
Mortality Causes in Captive Cantabrian capercaillie (Tetrao urogallus cantabricus) in Spain

 , , , , ,
, , , , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction

2. Materials and Methods
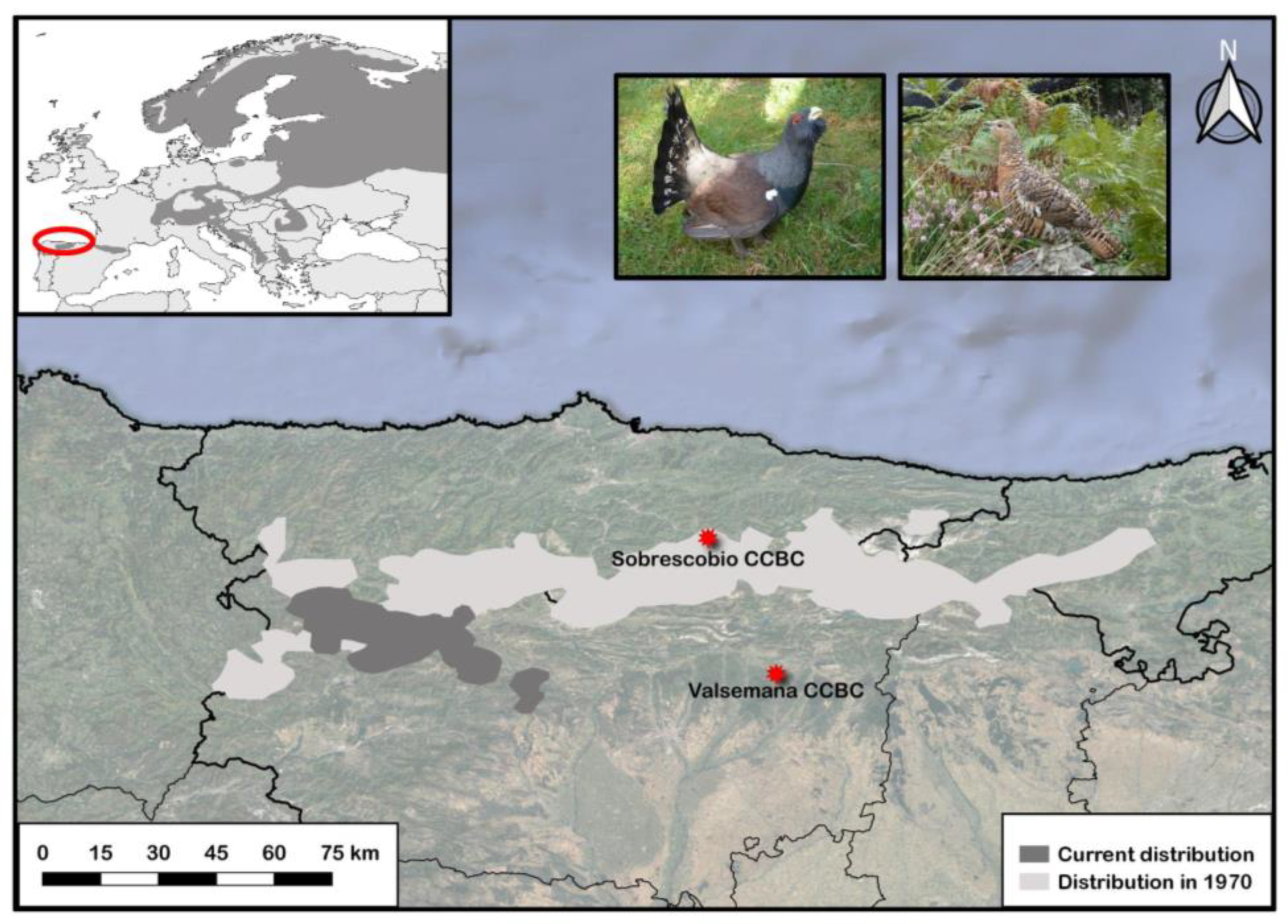
2.1. Sobrescobio CCBC: Facilities and Management of Captive Capercaillies
2.2. Data Collection
2.3. Diagnostic Procedures
3. Results
3.1. Classification of Causes of Death
3.2. Description of Causes of Death
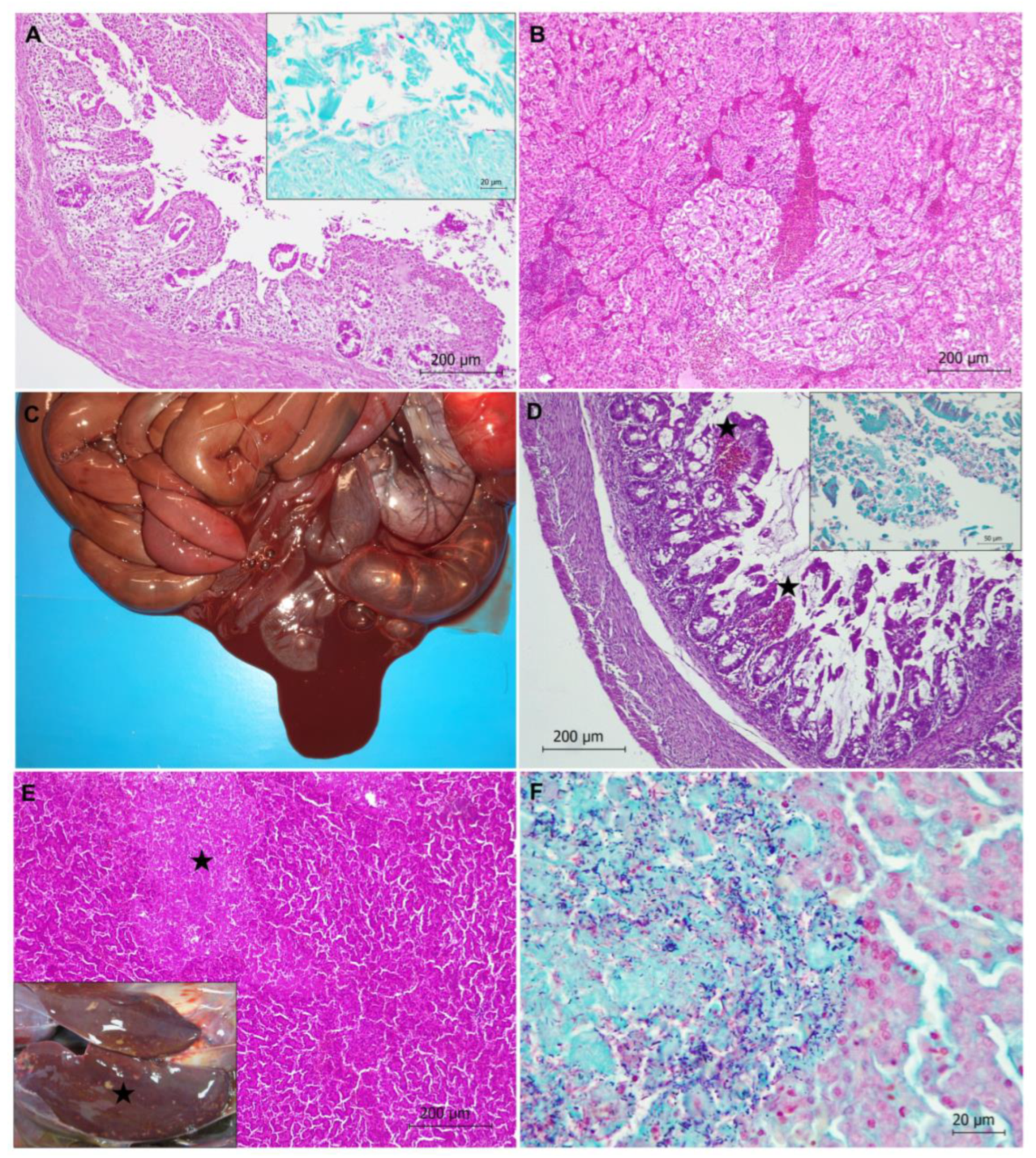
3.2.1. Infectious Diseases
Bacterial Diseases
Generalized Aspergillosis
3.2.2. Non-Infectious Diseases
Neurogenic Shock
Rupture of the Heart Apex and Cardiomyopathy
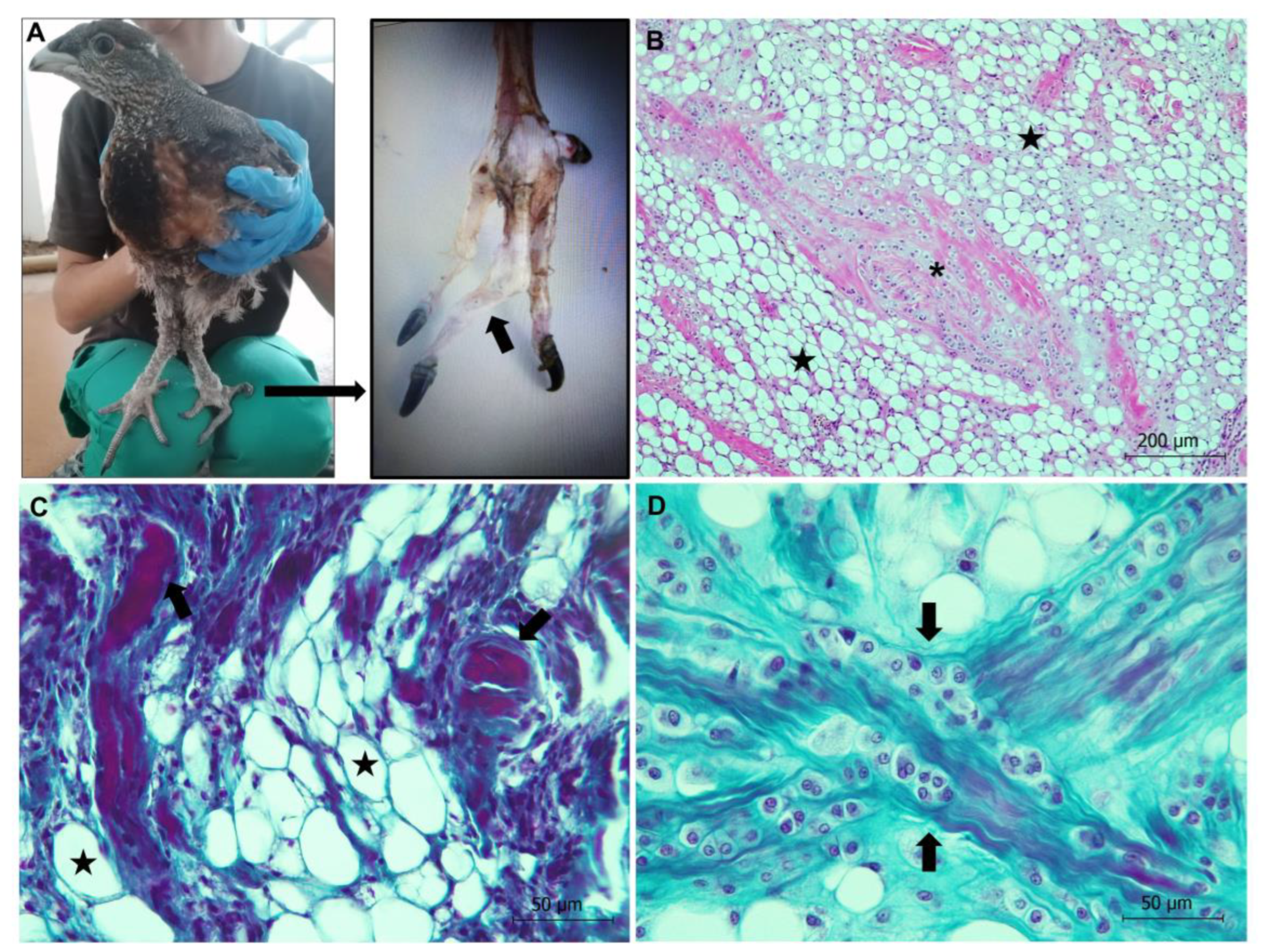
Valgus Leg Deformity
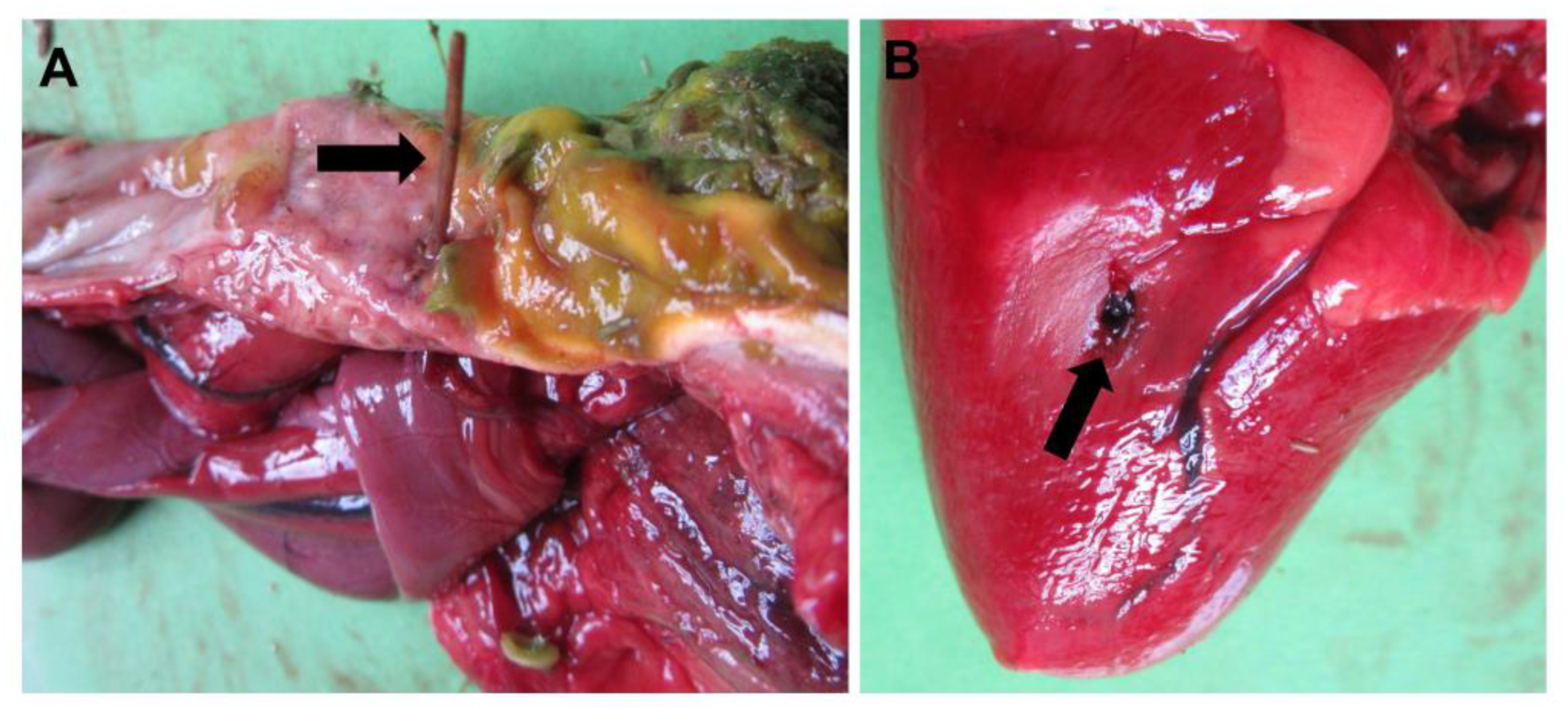
Duodenal Obstruction and Proventriculus Perforation
Exertional (Degenerative) Myopathy in Two Free-Ranging Capercaillies
