
Listening to Their Nights: Sleep Disruptions in Captive Housed Chimpanzees Affect Their Daytime Behavior


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sample
2.2. Nocturnal Sound Recording
2.3. Measuring Abiotic Factors of Night Quarters
2.4. Daytime Behavioral Sampling
2.5. Statistical Analysis
2.5.1. Correlation between Abiotic Factors and Nocturnal Events
2.5.2. Linear Mixed Models
3. Results
3.1. Correlation between Abiotic Factors and Nocturnal Events
3.2. Linear Mixed Models
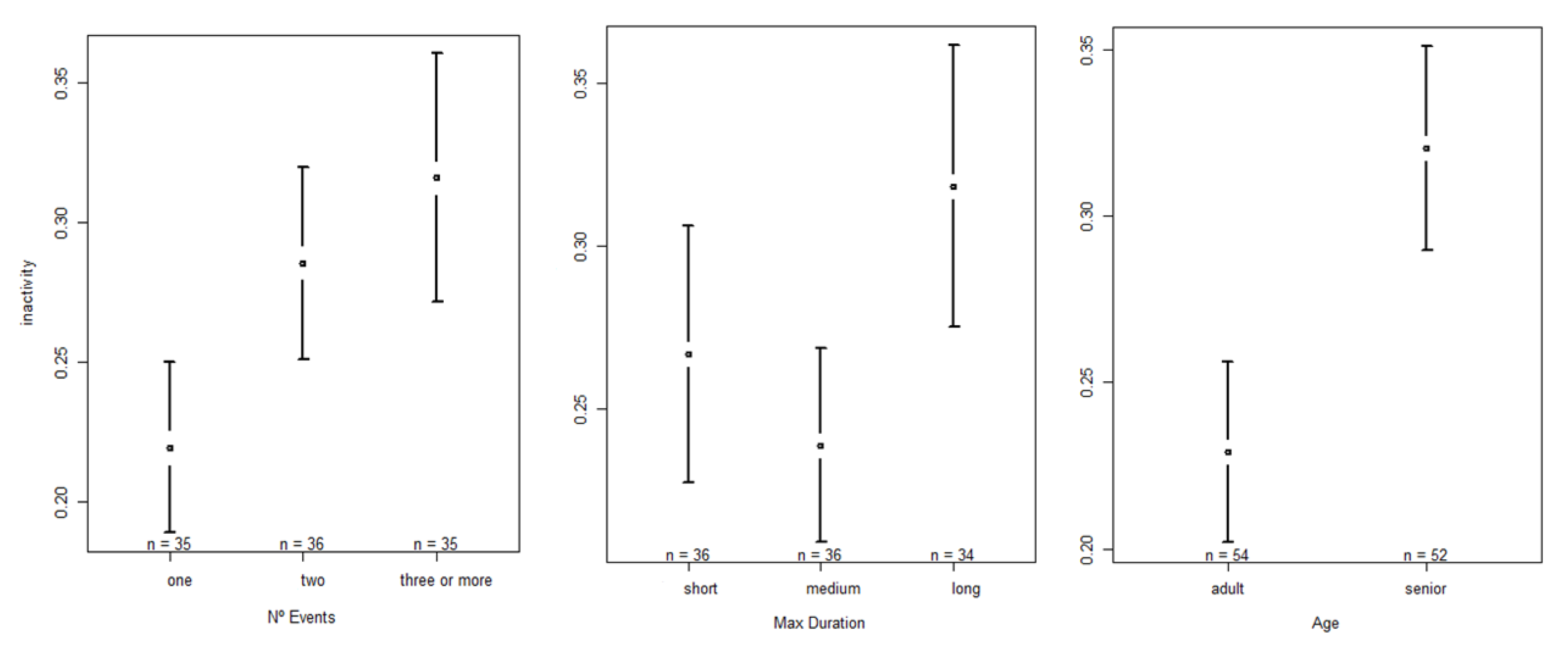
3.2.1. Impact on the Occurrence of Inactivity
3.2.2. Impact on the Occurrence of Self-Directed and Abnormal Behaviors
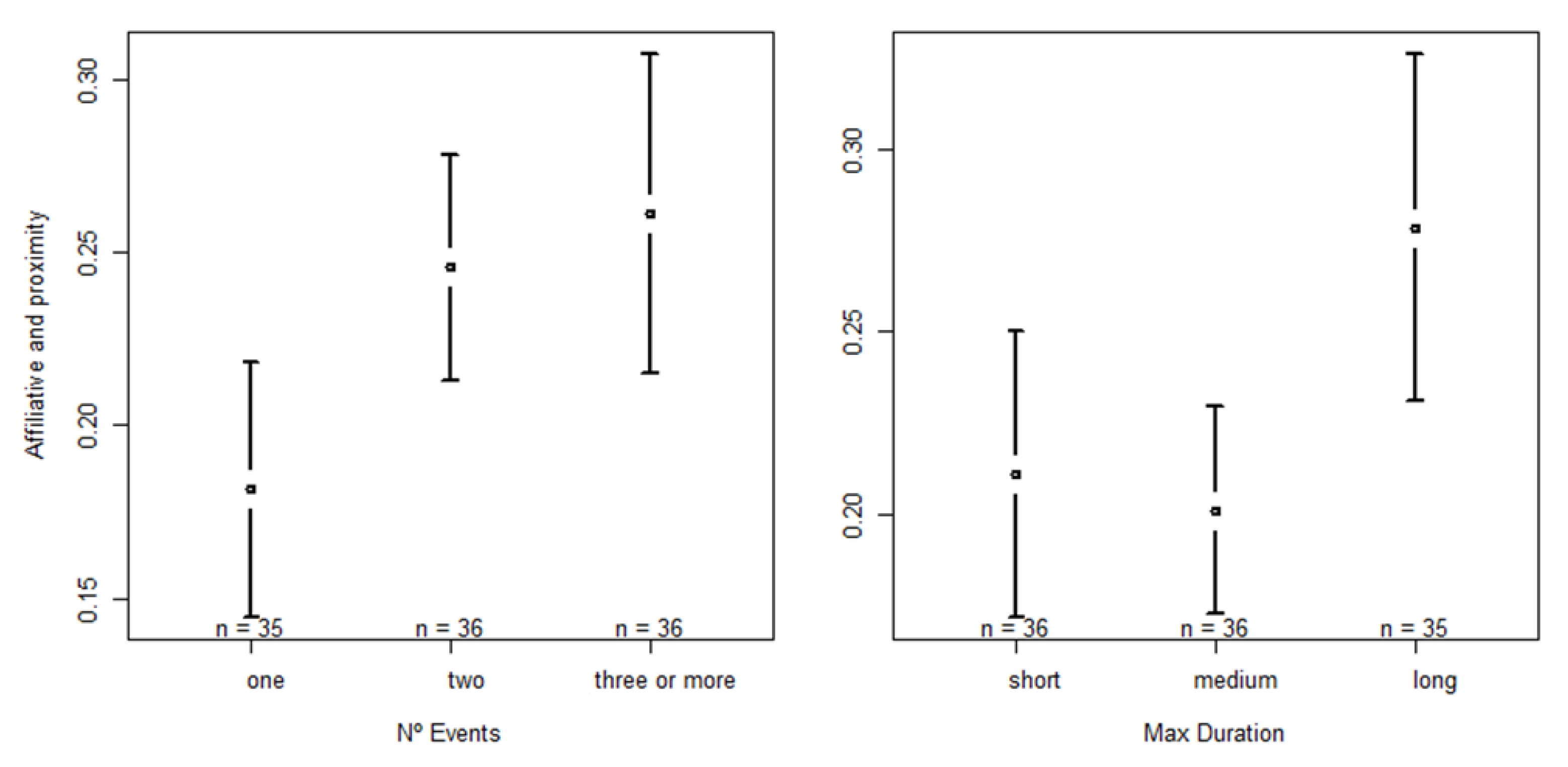
3.2.3. Impact on the Occurrence of Social Proximity and Affiliative Behaviors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geiger, S.D.; Sabanayagam, C.; Shankar, A. The Relationship between Insufficient Sleep and Self-Rated Health in a Nationally Representative Sample. J. Environ. Public Health 2012, 2012, e518263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, M.E.; Vaughn, B.V. Poor Sleep Challenging the Health of a Nation. Neurodiagn. J. 2012, 52, 233–249. [Google Scholar] [PubMed]
- Buxton, O.M.; Marcelli, E. Short and Long Sleep Are Positively Associated with Obesity, Diabetes, Hypertension, and Cardiovascular Disease among Adults in the United States. Soc. Sci. Med. 2010, 71, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Gangwisch, J.E.; Heymsfield, S.B.; Boden-Albala, B.; Buijs, R.M.; Kreier, F.; Pickering, T.G.; Rundle, A.G.; Zammit, G.K.; Malaspina, D. Sleep Duration as a Risk Factor for Diabetes Incidence in a Large US Sample. Sleep 2007, 30, 1667–1673. [Google Scholar] [CrossRef]
- Anderson, J.R. Sleep, Sleeping Sites, and Sleep-Related Activities: Awakening to Their Significance. Am. J. Primatol. 1998, 46, 63–75. [Google Scholar] [CrossRef]
- Anderson, J.R. Sleep-Related Behavioural Adaptations in Free-Ranging Anthropoid Primates. Sleep Med. Rev. 2000, 4, 355–373. [Google Scholar] [CrossRef]
- Diekelmann, S.; Born, J. The Memory Function of Sleep. Nat. Rev. Neurosci. 2010, 11, 114–126. [Google Scholar] [CrossRef]
- Siegel, J.M. Clues to the Functions of Mammalian Sleep. Nature 2005, 437, 1264–1271. [Google Scholar] [CrossRef]
