
Identification of Candidate Genes and Functional Pathways Associated with Body Size Traits in Chinese Holstein Cattle Based on GWAS Analysis


 ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Phenotypic Data and DNA Samples Collection
2.3. Phenotypic and Genetic Parameters
2.4. Genotyping and Quality Control
2.5. Population Stratification
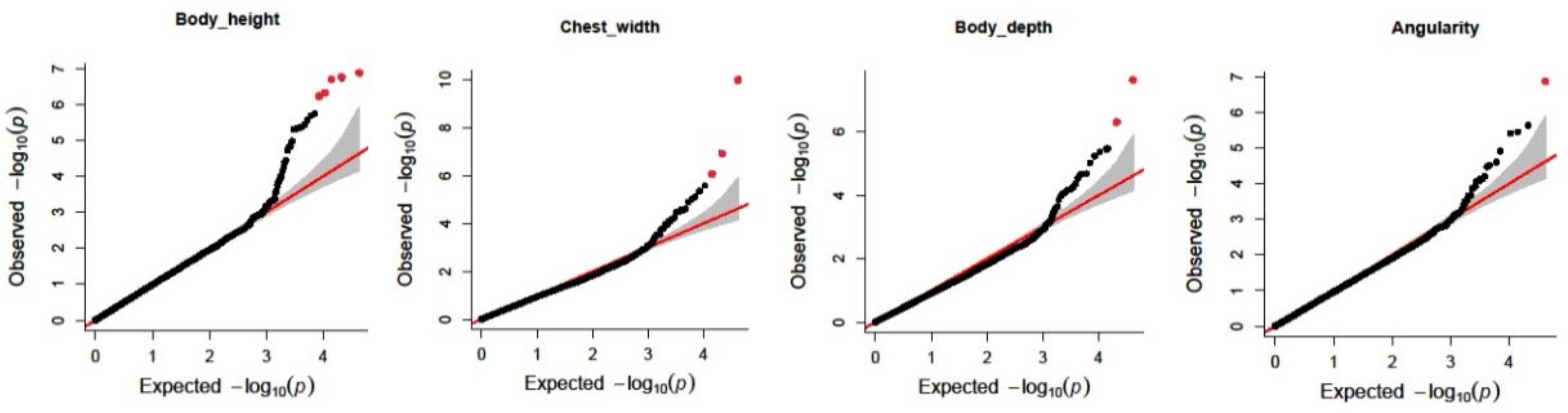
2.6. Genome-Wide Association Studies
2.7. Gene Identification and Functional Analysis
3. Results
3.1. Descriptive Statistics and Genetic Parameters Estimation of Body Size Traits
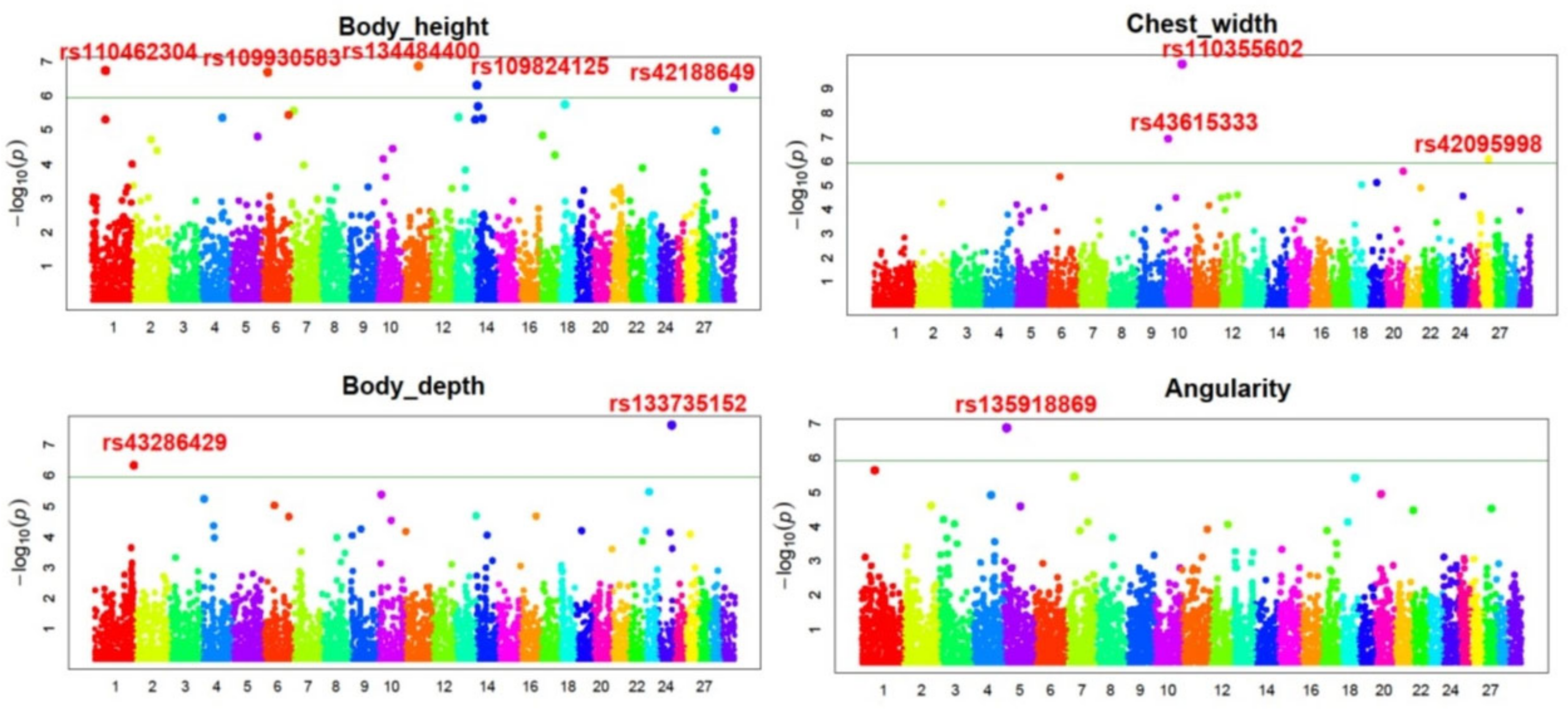
3.2. Candidate Genes Association
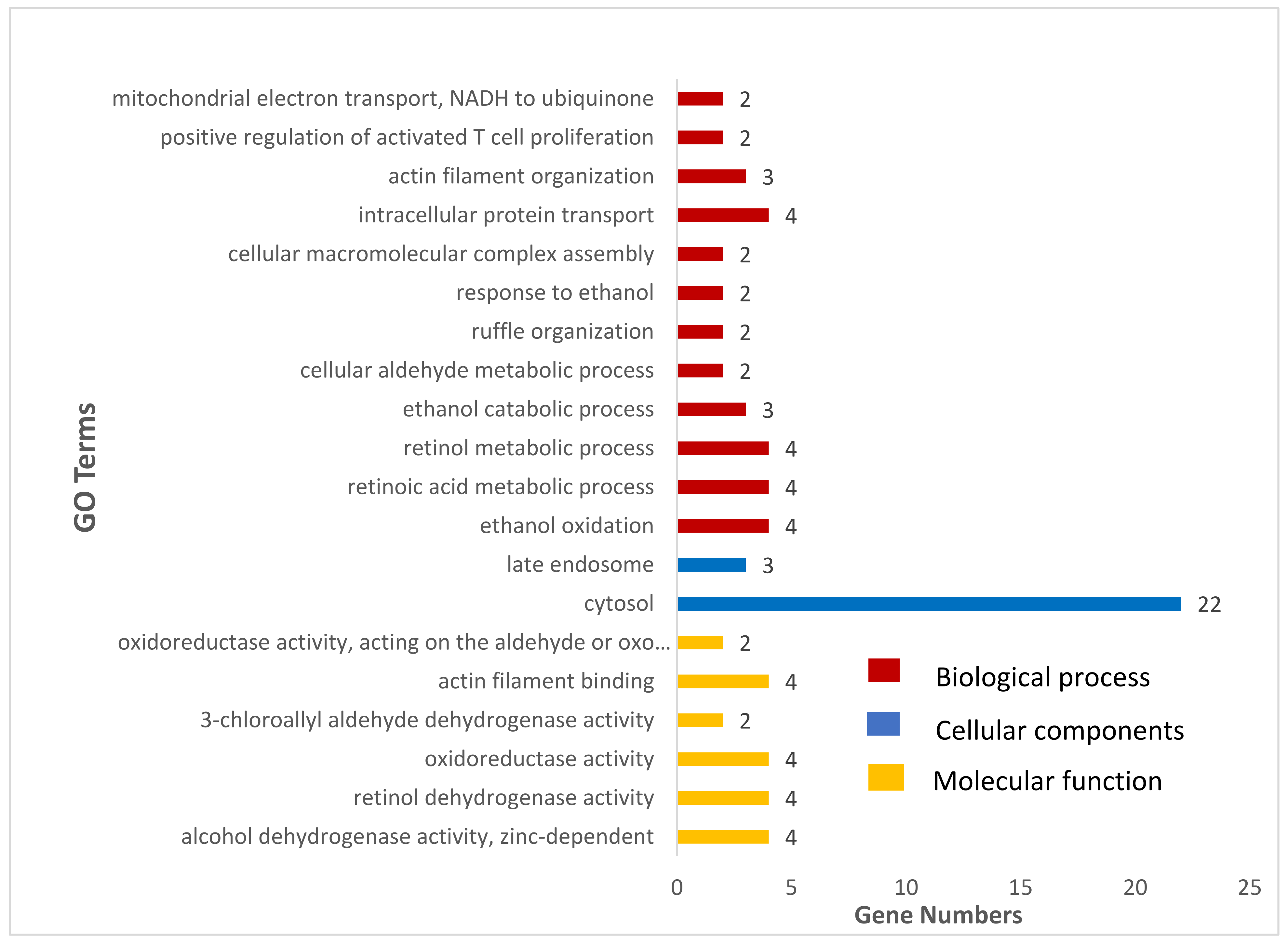
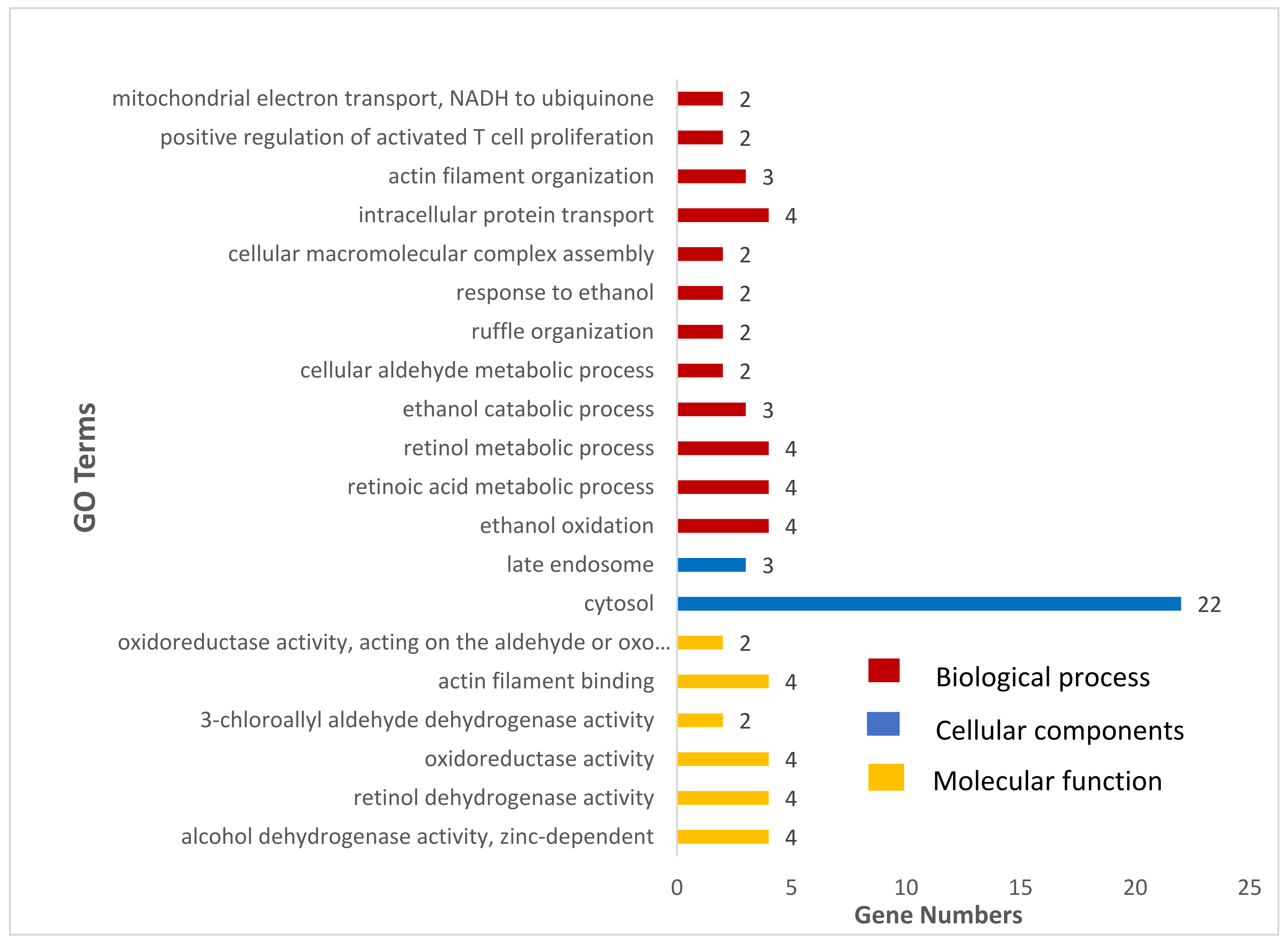
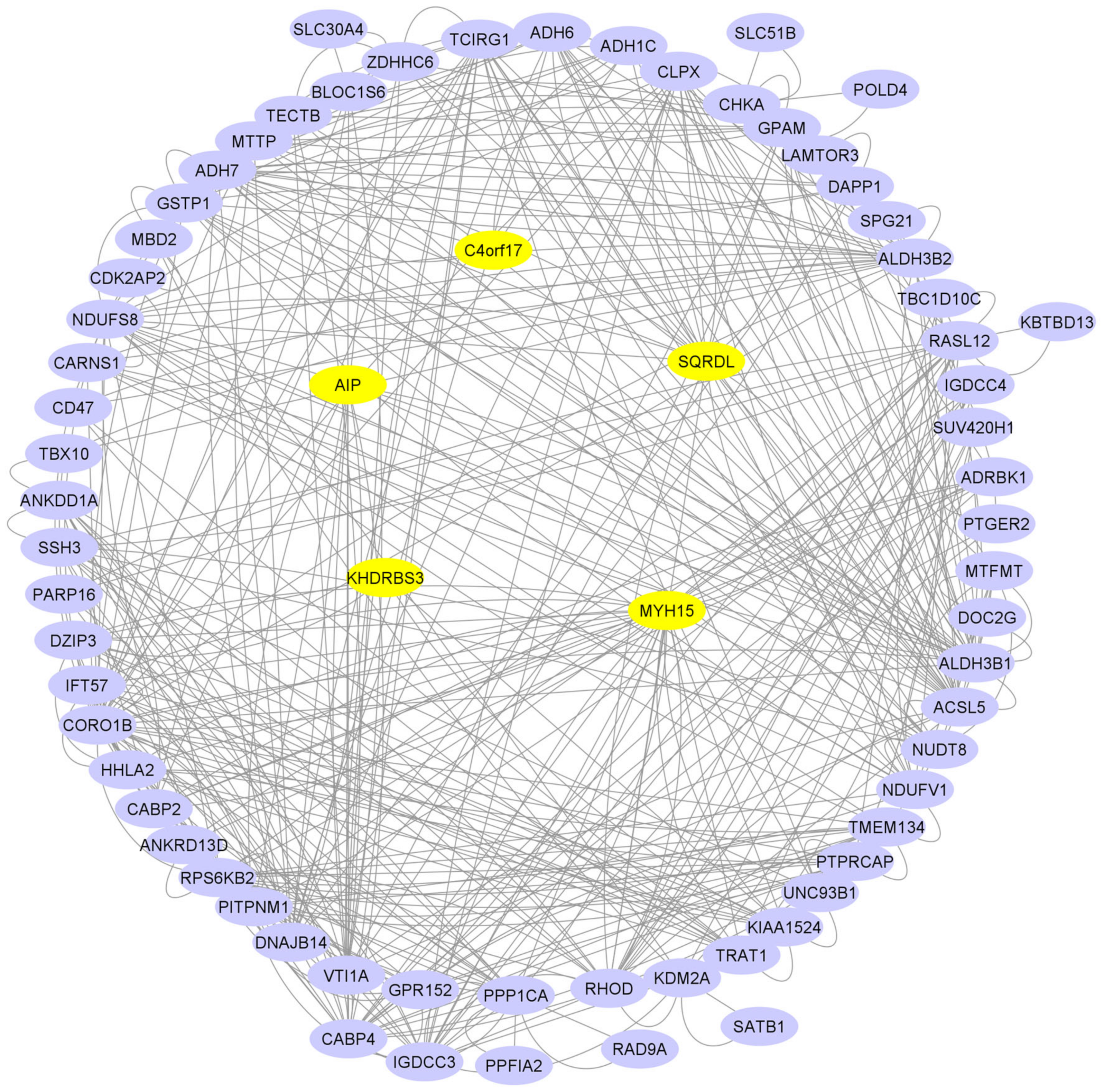
3.3. Gene-Set Enrichment and Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Styrkarsdottir, U.; Stefansson, O.A.; Gunnarsdottir, K.; Thorleifsson, G.; Lund, S.H.; Stefansdottir, L.; Juliusson, K.; Agustsdottir, A.B.; Zink, F.; Halldorsson, G.H.; et al. GWAS of bone size yields twelve loci that also affect height, BMD, osteoarthritis or fractures. Nat. Commun. 2019, 10, 2054. [Google Scholar] [CrossRef] [Green Version]
- Bouwman, A.C.; Daetwyler, H.D.; Chamberlain, A.J.; Ponce, C.H.; Sargolzaei, M.; Schenkel, F.S.; Sahana, G.; Govignon-Gion, A.; Boitard, S.; Dolezal, M.; et al. Meta-analysis of genome-wide association studies for cattle stature identifies common genes that regulate body size in mammals. Nat. Genet. 2018, 50, 362–367. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Cao, Y.; Shan, H.; Wu, J.; Song, X.; Jiang, Y. The GWAS Analysis of Body Size and Population Verification of Related SNPs in Hu Sheep. Front. Genet. 2021, 12, 642552. [Google Scholar] [CrossRef]
- Posbergh, C.J.; Huson, H.J. All sheeps and sizes: A genetic investigation of mature body size across sheep breeds reveals a polygenic nature. Anim. Genet. 2021, 52, 99–107. [Google Scholar] [CrossRef]
- Zhang, X.; Chu, Q.; Guo, G.; Dong, G.; Li, X.; Zhang, Q.; Zhang, S.; Zhang, Z.; Wang, Y. Genome-wide association studies identified multiple genetic loci for body size at four growth stages in Chinese Holstein cattle. PLoS ONE 2017, 12, e0175971. [Google Scholar] [CrossRef] [Green Version]
- An, B.; Xu, L.; Xia, J.; Wang, X.; Miao, J.; Chang, T.; Song, M.; Ni, J.; Xu, L.; Zhang, L.; et al. Multiple association analysis of loci and candidate genes that regulate body size at three growth stages in Simmental beef cattle. BMC Genet. 2020, 21, 32. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Huang, B.; Zhan, J.; Wang, J.; Qu, K.; Zhang, F.; Shen, J.; Jia, P.; Ning, Q.; Zhang, J.; et al. Whole-genome analyses identify loci and selective signals associated with body size in cattle. J. Anim. Sci. 2020, 98, skaa068. [Google Scholar] [CrossRef]