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Convention on Biological Diversity. Convention on Biological Diversity: Text and Annexes; Secretariat of the Convention on Biological Diversity: Montreal, QC, Canada, 1992; pp. 1–34. [Google Scholar]
- Maxted, N. (Ed.) In situ, ex situ conservation. In Encyclopedia of Biodiversity, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 313–323. [Google Scholar]
- Wakchaure, R.; Ganguly, S. Captive breeding in endangered wildlife: A review. J. Biol. Sci. Opi. 2016, 4, 186. [Google Scholar] [CrossRef]
- Snyder, N.F.; Derrickson, S.R.; Beissinger, S.R.; Wiley, J.W.; Smith, T.B.; Toone, W.D.; Miller, B. Limitations of captive breeding in endangered species recovery. Conserv. Biol. 1996, 10, 338–348. [Google Scholar] [CrossRef] [Green Version]
- Vargas, A.; Anderson, S.H. Growth and physical development of captive-raised black-footed ferrets (Mustela nigripes). Am. Midl. Nat. 1996, 135, 43–52. [Google Scholar] [CrossRef]
- Kreger, M.D.; Hatfield, J.S.; Estevez, I.; Gee, G.F.; Clugston, D.A. The effects of captive rearing on the behavior of newly-released whooping cranes (Grus americana). Appl. Anim. Behav. 2005, 93, 165–178. [Google Scholar] [CrossRef]
- Kraaijeveld-Smit, F.J.; Griffiths, R.A.; Moore, R.D.; Beebee, T.J. Captive breeding and the fitness of reintroduced species: A test of the responses to predators in a threatened amphibian. J. Appl. Ecol. 2006, 43, 360–365. [Google Scholar] [CrossRef]
- Van Heezik, Y.; Maloney, R.F.; Seddon, P.J. Movements of translocated captive-bred and released critically endangered kaki (black stilts) Himantopus novaezelandiae and the value of long-term post-release monitoring. Oryx 2009, 43, 639–647. [Google Scholar] [CrossRef] [Green Version]
