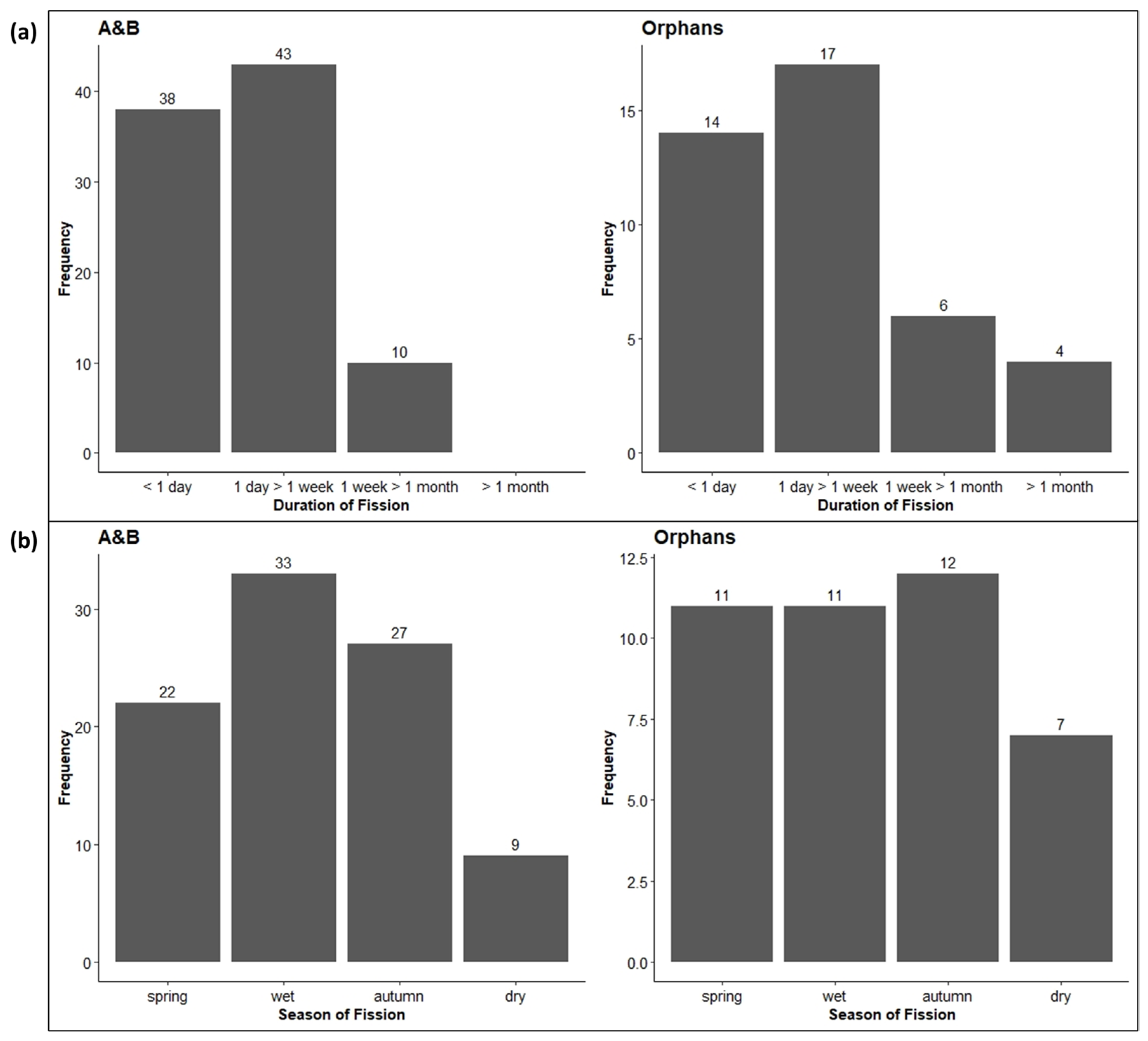
3.3.1. Fission–Fusion Dynamics
3.3.2. Affiliative Behaviour
3.4. Accelerated Reproduction
3.4.1. Age at First Parturition
3.4.2. Inter-Birth Intervals
3.4.3. Sex Ratios
3.5. Decision Making
3.5.1. Leadership
3.5.2. Space Use
4. Discussion
4.1. Calf Rearing Ability
4.2. Cohesiveness
4.3. Affiliative Interactions
4.4. Accelerated Reproduction
4.5. Sex Ratios
4.6. Decision-Making and the Role of Matriarchs
4.7. Effect of Translocation on the A&B Herd
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heyes, C.M. Social learning in animals: Categories and Mechanisms. Biol. Rev. 1994, 69, 207–231. [Google Scholar] [CrossRef] [PubMed]
- Taborsky, B.; Oliveira, R.F. Social competence: An evolutionary approach. Trends Ecol. Evol. 2012, 27, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Kempes, M.M.; Gulickx, M.M.C.; van Daalen, H.J.C.; Louwerse, A.L.; Sterck, E.H.M. Social competence is reduced in socially deprived Rhesus monkeys (Macaca mulatta). J. Comp. Psychol. 2008, 122, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Wechsler, B. Coping and coping strategies: A behavioural view. Appl. Anim. Behav. Sci. 1995, 43, 123–134. [Google Scholar] [CrossRef]
- McComb, K.; Shannon, G.; Durant, S.M.; Sayialel, K.; Slotow, R.; Poole, J.; Moss, C. Leadership in elephants: The adaptive value of age. Proc. R. Soc. B 2011, 278, 3270–3276. [Google Scholar] [CrossRef] [Green Version]
- Snyder-Mackler, N.; Burger, J.R.; Gaydosh, L.; Belsky, D.W.; Noppert, G.A.; Campos, F.A.; Bartolomucci, A.; Claire Yang, C.; Aiello, A.E.; O’Rand, A.; et al. Social determinants of health and survival in humans and other animals. Science 2020, 368, 843. [Google Scholar] [CrossRef] [PubMed]
- Holt-Lunstad, J.H.; Smith, T.M.; Layton, J.B. Social relationships and mortality risk: A meta-analytic review. PLoS Med. 2010, 7, e1000316. [Google Scholar] [CrossRef]
- Silk, J.B.; Beehner, J.C.; Bergman, T.J.; Crockford, C.; Engh, A.L.; Moscovice, L.R.; Wittig, R.M.; Seyfarth, R.M. Strong and consistent social bonds enhance longevity of female baboons. Curr. Biol. 2010, 20, 1359–1361. [Google Scholar] [CrossRef] [Green Version]
- Silk, J.B.; Alberts, S.C.; Altmann, J. Social bonds of female baboons enhance infant survival. Science 2003, 302, 1231–1234. [Google Scholar] [CrossRef] [Green Version]