- Gruber, L.; Ledinek, M.; Steininger, F.; Fuerst-Waltl, B.; Zottl, K.; Royer, M.; Krimberger, K.; Mayerhofer, M.; Egger-Danner, C. Body weight prediction using body size measurements in Fleckvieh, Holstein, and Brown Swiss dairy cows in lactation and dry periods. Arch. Anim. Breed. 2018, 61, 413–424. [Google Scholar] [CrossRef]
- Tebug, S.F.; Missohou, A.; Sabi, S.S.; Juga, J.; Poole, E.J.; Tapio, M.; Marshall, K. Using body measurements to estimate live weight of dairy cattle in low-input systems in Senegal. J. Appl. Anim. Res. 2018, 46, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Yin, T.; König, S. Genetic parameters for body weight from birth to calving and associations between weights with test-day, health, and female fertility traits. J. Dairy Sci. 2018, 101, 2158–2170. [Google Scholar] [CrossRef] [Green Version]
- Johanson, J.M.; Berger, P.J.; Tsuruta, S.; Misztal, I. A Bayesian threshold-linear model evaluation of perinatal mortality, dystocia, birth weight, and gestation length in a Holstein herd. J. Dairy Sci. 2011, 94, 450–460. [Google Scholar] [CrossRef] [Green Version]
- Berry, D.P.; Buckley, F.; Dillon, P.; Evans, R.D.; Rath, M.; Veerkamp, R.F. Genetic relationships among body condition score, body weight, milk yield, and fertility in dairy cows. J. Dairy Sci. 2003, 86, 2193–2204. [Google Scholar] [CrossRef] [Green Version]
- Pozveh, S.T.; Shadparvar, A.A.; Shahrbabak, M.M.; Taromsari, M.D. Genetic analysis of reproduction traits and their relationship with conformation traits in Holstein cows. Livest. Sci. 2009, 125, 84–87. [Google Scholar] [CrossRef]
- Plassais, J.; Kim, J.; Davis, B.W.; Karyadi, D.M.; Hogan, A.N.; Harris, A.C.; Decker, B.; Parker, H.G.; Ostrander, E.A. Whole genome sequencing of canids reveals genomic regions under selection and variants influencing morphology. Nat. Commun. 2019, 10, 1489. [Google Scholar] [CrossRef] [Green Version]
- Dechow, C.D.; Rogers, G.W.; Klei, L.; Lawlor, T.J. Heritabilities and correlations among body condition score, dairy form and selected linear type traits. J. Dairy Sci. 2003, 86, 2236–2242. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.B.; Wiggans, G.R.; Ma, L.; Sonstegard, T.S.; Lawlor, T.J.; Crooker, B.A.; Van Tassell, C.P.; Yang, J.; Wang, S.; Matukumalli, L.K.; et al. Genome-wide association analysis of thirty one production, health, reproduction and body conformation traits in contemporary U.S. Holstein cows. BMC Genom. 2011, 12, 408. [Google Scholar] [CrossRef] [Green Version]