- Krief, S.; Cibot, M.; Bortolamiol, S.; Seguya, A.; Krief, J.-M.; Masi, S. Wild Chimpanzees on the Edge: Nocturnal Activities in Croplands. PLoS ONE 2014, 9, e109925. [Google Scholar] [CrossRef] [Green Version]
- Pruetz, J.D. Nocturnal Behavior by a Diurnal Ape, the West African Chimpanzee (Pan Troglodytes Verus), in a Savanna Environment at Fongoli, Senegal. Am. J. Phys. Anthropol. 2018, 166, 541–548. [Google Scholar] [CrossRef]
- Banks, S.; Dinges, D.F. Behavioral and Physiological Consequences of Sleep Restriction. J. Clin. Sleep Med. 2007, 03, 519–528. [Google Scholar] [CrossRef] [Green Version]
- Brando, S.; Buchanan-Smith, H.M. The 24/7 Approach to Promoting Optimal Welfare for Captive Wild Animals. Behav. Process. 2018, 156, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Tobler, I. Is Sleep Fundamentally Different between Mammalian Species? Behav. Brain Res. 1995, 69, 35–41. [Google Scholar] [CrossRef]
- McNamara, P.; Barton, R.A.; Nunn, C.L. Evolution of Sleep: Phylogenetic and Functional Perspectives; Cambridge University Press: Cambridge, UK, 2010; ISBN 978-0-521-89497-5. [Google Scholar]
- Videan, E.N. Sleep in Captive Chimpanzee (Pan Troglodytes): The Effects of Individual and Environmental Factors on Sleep Duration and Quality. Behav. Brain Res. 2006, 169, 187–192. [Google Scholar] [CrossRef]
- Havercamp, K.; Morimura, N.; Hirata, S. Sleep Patterns of Aging Chimpanzees (Pan Troglodytes). Int. J. Primatol. 2021, 42, 89–104. [Google Scholar] [CrossRef]
- Goodall, J.M. Nest Building Behavior in the Free Ranging Chimpanzee. Ann. New York Acad. Sci. 1962, 102, 455–467. [Google Scholar] [CrossRef] [PubMed]
- Fruth, B.; Tagg, N.; Stewart, F. Sleep and Nesting Behavior in Primates: A Review. Am. J. Phys. Anthropol. 2018, 166, 499–509. [Google Scholar] [CrossRef]
- Van Lawick-Goodall, J. The Behaviour of Free-Living Chimpanzees in the Gombe Stream Reserve. Anim. Behav. Monogr. 1968, 1, 161–311. [Google Scholar] [CrossRef]
- Riss, D.; Goodall, J. Sleeping Behavior and Associations in a Group of Captive Chimpanzees. Folia Primatol. 1976, 25, 1–11. [Google Scholar] [CrossRef]