- Byrne, R.W.; Bates, L.A. Sociality, Evolution and Cognition. Curr. Biol. 2007, 17, R714–R723. [Google Scholar] [CrossRef]
- Connor, R.C. Dolphin social intelligence: Complex alliance relationships in bottlenose dolphins and a consideration of selective environments for extreme brain size evolution in mammals. Phil. Trans. R. Soc. B 2007, 362, 587–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheney, D.; Seyfarth, R.; Smuts, B. Social Relationships and social cognition in non-human primates. Science 1986, 234, 1361–1366. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.C.; Moss, C.J. Calf development and maternal rearing strategies. In The Amboseli Elephants: A Long-Term Perspective on a Long-Lived Mammal; Moss, C.J., Croze, H., Lee, P.C., Eds.; University of Chicago Press: Chicago, IL, USA, 2011; pp. 224–245. [Google Scholar]
- Pinter-Wollman, N.; Isbell, L.A.; Hart, L.A. The relationship between social behaviour and habitat familiarity in African elephants (Loxodonta africana). Proc. R. Soc. B 2009, 276, 1009–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedwig, D.; Poole, J.; Granli, P. Does Social Complexity Drive Vocal Complexity? Insights from the Two African Elephant Species. Animals 2021, 11, 3017. [Google Scholar] [CrossRef] [PubMed]
- Hart, B.L.; Hart, L.A.; McCoy, M.; Sarath, C.R. Cognitive behaviour in Asian elephants: Use and modification of branches for fly switching. Anim. Behav. 2001, 62, 839–847. [Google Scholar] [CrossRef] [Green Version]
- Kurt, F.; Garaï, M.E. The Asian Elephant in Captivity; Foundation Books, Cambridge University Press: New Delhi, India, 2007. [Google Scholar]
- Thornton, A.; Clutton-Brock, T. Social learning and the development of individual and group behaviour in mammal societies. Phil. Trans. R. Soc. B 2011, 366, 978–987. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, H.; Lusseau, D. Animal social networks as substrate for cultural behavioural diversity. J. Theor. Biol. 2012, 294, 19–28. [Google Scholar] [CrossRef]
- Esposito, R.M. Effect of Matriarchs on Group Interactions, Kinship Fitness, and Differences in Chemosensory Behaviour of African Elephants (Loxodonta africana). MSc Thesis, Georgia Southern University, Statesboro, GA, USA, 2008. Available online: https://digitalcommons.georgiasouthern.edu/etd/716 (accessed on 18 March 2022).