- Yin, T.; König, S. Genome-wide associations and detection of potential candidate genes for direct genetic and maternal genetic effects influencing dairy cattle body weight at different ages. Genet. Sel. Evol. 2019, 51, 4. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Fang, M.; Liu, L.; Wang, S.; Liu, J.; Ding, X.; Zhang, S.; Zhang, Q.; Zhang, Y.; Qiao, L.; et al. Genome wide association studies for body conformation traits in the Chinese Holstein cattle population. BMC Genom. 2013, 14, 897. [Google Scholar] [CrossRef] [Green Version]
- Abo-Ismail, M.K.; Brito, L.F.; Miller, S.P.; Sargolzaei, M.; Grossi, D.A.; Moore, S.S.; Plastow, G.; Stothard, P.; Nayeri, S.; Schenkel, F.S. Genome-wide association studies and genomic prediction of breeding values for calving performance and body conformation traits in Holstein cattle. Genet. Sel. Evol. 2017, 49, 82. [Google Scholar] [CrossRef] [Green Version]
- Madsen, P.; Jensen, J.A. A User’S Guide to DMU. A Package for Analysing Multivariate Mixed Models. Available online: https://dmu.ghpc.au.dk/dmu/index.html (accessed on 25 June 2022).
- Nazar, M.; Abdalla, I.M.; Chen, Z.; Ullah, N.; Liang, Y.; Chu, S.; Xu, T.; Mao, Y.; Yang, Z.; Lu, X. Genome-Wide Association Study for Udder Conformation Traits in Chinese Holstein Cattle. Animals 2022, 12, 2542. [Google Scholar] [CrossRef]
- Abdalla, I.M.; Lu, X.; Nazar, M.; Adam, A.; Arbab, I.; Xu, T.; Yousif, M.H.; Mao, Y.; Yang, Z. Genome-Wide Association Study Identifies Candidate Genes Associated with Feet and Leg Conformation Traits in Chinese Holstein Cattle. 2021, 11, 2259. Animals 2021, 11, 2259. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative Usage of Fixed and Random Effect Models for Powerful and Efficient Genome-Wide Association Studies. PLoS Genet. 2016, 12, e1005767. [Google Scholar] [CrossRef]
- Haldar, T.; Ghosh, S. Effect of Population Stratification on False Positive Rates of Population-Based Association Analyses of Quantitative Traits. Ann. Hum. Genet. 2012, 76, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Marees, A.T.; de Kluiver, H.; Stringer, S.; Vorspan, F.; Curis, E.; Marie-Claire, C.; Derks, E.M. A tutorial on conducting genome-wide association studies: Quality control and statistical analysis. Int. J. Methods Psychiatr. Res. 2018, 27, e1608. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Tian, F.; Pan, Y.; Buckler, E.S.; Zhang, Z. A SUPER powerful method for genome wide association study. PLoS ONE 2014, 9, e107684. [Google Scholar] [CrossRef]
- Armstrong, R.A. When to use the Bonferroni correction. Ophthalmic Physiol. Opt. 2014, 34, 502–508. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Tan, Q.; Collins, J.R.; Alvord, W.G.; Roayaei, J.; Stephens, R.; Baseler, M.W.; Lane, H.C.; Lempicki, R.A. The DAVID Gene Functional Classification Tool: A novel biological module-centric algorithm to functionally analyze large gene lists. Genome Biol. 2007, 8, R183. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Weedon, M.N.; Frayling, T.M. Reaching new heights: Insights into the genetics of human stature. Trends Genet. 2008, 24, 595–603. [Google Scholar] [CrossRef]