- Yerga, J.; Calzada, J.; Manteca, X.; Vargas, A.; Rivas, A. Early development and growth in captive-born Iberian lynx (Lynx pardinus). Zoo Biol. 2014, 33, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Padilla, J.; Estrada Acedo, A. Estrés fisiológico en urogallos cantábricos (Tetrao urogallus cantabricus) silvestres y en cautividad: Variaciones temporales e implicaciones metodológicas. Ecosistemas 2021, 30, 2161. [Google Scholar] [CrossRef]
- de Juana, E.; Kirwan, G.M. Western capercaillie (Tetrao urogallus). In Birds of the World; del Hoyo, J., Elliott, A., Sargatal, J., Christie, D., de Juana, E., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar] [CrossRef]
- Quevedo, M.; Bañuelos, M.J.; Sáez, O.; Obeso, J.R. Habitat selection by Cantabrian capercaillie Tetrao urogallus cantabricus at the edge of the species’ distribution. Wildlife Biol. 2006, 12, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Venturas, M.; García Álvarez, S.; Fajardo Alcántara, M.; Collada, C.; Gil, L. Species selection for reforestations: What happens with historical extinctions and habitat protection zones? A case study in the Cantabrian Range. Eur. J. Forest Res. 2013, 132, 107–120. [Google Scholar] [CrossRef]
- Blanco-Fontao, B.; Fernández-Gil, A.; Obeso, J.R.; Quevedo, M. Diet and habitat selection in Cantabrian Capercaillie (Tetrao urogallus cantabricus): Ecological differentiation of a rear-edge population. J. Ornithol. 2010, 151, 269–277. [Google Scholar] [CrossRef]
- Ministerio para la Transición Ecológica. Orden TEC/1078/2018, de 28 de Septiembre, por la que se Declara la Situación Crítica de Cistus heterophyllus carthaginensis, Lanius minor, Margaritifera auricularia, Marmaronetta angustirostris, Mustela lutreola, Pinna nobilis y Tetrao urogallus. Available online: https://www.boe.es/eli/es/o/2018/09/28/tec1078/dof/spa/pdf (accessed on 28 February 2023).