- Goldenberg, S.Z.; Wittemyer, G. Orphaned female elephant social bonds reflect lack of access to mature adults. Sci. Rep. 2017, 7, 14408. [Google Scholar] [CrossRef] [Green Version]
- Gobush, K.; Kerr, B.; Wasser, S. Genetic relatedness and disrupted social structure in a poached population of African elephants. Mol. Ecol. 2009, 18, 722–734. [Google Scholar] [CrossRef]
- Foley, C.A.H.; Faust, L.J. Rapid population growth in an elephant Loxodonta africana population recovering from poaching in Tarangire National Park, Tanzania. Oryx 2010, 44, 205–212. [Google Scholar] [CrossRef]
- Shannon, G.; Slotow, R.; Durant, S.M.; Sayialel, K.N.; Poole, J.; Moss, C.; McComb, K. Effects of social disruption in elephants persist decades after culling. Front. Zool. 2013, 10, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foley, C.A.H.; Pettorelli, N.; Foley, L. Severe drought and calf survival in elephants. Biol. Lett. 2008, 4, 541–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McComb, K.; Moss, C.; Durant, S.M.; Baker, L.; Sayialel, S. Matriarchs as repositories of social knowledge in African elephants. Science 2001, 292, 491–494. [Google Scholar] [CrossRef] [Green Version]
- Wittemyer, G.; Douglas-Hamilton, I.; Getz, W.M. The socioecology of elephants: Analysis of the processes creating multitiered social structures. Anim. Behav. 2005, 69, 1357–1371. [Google Scholar] [CrossRef] [Green Version]
- Lee, P.C.; Bussière, L.F.; Webber, C.E.; Poole, J.H.; Moss, C.J. Enduring consequences of early experiences: 40 year effects on survival and success among African elephants (Loxodonta africana). Biol. Lett. 2013, 9, 20130011. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, G.A.; Schore, A.N.; Brown, J.L.; Poole, J.H.; Moss, C.J. Elephant Breakdown. Nature 2005, 433, 807. [Google Scholar] [CrossRef]
- Tuomainen, U.; Candolin, U. Behavioural responses to human-induced environmental change. Biol. Rev. 2011, 86, 640–657. [Google Scholar] [CrossRef]
- Kalcher-Sommersguter, E.; Franz-Schaider, C.; Crailsheim, K. Social competence of adult Chimpanzees (Pan troglodytes) with severe deprivation history: A relational approach. Int. J. Comp. Psychol. 2013, 26, 135–157. [Google Scholar] [CrossRef]
- Tung, J.; Archie, E.A.; Altmann, J.; Alberts, S. Cumulative early life adversity predicts longevity in wild baboons. Nat. Commun. 2016, 7, 11181. [Google Scholar] [CrossRef] [Green Version]
- Taborsky, B. Mothers determine offspring size in response to own juvenile growth conditions. Biol. Lett. 2006, 2, 225–228. [Google Scholar] [CrossRef]
- Zipple, M.N.; Archie, E.A.; Tung, J.; Altmann, J.; Alberts, S.C. Intergenerational effects of early adversity on survival in wild baboons. eLife 2019, 8, e47433. [Google Scholar] [CrossRef] [PubMed]
- Helle, H.; Koskela, W.; Mappes, T. Life in varying environments: Experimental evidence for delayed effects of juvenile environment on adult life history. J. Anim. Ecol. 2012, 81, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Burton, T.; Metcalfe, N.B. Can environmental conditions experienced in early life influence future generations? Proc. R. Soc. B 2014, 281, 20140311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garaï, M.E.; Slotow, R.; Carr, R.; Reilly, B. Elephant reintroductions to small fences reserves in South Africa. Pachyderm 2004, 37, 28–36. [Google Scholar]
- Bradshaw, G.A.; Schore, A.N. How elephants are opening doors: Developmental neuroethology, attachment and social context. Ethology 2007, 113, 426–436. [Google Scholar] [CrossRef] [Green Version]
- Pretorius, Y.; Garaï, M.E.; Bates, L.A. The status of African elephant Loxodonta africana populations in South Africa. Oryx 2019, 53, 757–763. [Google Scholar] [CrossRef]
- EMOA, Elephant Management & Owners Association; (Vaalwater, South Africa). Unpublished database. 2001.