- Doran, A.G.; Berry, D.P.; Creevey, C.J. Whole genome association study identifies regions of the bovine genome and biological pathways involved in carcass trait performance in Holstein-Friesian cattle. BMC Genom. 2014, 15, 837. [Google Scholar] [CrossRef] [Green Version]
- Jin, C.; Wang, M.; Song, W.; Kong, X.; Zhang, F.; Zhang, Q.; He, Y. Genome-wide identification, characterization and expression profiling of myosin family genes in sebastes schlegelii. Genes 2021, 12, 808. [Google Scholar] [CrossRef]
- Bolormaa, S.; Pryce, J.E.; Reverter, A.; Zhang, Y.; Barendse, W.; Kemper, K.; Tier, B.; Savin, K.; Hayes, B.J.; Goddard, M.E. A Multi-Trait, Meta-analysis for Detecting Pleiotropic Polymorphisms for Stature, Fatness and Reproduction in Beef Cattle. PLoS Genet. 2014, 10, e1004198. [Google Scholar] [CrossRef] [Green Version]
- Bolormaa, S.; Hayes, B.J.; van der Werf, J.H.J.; Pethick, D.; Goddard, M.E.; Daetwyler, H.D. Detailed phenotyping identifies genes with pleiotropic effects on body composition. BMC Genom. 2016, 17, 224. [Google Scholar] [CrossRef] [Green Version]
- Kong, B.W.; Hudson, N.; Seo, D.; Lee, S.; Khatri, B.; Lassiter, K.; Cook, D.; Piekarski, A.; Dridi, S.; Anthony, N.; et al. RNA sequencing for global gene expression associated with muscle growth in a single male modern broiler line compared to a foundational Barred Plymouth Rock chicken line. BMC Genom. 2017, 18, 82. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Du, H.; Yang, C.; Li, Q.; Qiu, M.; Song, X.; Yu, C.; Jiang, X.; Liu, L.; Hu, C.; et al. Comparative transcriptome analysis reveals regulators mediating breast muscle growth and development in three chicken breeds. Anim. Biotechnol. 2019, 30, 233–241. [Google Scholar] [CrossRef]
- Nihashi, Y.; Umezawa, K.; Shinji, S.; Hamaguchi, Y.; Kobayashi, H.; Kono, T.; Ono, T.; Kagami, H.; Takaya, T. Distinct cell proliferation, myogenic differentiation, and gene expression in skeletal muscle myoblasts of layer and broiler chickens. Sci. Rep. 2019, 9, 16527. [Google Scholar] [CrossRef] [Green Version]
- Buaban, S.; Lengnudum, K.; Boonkum, W.; Phakdeedindan, P. Genome-wide association study on milk production and somatic cell score for Thai dairy cattle using weighted single-step approach with random regression test-day model. J. Dairy Sci. 2022, 105, 468–494. [Google Scholar] [CrossRef] [PubMed]
- da Cruz, A.S.; Silva, D.C.; Minasi, L.B.; de Farias Teixeira, L.K.; Rodrigues, F.M.; da Silva, C.C.; do Carmo, A.S.; da Silva, M.V.G.B.; Utsunomiya, Y.T.; Garcia, J.F.; et al. Single-Nucleotide Polymorphism Variations Associated With Specific Genes Putatively Identified Enhanced Genetic Predisposition for 305-Day Milk Yield in the Girolando Crossbreed. Front. Genet. 2021, 11, 1720. [Google Scholar] [CrossRef]