- Castroviejo, J.; Delibes, M.; García-Dory, M.Á.; Junco, E. Censo de urogallos cantábricos (Tetrao urogallus cantabricus). Asturnatura 1974, 2, 53–74. [Google Scholar]
- Jiménez, J.; Godinho, R.; Pinto, D.; Lopes, S.; Castro, D.; Cubero, D.; Osorio, M.A.; Piqué, J.; Moreno-Opo, R.; Quirós, P.; et al. The Cantabrian capercaillie: A population on the edge. Sci. Total Environ. 2022, 821, 153523. [Google Scholar] [CrossRef]
- Storch, I. Conservation status and threats to grouse worldwide: An overview. Wildlife Biol. 2000, 6, 195–204. [Google Scholar] [CrossRef]
- Moss, R.; Oswald, J.; Baines, D. Climate change and breeding success: Decline of the capercaillie in Scotland. J. Anim. Ecol. 2001, 70, 47–61. [Google Scholar] [CrossRef]
- Penteriani, V.; Zarzo-Arias, A.; Novo-Fernández, A.; Bombieri, G.; López-Sánchez, C.A. Responses of an endangered brown bear population to climate change based on predictable food resource and shelter alterations. Glob. Change Biol. 2019, 25, 1133–1151. [Google Scholar] [CrossRef]
- Ministerio de Medio Ambiente. Estrategia para la Conservación del Urogallo Cantábrico (Tetrao urogallus cantabricus) en España. Comisión Nacional de Protección de la Naturaleza, Grupo de Trabajo del Urogallo. Madrid: Ministerio de Medio Ambiente. Available online: https://www.miteco.gob.es/es/biodiversidad/publicaciones/pbl_estrategia_urogallo_cantabrico_tcm30-197267.pdf (accessed on 28 February 2023).
- Lifeurogallo. Available online: http://lifeurogallo.es/sites/default/files/resumen_ejecutivo_web.pdf (accessed on 28 February 2023).
- Royal Decree 53/2013. Available online: https://www.boe.es/eli/es/rd/2013/02/01/53 (accessed on 28 February 2023).