- National Norms & Standards for the Management of Elephants. Government Gazette. 2018; 641, No. 42015. Available online: https://www.environment.gov.za/sites/default/files/gazetted_notices/nemba_elephantsinsa_g30833gon251.pdf# (accessed on 28 September 2020).
- Van der Water, A.; Garaï, M.E.; Henley, M.; Bates, L.; di Minin, E.; Slotow, R. Assessing the consequences of elephant management interventions on animal well-being, human well-being and environmental health. In prep.
- Parker, J.M.; Webb, C.T.; Daballen, D.; Goldenberg, S.G.; Lepieri, J.; Letitiya, D.; Lolchuragi, D.; Leadismo, C.; Douglas-Hamilton, I.; Wittemyer, G. Poaching of African elephants indirectly decreases population growth through lowered orphan survival. Curr. Biol. 2021, 31, 4156–4162. [Google Scholar] [CrossRef]
- Goldenberg, S.Z.; Wittemyer, G. Orphaning and natal group dispersal are associated with social costs in female elephants. Anim. Behav. 2018, 143, 1–8. [Google Scholar] [CrossRef]
- Belsky, J.; Ruttle, P.L.; Boyce, T.; Armstrong, J.M.; Essex, M.J. Early adversity, elevated stress physiology, accelerated sexual maturation and poor health in females. Dev. Psychol. 2015, 51, 816–822. [Google Scholar] [CrossRef]
- Maestripieri, D. Effects of early experience on female behavioural and reproductive development in rhesus macaques. Proc. R. Soc. B 2005, 272, 1243–1248. [Google Scholar] [CrossRef] [Green Version]
- Cameron, N.M. Maternal programming of reproductive function and behaviour in female rats. Front. Evol. Neurosci. 2011, 3, 10. [Google Scholar] [CrossRef]
- Rickard, I.J.; Frankenhuis, W.E.; Nettle, D. Why are childhood family factors associated with timing of maturation? A role for internal prediction. Perspect. Psychol. Sci. 2014, 9, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Garaï, M.E. The Development of Social Behaviour in Translocated Juvenile African Elephants Loxodonta africana (Blumenbach). Ph.D Thesis, University of Pretoria, Pretoria, South Africa, 1997. [Google Scholar]
- Van Os, R.; Vos, S. The impact of the African elephant (Loxodonta africana) on the vegetation of Pongola Game Reserve in South Africa. In Animal Management; Van Hall Institute: Leeuwarden, The Netherlands, 2000. [Google Scholar]
- Thornley, R.; Spencer, M.; Zitzer, H.R.; Parr, C.L. Woody vegetation damage by the African elephant during severe drought at Pongola Game Reserve, South Africa. Afr. J. Ecol. 2020, 58, 658–673. [Google Scholar] [CrossRef]
- Zitzer, H.R.; Boult, V.L. Vasectomies of male African elephants as a population management tool: A case study. Bothalia 2018, 48, a2313. [Google Scholar] [CrossRef]
- Altmann, J. Observational study of behaviour: Sampling methods. Behaviour 1974, 49, 227–267. [Google Scholar] [CrossRef] [Green Version]
- Mutinda, H.; Poole, J.H.; Moss, C.J. Decision making and leadership in using the ecosystem. In The Amboseli Elephants: A Long-Term Perspective on a Long-Lived Mammal; Moss, C.J., Croze, H., Lee, P.C., Eds.; University of Chicago Press: Chicago, IL, USA, 2011; pp. 246–259. [Google Scholar]
- Moss, C.J.; Lee, P.C. Female social dynamics: Fidelity and flexibility. In The Amboseli Elephants: A Long-Term Perspective on a Long-Lived Mammal; Moss, C.J., Croze, H., Lee, P.C., Eds.; University of Chicago Press: Chicago, IL, USA, 2011; pp. 205–223. [Google Scholar]
- Moss, C.J.; Lee, P.C. Female reproductive strategies: Individual life histories. In The Amboseli Elephants: A Long-Term Perspective on a Long-Lived Mammal; Moss, C.J., Croze, H., Lee, P.C., Eds.; University of Chicago Press: Chicago, IL, USA, 2011; pp. 187–204. [Google Scholar]