- Zamma, K. What Makes Wild Chimpanzees Wake up at Night? Primates J. Primatol. 2014, 55, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Piel, A.K. Temporal Patterns of Chimpanzee Loud Calls in the Issa Valley, Tanzania: Evidence of Nocturnal Acoustic Behavior in Wild Chimpanzees. Am. J. Phys. Anthropol. 2018, 166, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Tagg, N.; McCarthy, M.; Dieguez, P.; Bocksberger, G.; Willie, J.; Mundry, R.; Stewart, F.; Arandjelovic, M.; Widness, J.; Landsmann, A.; et al. Nocturnal Activity in Wild Chimpanzees (Pan Troglodytes): Evidence for Flexible Sleeping Patterns and Insights into Human Evolution. Am. J. Phys. Anthropol. 2018, 166, 510–529. [Google Scholar] [CrossRef] [PubMed]
- Pruetz, J.; Bertolani, P. Chimpanzee (Pan Troglodytes Verus) Behavioral Responses to Stresses Associated with Living in a Savannah-Mosaic Environment: Implications for Hominin Adaptations to Open Habitats. PaleoAnthropology 2009, 2009, 252–262. [Google Scholar] [CrossRef]
- Baker, K.C.; Aureli, F. The Neighbor Effect: Other Groups Influence Intragroup Agonistic Behavior in Captive Chimpanzees. Am. J. Primatol. 1996, 40, 283–291. [Google Scholar] [CrossRef]
- Videan, E.N.; Fritz, J.; Schwandt, M.; Howell, S. Neighbor Effect: Evidence of Affiliative and Agonistic Social Contagion in Captive Chimpanzees (Pan Troglodytes). Am. J. Primatol. 2005, 66, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Landolt, H.-P.; Holst, S.C.; Sousek, A. Effects of Acute and Chronic Sleep Deprivation. In Sleep Medicine Textbook; European Sleep Research Society: Regensburg, Germany, 2014; pp. 49–62. ISBN 978-1-119-03893-1. [Google Scholar]
- McKenna, J.T.; Tartar, J.L.; Ward, C.P.; Thakkar, M.M.; Cordeira, J.W.; McCarley, R.W.; Strecker, R.E. Sleep Fragmentation Elevates Behavioral, Electrographic and Neurochemical Measures of Sleepiness. Neuroscience 2007, 146, 1462–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tougeron, K.; Abram, P.K. An Ecological Perspective on Sleep Disruption. Am. Nat. 2017, 190, E55–E66. [Google Scholar] [CrossRef]
- Schork, I.G.; Manzo, I.A.; De Oliveira, M.R.B.; da Costa, F.V.; Young, R.J.; de Azevedo, C.S. The Cyclic Interaction between Daytime Behavior and the Sleep Behavior of Laboratory Dogs. Sci. Rep. 2022, 12, 478. [Google Scholar] [CrossRef]
- Kamphuis, J.; Meerlo, P.; Koolhaas, J.M.; Lancel, M. Poor Sleep as a Potential Causal Factor in Aggression and Violence. Sleep Med. 2012, 13, 327–334. [Google Scholar] [CrossRef]