- Clancey, E.; Kiser, J.N.; Moraes, J.G.N.; Dalton, J.C.; Spencer, T.E.; Neibergs, H.L. Genome-wide association analysis and gene set enrichment analysis with SNP data identify genes associated with 305-day milk yield in Holstein dairy cows. Anim. Genet. 2019, 50, 254–258. [Google Scholar] [CrossRef]
- Mudadu, M.A.; Porto-Neto, L.R.; Mokry, F.B.; Tizioto, P.C.; Oliveira, P.S.N.; Tullio, R.R.; Nassu, R.T.; Niciura, S.C.M.; Tholon, P.; Alencar, M.M.; et al. Genomic structure and marker-derived gene networks for growth and meat quality traits of Brazilian Nelore beef cattle. BMC Genom. 2016, 17, 235. [Google Scholar] [CrossRef] [Green Version]
- Srikanth, K.; Lee, S.H.; Chung, K.Y.; Park, J.E.; Jang, G.W.; Park, M.R.; Kim, N.Y.; Kim, T.H.; Chai, H.H.; Park, W.C.; et al. A gene-set enrichment and protein–protein interaction network-based gwas with regulatory snps identifies candidate genes and pathways associated with carcass traits in hanwoo cattle. Genes 2020, 11, 316. [Google Scholar] [CrossRef] [Green Version]
- Deng, T.X.; Ma, X.Y.; Lu, X.R.; Duan, A.Q.; Shokrollahi, B.; Shang, J.H. Signatures of selection reveal candidate genes involved in production traits in Chinese crossbred buffaloes. J. Dairy Sci. 2022, 105, 1327–1337. [Google Scholar] [CrossRef] [PubMed]
- Trivellin, G.; Korbonits, M. REVIEW AIP and its interacting partners. J. Endocrinol. 2011, 210, 137–155. [Google Scholar] [CrossRef] [Green Version]
- Stojanovic, M.; Wu, Z.; Stiles, C.E.; Miljic, D.; Soldatovic, I.; Pekic, S.; Doknic, M.; Petakov, M.; Popovic, V.; Strasburger, C.; et al. Circulating aryl hydrocarbon receptor-interacting protein (Aip) is independent of gh secretion. Endocr. Connect. 2019, 8, 326–337. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Liu, Y.; Sun, X.; Lin, Y.; Yin, D.; Xu, S.; Yang, G. Insights into body size variation in cetaceans from the evolution of body-size-related genes. BMC Evol. Biol. 2019, 19, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osterberg, J.S.; Cammen, K.M.; Schultz, T.F.; Clark, B.W.; Di Giulio, R.T. Genome-wide scan reveals signatures of selection related to pollution adaptation in non-model estuarine Atlantic killifish (Fundulus heteroclitus). Aquat. Toxicol. 2018, 200, 73–82. [Google Scholar] [CrossRef]
- Yin, T.; Jaeger, M.; Scheper, C.; Grodkowski, G.; Sakowski, T.; Klopcic, M.; Bapst, B.; König, S. Multi-breed genome-wide association studies across countries for electronically recorded behavior traits in local dual-purpose cows. PLoS ONE 2019, 14, e0221973. [Google Scholar] [CrossRef]