- Boult, V.L.; Quaife, T.; Fishlock, V.; Moss, C.J.; Lee, P.C.; Sibly, R.M. Individual-based modelling of elephant population dynamics using remote sensing to estimate food availability. Ecol. Model. 2018, 387, 187–195. [Google Scholar] [CrossRef]
- Lee, P.C.; Moss, C.J. Early maternal investment in male and female African elephant calves. Behav. Ecol. Sociobiol. 1986, 18, 353–361. [Google Scholar] [CrossRef]
- Whiten, A. A second inheritance system: The extension of biology through culture. Interface Focus 2017, 7, 20160142. [Google Scholar] [CrossRef] [Green Version]
- Goldenberg, S.Z.; Douglas-Hamilton, I.; Wittemyer, G. Inter-generational change in African elephant range use is associated with poaching risk, primary productivity and adult mortality. Proc. R. Soc. B 2018, 285, 20180286. [Google Scholar] [CrossRef]
- Ahem, T.H.; Young, L.Y. The impact of early life family structure on adult social attachment, alloparental behaviour, and the neuropeptide systems regulating affiliative behaviours in the monogamous prairie vole (Microtus ochrogaster). Front. Behav. Neurosci. 2009, 3, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Diorio, J.; Day, J.C.; Francis, D.D.; Meaney, M.J. Maternal care, hippocampal synaptogenesis and cognitive development in rats. Nat. Neurosci. 2000, 3, 799–806. [Google Scholar] [CrossRef]
- Aureli, F.; Schaffner, C.M.; Boesch, C.; Bearder, S.K.; Call, J.; Chapman, C.A.; Connor, R.; Di Fiore, A.; Dunbar, R.I.M.; Henzi, S.P.; et al. Fission-Fusion Dynamics: New Research Frameworks. Curr. Anthropol. 2008, 49, 627–654. [Google Scholar] [CrossRef]
- Kappeler, P.M.; van Schaik, C.P. Evolution of Primate Social Systems. Int. J. Primat. 2002, 23, 707–740. [Google Scholar] [CrossRef]
- Emlen, S.T. An evolutionary theory of the family. Proc. Natl. Acad. Sci. USA 1995, 92, 8092–8099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, P.C.; Moss, C.J. Wild Female African Elephants (Loxodonta africana) Exhibit Personality Traits of Leadership and Social Integration. J. Comp. Psychol. 2012, 126, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Fishlock, V.; Lee, P.C. Forest elephants: Fission-fusion and social arenas. Anim. Behav. 2013, 2, 357–363. [Google Scholar] [CrossRef]
- Beckoff, M. Mammalian Dispersal and the Ontogeny of Individual Behavioural Phenotypes. Am. Nat. 1977, 111, 715–732. [Google Scholar] [CrossRef]
- Blumenstein, D.T.; Wey, T.W.; Tang, K. A test of the social cohesion hypothesis: Interactive female marmots remain at home. Proc. R. Soc. B 2009, 276, 3007–3012. [Google Scholar] [CrossRef] [Green Version]
- Mellor, D.J. Positive animal welfare states and encouraging environment-focused and animal-to-animal interactive behaviours. N. Z. Vet. J. 2015, 63, 9–16. [Google Scholar] [CrossRef]
- Smith, J.E.; Ortiz, C.A.; Buhbe, M.T.; van Vugt, M. Obstacles and opportunities for female leadership in mammalian societies: A comparative perspective. Leadersh. Q. 2020, 31, 101267. [Google Scholar] [CrossRef]
- Plotnik, J.M.; de Waal, F.B.M. Asian elephants (Elephas maximus) reassure others in distress. PeerJ 2014, 2, e278. [Google Scholar] [CrossRef] [PubMed]
- Buss, I.O.; Smith, N.S. Observations on reproduction and breeding behaviour of the African elephant. J. Wildl. Manag. 1966, 30, 375–388. [Google Scholar] [CrossRef]
- Laws, R.M. Aspects of reproduction in the African elephant, Loxodonta africana. J. Reprod. Fertil. 1969, 6, 193–217. [Google Scholar]