- Walker, M.P. The Role of Sleep in Cognition and Emotion. Ann. New York Acad. Sci. 2009, 1156, 168–197. [Google Scholar] [CrossRef]
- Willner, P.; Muscat, R.; Papp, M. Chronic Mild Stress-Induced Anhedonia: A Realistic Animal Model of Depression. Neurosci. Biobehav. Rev. 1992, 16, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Samson, D.R.; Vining, A.; Nunn, C.L. Sleep Influences Cognitive Performance in Lemurs. Anim. Cogn. 2019, 22, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Cheeta, S.; Ruigt, G.; van Proosdij, J.; Willner, P. Changes in Sleep Architecture Following Chronic Mild Stress. Biol. Psychiatry 1997, 41, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Abou-Ismail, U.A.; Burman, O.H.P.; Nicol, C.J.; Mendl, M. Let Sleeping Rats Lie: Does the Timing of Husbandry Procedures Affect Laboratory Rat Behaviour, Physiology and Welfare? Appl. Anim. Behav. Sci. 2008, 111, 329–341. [Google Scholar] [CrossRef]
- Ruckerbusch, Y. The Hypnogram as an Index of Adaptation of Farm Animals to Changes in Their Environment. Appl. Anim. Ethol. 1975, 2, 3–18. [Google Scholar] [CrossRef]
- Hokkanen, A.-H.; Hänninen, L.; Tiusanen, J.; Pastell, M. Predicting Sleep and Lying Time of Calves with a Support Vector Machine Classifier Using Accelerometer Data. Appl. Anim. Behav. Sci. 2011, 134, 10–15. [Google Scholar] [CrossRef]
- Meagher, R.K.; Campbell, D.L.M.; Dallaire, J.A.; Díez-León, M.; Palme, R.; Mason, G.J. Sleeping Tight or Hiding in Fright? The Welfare Implications of Different Subtypes of Inactivity in Mink. Appl. Anim. Behav. Sci. 2013, 144, 138–146. [Google Scholar] [CrossRef] [Green Version]
- Owczarczak-Garstecka, S.C.; Burman, O.H.P. Can Sleep and Resting Behaviours Be Used as Indicators of Welfare in Shelter Dogs (Canis Lupus Familiaris)? PLoS ONE 2016, 11, e0163620. [Google Scholar] [CrossRef] [Green Version]
- Holdgate, M.R.; Meehan, C.L.; Hogan, J.N.; Miller, L.J.; Rushen, J.; de Passillé, A.M.; Soltis, J.; Andrews, J.; Shepherdson, D.J. Recumbence Behavior in Zoo Elephants: Determination of Patterns and Frequency of Recumbent Rest and Associated Environmental and Social Factors. PLoS ONE 2016, 11, e0153301. [Google Scholar] [CrossRef] [Green Version]