- Sheet, S.; Krishnamoorthy, S.; Cha, J.; Choi, S.; Choi, B.-W. Identification of Candidate Genes and Pathways Associated with Obesity-Related Traits in Canines via Gene-Set Enrichment and Pathway-Based GWAS Analysis. Animals 2020, 10, 2071. [Google Scholar] [CrossRef]
- Strillacci, M.G.; Moradi-Shahrbabak, H.; Davoudi, P.; Ghoreishifar, S.M.; Mokhber, M.; Masroure, A.J.; Bagnato, A. A genome-wide scan of copy number variants in three Iranian indigenous river buffaloes. BMC Genom. 2021, 22, 305. [Google Scholar] [CrossRef]
- Jackson, M.R.; Melideo, S.L.; Jorns, M.S. Role of Human Sulfide: Quinone Oxidoreductase in H2S Metabolism, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2015; Volume 554. [Google Scholar]
- Zhuang, Z.; Xu, L.; Yang, J.; Gao, H.; Zhang, L.; Gao, X.; Li, J.; Zhu, B. Weighted single-step genome-wide association study for growth traits in chinese simmental beef cattle. Genes 2020, 11, 189. [Google Scholar] [CrossRef] [Green Version]
- De Las Heras-Saldana, S.; Clark, S.A.; Duijvesteijn, N.; Gondro, C.; Van Der Werf, J.H.J.; Chen, Y. Combining information from genome-wide association and multi-tissue gene expression studies to elucidate factors underlying genetic variation for residual feed intake in Australian Angus cattle. BMC Genom. 2019, 20, 939. [Google Scholar] [CrossRef] [Green Version]
- Nazar, M.; Lu, X.; Abdalla, I.M.; Ullah, N.; Fan, Y.; Chen, Z.; Arbab, A.A.I.; Mao, Y.; Yang, Z. Genome-Wide Association Study Candidate Genes on Mammary System-Related Teat-Shape Conformation Traits in Chinese Holstein Cattle. Genes 2021, 12, 2020. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, H.; Yao, C.; Pang, D.; Lu, C.; Xu, A.; Huang, P.; Yu, H. Data mining and validation of ampk pathway as a novel candidate role affecting intramuscular fat content in pigs. Animals 2019, 9, 137. [Google Scholar] [CrossRef] [Green Version]
- Zhao, K.; Chen, Y.H.; Penner, G.B.; Oba, M.; Guan, L.L. Transcriptome analysis of ruminal epithelia revealed potential regulatory mechanisms involved in host adaptation to gradual high fermentable dietary transition in beef cattle. BMC Genom. 2017, 18, 976. [Google Scholar] [CrossRef] [Green Version]
- Krone, K.G.; Ward, A.K.; Madder, K.M.; Hendrick, S.; Mckinnon, J.J.; Buchanan, F.C. Interaction of Vitamin A supplementation level with ADH1C genotype on intramuscular fat in beef steers. Animal 2015, 10, 403–409. [Google Scholar] [CrossRef] [Green Version]
- Ghafouri, F.; Bahrami, A.; Sadeghi, M.; Miraei-Ashtiani, S.R.; Bakherad, M.; Barkema, H.W.; Larose, S. Omics Multi-Layers Networks Provide Novel Mechanistic and Functional Insights Into Fat Storage and Lipid Metabolism in Poultry. Front. Genet. 2021, 12, 646297. [Google Scholar] [CrossRef]
- Lee, H.J.; Chung, Y.J.; Jang, S.; Seo, D.W.; Lee, H.K.; Yoon, D.; Lim, D.; Lee, S.H. Genome-wide identification of major genes and genomic prediction using high-density and text-mined gene-based SNP panels in Hanwoo (Korean cattle). PLoS ONE 2020, 15, e0241848. [Google Scholar] [CrossRef]