- Walsh, T.J.; Wissel, M.C.; Grantham, K.J.; Petraitiene, R.; Petraitis, V.; Kasal, M.; Francesconi, A.; Cotton, M.P.; Hughes, J.E.; Greene, L.; et al. Molecular detection and species-specific identification of medically important Aspergillus species by real-time PCR in experimental invasive pulmonary aspergillosis. J. Clin. Microbiol. 2011, 49, 4150–4157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, G.D.; Garner, M.M.; Raymond, J.T. Causes of mortality in captive Attwater´s prairie chickens (Tympanuchus cupido attwateri) at the San Antonio Zoo, 1997–2000. J. Zoo Wildl. Med. 2002, 33, 236–241. [Google Scholar] [CrossRef]
- Bailey, T.A.; Nicholls, P.K.; Samour, J.H.; Naldo, J.; Wernery, U.; Howlett, J.C. Postmortem findings in bustards in the United Arab Emirates. Avian Dis. 1996, 40, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Hanselmann, R.; Hallager, S.; Murray, S.; Mazet, J. Causes of morbidity and mortality in captive kori bustards (Ardeotis kori) in the United States. J. Zoo Wildl. Med. 2013, 44, 348–363. [Google Scholar] [CrossRef] [PubMed]
- Gibson, D.J.; Nemeth, N.M.; Beaufrère, H.; Varga, C.; Eagalle, T.; Susta, L. Captive Psittacine birds in Ontario, Canada: A 19-year retrospective study of the causes of morbidity and mortality. J. Comp. Pathol. 2019, 171, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, J.; Goryo, M.; Makara, M.; Nakamura, K.; Okada, K. Necrotic hepatitis due to Clostridium perfringens infection in newly hatched broiler chicks. J. Vet. Med. 2003, 65, 1249–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcano, V.; Gamble, T.; Maschek, K.; Stabler, L.; Fletcher, O.; Davis, J.; Troan, B.V.; Villegas, A.M.; Tsai, Y.Y.; Barbieri, N.L.; et al. Necrotizing hepatitis associated with Clostridium perfringens in broiler chicks. Avian Dis. 2002, 66, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Pavlickova, S.; Dolezalova, M.; Holko, I. Resistance and virulence factors of Escherichia coli isolated from chicken. J. Environ. Sci. Health B 2015, 50, 417–421. [Google Scholar] [CrossRef]
- Stuve, G.; Hofshagen, M.; Holt, G. Necrotizing lesions in the intestine, gizzard, and liver in captive capercaillies (Tetrao urogallus) associated with Clostridium perfringens. J. Wildl. Dis. 1992, 28, 598–602. [Google Scholar] [CrossRef] [Green Version]
- Prescott, J.F.; Parreira, V.R.; Mehdizadeh Gohari, I.; Lepp, D.; Gong, J. The pathogenesis of necrotic enteritis in chickens: What we know and what we need to know: A review. Avian Pathol. 2016, 45, 288–294. [Google Scholar] [CrossRef] [Green Version]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.M.; Finlay, B.B. Gut microbiota in health and disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef] [Green Version]
- Moore, R.J. Necrotic enteritis predisposing factors in broiler chickens. Avian Pathol. 2016, 45, 275–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.; Morgan, N.K.; Moss, A.F.; Li, L.; Ader, P.; Choct, M. The flow of non-starch polysaccharides along the gastrointestinal tract of broiler chickens fed either a wheat- or maize-based diet. Anim. Nutr. 2022, 9, 138–142. [Google Scholar] [CrossRef]
- Daneshmand, A.; Kermanshahi, H.; Mohammed, J.; Sekhavati, M.H.; Javadmanesh, A.; Ahmadian, M.; Alizadeh, M.; Razmyar, J.; Kulkarni, R.R. Intestinal changes and immune responses during Clostridium perfringens-induced necrotic enteritis in broiler chickens. Poult. Sci. J. 2022, 101, 1–9. [Google Scholar] [CrossRef]
- Hofshagen, M.; Stenwig, H. Toxin production by Clostridium perfringens isolated from broiler chickens and capercaillies (Tetrao urogallus) with and without necrotizing enteritis. Avian Dis. 1992, 36, 837–843. [Google Scholar] [CrossRef]