- Barrett, C.E.; Noble, P.; Hanson, E.; Pine, D.S.; Winslow, J.T.; Nelson, E.E. Early Adverse Rearing Experiences Alter Sleep–Wake Patterns and Plasma Cortisol Levels in Juvenile Rhesus Monkeys. Psychoneuroendocrinology 2009, 34, 1029–1040. [Google Scholar] [CrossRef] [Green Version]
- Vining, A.Q.; Nunn, C.L.; Samson, D.R. Enriched Sleep Environments Lengthen Lemur Sleep Duration. PLoS ONE 2021, 16, e0253251. [Google Scholar] [CrossRef] [PubMed]
- Morimura, N.; Fujisawa, M.; Mori, Y.; Teramoto, M. Environmental Influences on Sleep Behavior in Captive Male Chimpanzees (Pan Troglodytes). Int. J. Primatol. 2012, 33, 822–829. [Google Scholar] [CrossRef] [Green Version]
- Zamansky, A.; Sinitca, A.M.; Kaplun, D.I.; Plazner, M.; Schork, I.G.; Young, R.J.; de Azevedo, C.S. Analysis of Dogs’ Sleep Patterns Using Convolutional Neural Networks. In Artificial Neural Networks and Machine Learning—ICANN 2019: Image Processing; Tetko, I.V., Kůrková, V., Karpov, P., Theis, F., Eds.; Springer International Publishing: Munich, Germany, 2019; Volume 11729, pp. 472–483. [Google Scholar]
- Melvin, E.; Samson, D.; Nunn, C.L. Eulerian Videography Technology Improves Classification of Sleep Architecture in Primates. Primates 2019, 60, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Mielke, A.; Zuberbühler, K. A Method for Automated Individual, Species and Call Type Recognition in Free-Ranging Animals. Anim. Behav. 2013, 86, 475–482. [Google Scholar] [CrossRef]
- Kalan, A.K.; Piel, A.K.; Mundry, R.; Wittig, R.M.; Boesch, C.; Kühl, H.S. Passive Acoustic Monitoring Reveals Group Ranging and Territory Use: A Case Study of Wild Chimpanzees (Pan Troglodytes). Front. Zool. 2016, 13, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astaras, C.; Linder, J.; Wrege, P.; Orume, R.; Macdonald, D. Passive Acoustic Monitoring as a Law Enforcement Tool for Afrotropical Rainforests. Front. Ecol. Environ. 2017, 15, 233–234. [Google Scholar] [CrossRef]
- Wark, J.; Cronin, K.; Niemann, T.; Shender, M.; Horrigan, A.; Kao, A.; Ross, M. Monitoring the Behavior and Habitat Use of Animals to Enhance Welfare Using the ZooMonitor App. Anim. Behav. Cogn. 2019, 6, 158–167. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; SAGE Publications: New York, NY, USA, 2018; ISBN 978-1-5443-3648-0. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org (accessed on 14 January 2023).