- Hanks, J. Reproduction of elephants, Loxodonta africana, in the Luangwa Valley, Zambia. J. Reprod. Fert. 1972, 30, 13–26. [Google Scholar] [CrossRef] [Green Version]
- Moss, C.J. Elephant Memories; Elm Tree Books: London, UK, 1988; pp. 119–143. [Google Scholar]
- Kuiper, T.R.; Druce, D.J.; Druce, H.C. Demography and social dynamics of an African elephant population 35 years after reintroduction as juveniles. J. Appl. Ecol. 2018, 55, 2898–2907. [Google Scholar] [CrossRef]
- Whyte, I.J. Conservation Management of Kruger National Park Elephant Population. Ph.D. Thesis, Faculty of Natural and Agricultural Sciences, University of Pretoria, Pretoria, South Africa, 2001. [Google Scholar]
- Lee, P.C.; Moss, C.J.; Njiraini, N.; Poole, J.H.; Sayialel, K.; Fishlock, V.L. Cohort consequences of drought and family disruption for male and female African elephants. Behav. Ecol. 2021, 33, 408–418. [Google Scholar] [CrossRef]
- Moss, C.H. The demography of an African elephant (Loxodonta africana ) population in Amboseli, Kenya. J. Zool. Lond. 2001, 255, 145–156. [Google Scholar] [CrossRef]
- Whitehouse, A.M.; Hall-Martin, A. Elephants in Addo Elephant National Park, South Africa: Reconstruction of the population’s history. Oryx 2000, 34, 46–55. [Google Scholar] [CrossRef]
- Oerke, A.-K. Monitoring of sexual maturity of female elephants in EEP populations. EAZA News 2004, 47, 34–35. Available online: https://www.yumpu.com/en/document/read/4892997/from-the-african-elephant-eep-european-association-of-zoos-and- (accessed on 8 June 2022).
- Reichert, S.; Berger, V.; Jackson, J.; Chapman, S.N.; Htut, W.; Mar, K.U.; Lummaa, V. Maternal age at birth shapes offspring life-history trajectory across generations in long-lived Asian elephants. J. Anim. Ecol. 2019, 89, 996–1007. [Google Scholar] [CrossRef] [PubMed]
- Nettle, D.; Frankenhuis, W.E.; Rickard, I.J. The evolution of predictive adaptive responses in human life history. Proc. R. Soc. B 2013, 280, 1343. [Google Scholar] [CrossRef] [PubMed]
- Hein, S.; Jacob, J. Recovery of small rodent populations after population collapse. Wildl. Res. 2015, 42, 108–118. [Google Scholar] [CrossRef]
- Poole, J.H.; Granli, P. The Gorongosa elephants through war and recovery: Tusklessness, population size, structure and reproductive parameters. Pachyderm 2022. In print. [Google Scholar]
- Clutton-Brock, T.H.; Iason, G.R. Sex variation in mammals. Q. Rev. Biol. 1986, 61, 339–374. [Google Scholar] [CrossRef]
- Trivers, R.L.; Willard, D.E. Natural selection of parental ability to vary the rex ratio of offspring. Science 1973, 179, 90–91. [Google Scholar] [CrossRef] [Green Version]
- Conradt, L.; Roper, T.J. Group Decision-making in animals. Nature 2003, 421, 155–158. [Google Scholar] [CrossRef]
- Kerth, G.; Ebert, C.; Schmidtke, C. Group decision making in fission–fusion societies: Evidence from two-field experiments in Bechstein’s bats. Proc. R. Soc. B 2006, 273, 2785–2790. [Google Scholar] [CrossRef]
- Archie, E.A.; Morrison, T.A.; Foley, C.A.H.; Moss, C.J.; Alberts, S.C. Dominance rank relationships among wild female African elephants, Loxodonta Africana. Anim. Behav. 2006, 71, 117–127. [Google Scholar] [CrossRef]
- Meaney, M.J. Maternal care, gene expression, and the transmission of individual differences in stress reactivity across generations. Annu. Rev. Neurosci. 2001, 24, 1161–1192. [Google Scholar] [CrossRef]
- Shea, N.; Pen, I.; Uller, T. Three epigenetic information channels and their different roles in evolution. J. Evol. Biol. 2011, 24, 1178–1187. [Google Scholar] [CrossRef] [Green Version]