- Li, J.; Wang, Y.; Mukiibi, R.; Karisa, B.; Plastow, G.S.; Li, C. Integrative analyses of genomic and metabolomic data reveal genetic mechanisms associated with carcass merit traits in beef cattle. Sci. Rep. 2022, 12, 3389. [Google Scholar] [CrossRef]
- Li, J.; Mukiibi, R.; Wang, Y.; Plastow, G.S.; Li, C. Identification of candidate genes and enriched biological functions for feed efficiency traits by integrating plasma metabolites and imputed whole genome sequence variants in beef cattle. BMC Genom. 2021, 22, 823. [Google Scholar] [CrossRef]
- Mukiibi, R.; Vinsky, M.; Keogh, K.; Fitzsimmons, C.; Stothard, P.; Waters, S.M.; Li, C. Liver transcriptome profiling of beef steers with divergent growth rate, feed intake, or metabolic body weight phenotypes. J. Anim. Sci. 2019, 97, 4386–4404. [Google Scholar] [CrossRef] [Green Version]
- Medeiros De Oliveira Silva, R.; Stafuzza, N.B.; Fragomeni, B.D.O.; Ferreira De Camargo, G.M.; Ceacero, T.M.; Cyrillo, J.N.D.S.G.; Baldi, F.; Boligon, A.A.; Mercadante, M.E.Z.; Lourenco, D.L.; et al. Genome-wide association study for carcass traits in an experimental nelore cattle population. PLoS ONE 2017, 12, e0169860. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Mean | Std. Error | Min. | Max. | Std. Deviation |
|---|---|---|---|---|---|
| BH | 7.8608 | 0.04402 | 1.00 | 9.00 | 1.3807 |
| BD | 7.1189 | 0.05837 | 3.00 | 9.00 | 1.8308 |
| CW | 8.2053 | 0.03573 | 4.00 | 9.00 | 1.1206 |
| ANG | 6.1753 | 0.03895 | 2.00 | 9.00 | 1.2218 |
| Trait | BH | BD | CW | ANG |
|---|---|---|---|---|
| BH | 0.48 (0.03) | 0.939 | 0.703 | 0.517 |
| BD | 0.096 | 0.10 (0.01) | 0.8578 | 0.052 |
| CW | 0.164 | 0.784 | 0.17 (0.01) | 0.173 |
| ANG | 0.140 | −0.334 | −0.277 | 0.19 (0.01) |
| Trait | SNP | rs. SNP | Chr. | Position (bp) | MAF | p-Value | Nearest Gene | Distance (kb) |
|---|---|---|---|---|---|---|---|---|
| BH | BovineHD1100016691 | rs134484400 | 11 | 57,877,493 | 0.3748 | 1.36 × 10−07 | - | - |
| ARS-BFGL-NGS-18743 | rs110462304 | 1 | 53,214,714 | 0.3391 | 1.86 × 10−07 | MYH15 | Within | |
| Hapmap28262-BTA-143868 | rs109930583 | 6 | 25,113,885 | 0.0643 | 2.06 × 10−07 | LOC112447047 C6H4orf17 | Within | |
| Hapmap23799-BTC-047701 | rs109824125 | 14 | 6,680,908 | 0.3411 | 4.98 × 10−07 | KHDRBS3 | 100 kb | |
| ARS-BFGL-NGS-24800 | rs42188649 | 29 | 45,369,368 | 0.1940 | 5.80 × 10−07 | AIP | Within | |
| BD | BovineHD2400015228 | rs133735152 | 24 | 53,151,923 | 0.4279 | 2.33 × 10−08 | DCC | 100 kb |
| BTB-00074122 | rs43286429 | 1 | 155,166,581 | 0.2502 | 4.71 × 10−07 | LOC112447004 | 100 kb | |
| CW | BovineHD1000018705 | rs110355602 | 10 | 64,626,909 | 0.3784 | 9.45 × 10−11 | SQOR | 200 kb |