- Perez-Olmos, I.; Ibanez-Pinilla, M. Night Shifts, Sleep Deprivation, and Attention Performance in Medical Students. Int. J. Med. Educ. 2014, 5, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Scheltens, P.; Visscher, F.; Van Keimpema, A.R.; Lindeboom, J.; Taphoorn, M.J.; Wolters, E.C. Sleep Apnea Syndrome Presenting with Cognitive Impairment. Neurology 1991, 41, 155–156. [Google Scholar] [CrossRef]
- de Waal, F.B.M.; van Roosmalen, A. Reconciliation and Consolation among Chimpanzees. Behav. Ecol. Sociobiol. 1979, 5, 55–66. [Google Scholar] [CrossRef]
- Fraser, O.N.; Aureli, F. Reconciliation, Consolation and Postconflict Behavioral Specificity in Chimpanzees. Am. J. Primatol. 2008, 70, 1114–1123. [Google Scholar] [CrossRef] [PubMed]
- Webb, C.E.; Franks, B.; Romero, T.; Higgins, E.T.; de Waal, F.B.M. Individual Differences in Chimpanzee Reconciliation Relate to Social Switching Behaviour. Anim. Behav. 2014, 90, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Maestripieri, D.; Schino, G.; Aureli, F.; Troisi, A. A Modest Proposal: Displacement Activities as an Indicator of Emotions in Primates. Anim. Behav. 1992, 44, 967–979. [Google Scholar] [CrossRef]
- Leavens, D.A.; Aureli, F.; Hopkins, W.D.; Hyatt, C.W. Effects of Cognitive Challenge on Self-Directed Behaviors by Chimpanzees (Pan Troglodytes). Am. J. Primatol. 2001, 55, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aureli, F.; Cords, M.; van Schaik, C.P. Conflict Resolution Following Aggression in Gregarious Animals: A Predictive Framework. Anim. Behav. 2002, 64, 325–343. [Google Scholar] [CrossRef]
- Minkel, J.D.; McNealy, K.; Gianaros, P.J.; Drabant, E.M.; Gross, J.J.; Manuck, S.B.; Hariri, A.R. Sleep Quality and Neural Circuit Function Supporting Emotion Regulation. Biol. Mood Anxiety Disord. 2012, 2, 22. [Google Scholar] [CrossRef] [Green Version]
- Mauss, I.B.; Troy, A.S.; LeBourgeois, M.K. Poorer Sleep Quality Is Associated with Lower Emotion-Regulation Ability in a Laboratory Paradigm. Cogn. Emot. 2013, 27, 567–576. [Google Scholar] [CrossRef] [Green Version]
- O’Leary, K.; Bylsma, L.M.; Rottenberg, J. Why Might Poor Sleep Quality Lead to Depression? A Role for Emotion Regulation. Cogn. Emot. 2017, 31, 1698–1706. [Google Scholar] [CrossRef]
- Mason, G.; Rushen, J. Stereotypic Animal Behaviour: Fundamentals and Applications to Welfare; CABI: Wallingford, UK, 2008; ISBN 978-1-84593-465-1. [Google Scholar]
- Jacobson, S.L.; Ross, S.R.; Bloomsmith, M.A. Characterizing Abnormal Behavior in a Large Population of Zoo-Housed Chimpanzees: Prevalence and Potential Influencing Factors. PeerJ 2016, 4, e2225. [Google Scholar] [CrossRef] [Green Version]
- Birkett, L.P.; Newton-Fisher, N.E. How Abnormal Is the Behaviour of Captive, Zoo-Living Chimpanzees? PLoS ONE 2011, 6, e20101. [Google Scholar] [CrossRef] [PubMed]
- Goldsborough, Z.; Sterck, E.H.M.; de Waal, F.; Webb, C.E. Individual Variation in Chimpanzee (Pan Troglodytes) Repertoires of Abnormal Behaviour. Anim. Welf. 2022, 31, 125–135. [Google Scholar] [CrossRef]
- Neal Webb, S.J.; Hau, J.; Lambeth, S.P.; Schapiro, S.J. Differences in Behavior Between Elderly and Nonelderly Captive Chimpanzees and the Effects of the Social Environment. J. Am. Assoc. Lab. Anim. Sci. JAALAS 2019, 58, 783–789. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayuso, P.R.; Feliu, O.; Riba, D.; Crailsheim, D. Listening to Their Nights: Sleep Disruptions in Captive Housed Chimpanzees Affect Their Daytime Behavior. Animals 2023, 13, 696. https://doi.org/10.3390/ani13040696
Ayuso PR, Feliu O, Riba D, Crailsheim D. Listening to Their Nights: Sleep Disruptions in Captive Housed Chimpanzees Affect Their Daytime Behavior. Animals. 2023; 13(4):696. https://doi.org/10.3390/ani13040696
Chicago/Turabian StyleAyuso, Pablo R., Olga Feliu, David Riba, and Dietmar Crailsheim. 2023. "Listening to Their Nights: Sleep Disruptions in Captive Housed Chimpanzees Affect Their Daytime Behavior" Animals 13, no. 4: 696. https://doi.org/10.3390/ani13040696