- Jensen, P. Transgenerational epigenetic effects on animal behaviour. Prog. Biophys. Mol. Biol. 2013, 113, 447–454. [Google Scholar] [CrossRef] [Green Version]
- Jablonka, E.; Raz, G. Transgenerational epigenetic inheritance: Prevalence, mechanisms, and implications for the study of heredity and evolution. Q. Rev. Biol. 2009, 84, 131–176. [Google Scholar] [CrossRef] [Green Version]
- Ledón-Rettig, C.C.; Richards, C.L.; Martin, L.M. Epigenetics for behavioural ecologists. Behav. Ecol. 2013, 24, 311–324. [Google Scholar] [CrossRef] [Green Version]
- Garaï, M.E.; (Elephant Reintegration Trust, Port Alfred, South Africa); Pretorius, Y.; (Mammal Research Institute, University of Pretoria, Pretoria, South Africa). Assessment of Mabula Private Game Reserve’s Elephants. Unpublished Report for MPGR management. 2011. [Google Scholar]
- Brakes, P.; Dall, S.R.X.; Aplin, L.M.; Bearhop, S.; Carroll, E.L.; Ciucci, P.; Fishlock, V.; Ford, J.K.B.; Garland, E.C.; Keith, S.A.; et al. Animal cultures matter for conservation. Science 2019, 363, 1032–1034. [Google Scholar] [CrossRef]
- Schuppli, C.; van Schaik, C.P. Animal cultures: How we’ve only seen the tip of the iceberg. Evol. Hum. Sci. 2019, 1, e2. [Google Scholar] [CrossRef] [Green Version]
- Whiten, A.; Caldwell, C.A.; Mesoudi, A. Cultural diffusion in humans and other animals. Curr. Opin. Psychol. 2016, 8, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, H. Conserving and managing animals that learn socially and share cultures. Learn. Behav. 2010, 38, 329–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantor, M.; Shoemaker, L.G.; Cabral, R.B.; Flores, C.O.; Varga, M.; Whitehead, H. Multilevel animal societies can emerge from cultural transmission. Nat. Commun. 2015, 6, 8091. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Origin | Numbers |
|---|---|---|
| 9 June 1997 | Translocated from KNP as an intact family group (A family). | Eight made up of:
|
| 13 June 1997 | Translocated from KNP as an intact family group (B family). | Nine made up of:
|
| July 1998 | Translocated from KNP. | Three adult males |
| March 2000 | KNP culling orphans initially translocated to a nearby reserve but subsequently moved into PGR (the Orphans). | Five made up of:
|
| August 2001 | Initially translocated from KNP to a nearby reserve, but subsequently moved into PGR. | Three adult males |
| Group | Age Classification | Numbers |
|---|---|---|
| A&B herd | Adult females | 17 |
| Sub-adult females | 5 | |
| Sub-adult males | 4 | |
| Juveniles | 21 | |
| Calves | 12 | |
| Orphans | Adult females | 5 |
| Sub-adult females | 0 | |
| Sub-adult males | 5 | |
| Juveniles | 11 | |
| Calves | 1 | |
| Adult males | Independent bulls | 13 |
| Total | 94 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garaï, M.E.; Boult, V.L.; Zitzer, H.R. Identifying the Effects of Social Disruption through Translocation on African Elephants (Loxodonta africana), with Specifics on the Social and Ecological Impacts of Orphaning. Animals 2023, 13, 483. https://doi.org/10.3390/ani13030483
Garaï ME, Boult VL, Zitzer HR. Identifying the Effects of Social Disruption through Translocation on African Elephants (Loxodonta africana), with Specifics on the Social and Ecological Impacts of Orphaning. Animals. 2023; 13(3):483. https://doi.org/10.3390/ani13030483
Chicago/Turabian StyleGaraï, Marion E., Victoria L. Boult, and Heike R. Zitzer. 2023. "Identifying the Effects of Social Disruption through Translocation on African Elephants (Loxodonta africana), with Specifics on the Social and Ecological Impacts of Orphaning" Animals 13, no. 3: 483. https://doi.org/10.3390/ani13030483