| BovineHD1000004064 | rs43615333 | 10 | 12,056,720 | 0.4366 | 1.17 × 10−07 | UBAP1L | Within | |
| BTB-00939179 | rs42095998 | 26 | 33,060,404 | 0.2293 | 8.22 × 10−07 | VTI1A | Within | |
| ANG | BovineHD0500003481 | rs135918869 | 5 | 11,715,037 | 0.1649 | 1.32 × 10−07 | CCDC59 | 100 kb |
| Terms | Description | % | p-Value | Gene Name |
|---|---|---|---|---|
| bta00982 | Drug metabolism-cytochrome P450 | 0.063892 | 1.49 × 10−07 | LOC508879, ADH1C, GSTP1, ALDH3B1, ADH7, ADH6 |
| bta00980 | Metabolism of xenobiotics by cytochrome P450 | 0.063892 | 1.65 × 10−07 | LOC508879, ADH1C, GSTP1, ALDH3B1, ADH7, ADH6 |
| bta05204 | Chemical carcinogenesis | 0.063892 | 4.65 × 10−07 | LOC508879, ADH1C, GSTP1, ALDH3B1, ADH7, ADH6 |
| bta00350 | Tyrosine metabolism | 0.045637 | 2.47 × 10−05 | LOC508879, ADH1C, ALDH3B1, ADH7, ADH6 |
| bta00010 | Glycolysis/Gluconeogenesis | 0.045637 | 1.50 × 10−04 | LOC508879, ADH1C, ALDH3B1, ADH7, ADH6 |
| bta00071 | Fatty acid degradation | 0.03651 | 7.24 × 10−04 | ADH1C, ACSL5, ADH7, ADH6 |
| bta00340 | Histidine metabolism | 0.027382 | 0.004052 | LOC508879, ALDH3B1, CARNS1 |
| bta00410 | beta-Alanine metabolism | 0.027382 | 0.009524 | LOC508879, ALDH3B1, CARNS1 |
| bta01100 | Metabolic pathways | 0.109529 | 0.012732 | POLD4, NDUFS8, LOC508879, GPAM, CHKA, ADH1C, ALDH3B1, ACSL5, TCIRG1, ADH7, NDUFV1, ADH6 |
| bta00830 | Retinol metabolism | 0.027382 | 0.024631 | ADH1C, ADH7, ADH6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdalla, I.M.; Hui, J.; Nazar, M.; Arbab, A.A.I.; Xu, T.; Abdu, S.M.N.; Mao, Y.; Yang, Z.; Lu, X. Identification of Candidate Genes and Functional Pathways Associated with Body Size Traits in Chinese Holstein Cattle Based on GWAS Analysis. Animals 2023, 13, 992. https://doi.org/10.3390/ani13060992
Abdalla IM, Hui J, Nazar M, Arbab AAI, Xu T, Abdu SMN, Mao Y, Yang Z, Lu X. Identification of Candidate Genes and Functional Pathways Associated with Body Size Traits in Chinese Holstein Cattle Based on GWAS Analysis. Animals. 2023; 13(6):992. https://doi.org/10.3390/ani13060992
Chicago/Turabian StyleAbdalla, Ismail Mohamed, Jiang Hui, Mudasir Nazar, Abdelaziz Adam Idriss Arbab, Tianle Xu, Shaima Mohamed Nasr Abdu, Yongjiang Mao, Zhangping Yang, and Xubin Lu. 2023. "Identification of Candidate Genes and Functional Pathways Associated with Body Size Traits in Chinese Holstein Cattle Based on GWAS Analysis" Animals 13, no. 6: 992. https://doi.org/10.3390/ani13060992
APA StyleAbdalla, I. M., Hui, J., Nazar, M., Arbab, A. A. I., Xu, T., Abdu, S. M. N., Mao, Y., Yang, Z., & Lu, X. (2023). Identification of Candidate Genes and Functional Pathways Associated with Body Size Traits in Chinese Holstein Cattle Based on GWAS Analysis. Animals, 13(6), 992. https://doi.org/10.3390/ani13060992