- Schales, K.; Gerlach, H.; Kösters, J. Investigations on the aerobic flora and Clostridium perfringens in fecal specimens from free-living and captive capercaillies (Tetrao urogallus L., 1758). J. Vet. Med. 1993, 40, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Wieneman, T.; Schmitt-Wagner, D.; Meuser, K.; Segelbacher, G.; Schink, B.; Brune, A.; Berthold, P. The bacterial microbiota in the ceca of Capercaillie (Tetrao urogallus) differs between wild and captive birds. Syst. Appl. Microbiol. 2011, 34, 542–551. [Google Scholar] [CrossRef]
- Wang, W.; Li, Z.; Han, Q.; Guo, Y.; Zhang, B.; D’inca, R. Dietary live yeast and mannan-oligosaccharide supplementation attenuate intestinal inflammation and barrier dysfunction induced by Escherichia coli in broilers. Br. J. Nutr. 2016, 116, 1878–1888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, N.; Smyth, J.A. Oral vaccination of broiler chickens against necrotic enteritis using a non-virulent NetB positive strain of Clostridium perfringens type A. Vaccine 2017, 35, 6858–6865. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.T.; Zeng, X.F.; Chen, A.G.; Zhou, L.; Zhang, L.; Xiao, Y.P.; Yang, C.M. Effects of a probiotic, Enterococcus faecium, on growth performance, intestinal morphology, immune response, and cecal microflora in broiler chickens challenged with Escherichia coli K88. Poult. Sci. J. 2013, 92, 2949–2955. [Google Scholar] [CrossRef] [PubMed]
- Schales, C.; Gerlach, H.; Kösters, J. Investigations on the antibacterial effect of conifer needle oils on bacteria isolated from the feces of captive Capercaillies (Tetrao urogallus L., 1758). Zentralbl. Veterinarmed. B 1993, 40, 381–390. [Google Scholar] [CrossRef]
- Wegge, P.; Kastdalen, L. Habitat and diet of young grouse broods: Resource partitioning between Capercaillie (Tetrao urogallus) and Black Grouse (Tetrao tetrix) in boreal forests. J. Ornithol. 2008, 149, 237–244. [Google Scholar] [CrossRef]
- Schmidt, A. Georg Fresenius und die spezies Aspergillus fumigatus: Georg Fresenius and the species Aspergillus fumigatus. Mycoses 1998, 41, 89–91. [Google Scholar] [CrossRef]
- Beernaert, L.A.; Pasmans, F.; Van Waeyenberghe, L.; Haesebrouck, F.; Martel, A. Aspergillus infections in birds: A review. Avian Pathol. 2010, 39, 325–331. [Google Scholar] [CrossRef] [Green Version]
- Arné, P.; Risco-Castillo, V.; Jouvion, G.; Le Barzic, C.; Guillot, J. Aspergillosis in Wild Birds. J. Fungi 2021, 7, 241. [Google Scholar] [CrossRef]
- De Herdt, P. Aspergillose bij papegaaien. Vlaams Diergeneeskd Tijdschr. 1996, 65, 343–344. [Google Scholar]
- Schmidt, A.; Schmidt, D.I.J.B. Georg W. Fresenius and the description of the species Aspergillus fumigatus in 1863. Contrib. Microbiol. 1999, 2, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Domingo, M.; Marco-Sanchez, I.; Marco-Valle, A.J.; Pumarola, M. Heart rupture and haemopericardium in capercaillie (Tetrao urogallus) reared in captivity. Avian Pathol. 1991, 20, 363–366. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.W.; Kidd, M.T., Jr.; Kadhim, H.J.; Shouse, S.; Orlowski, S.K.; Hiltz, J.; Anthony, N.B.; Kuenzel, W.J.; Kong, B.C. Characterization of stress response involved in chicken myopathy. Gen. Comp. Endocrinol. 2020, 295, 113526. [Google Scholar] [CrossRef]
- Hasegawa, Y.; Hosotani, M.; Saito, M.; Nagasawa, T.; Mori, Y.; Kawasaki, T.; Yamada, M.; Maeda, N.; Watanabe, T.; Iwasaki, T. Mitochondrial characteristics of chicken breast muscle affected by wooden breast. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2022, 273, 111296. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.Q.; Ren, F.Z.; Jiang, Y.Y.; Xiao, C.; Lei, X.G. Selenoproteins protect against avian nutritional muscular dystrophy by metabolizing peroxides and regulating redox/apoptotic signaling. Free Radic. Biol. Med. 2015, 83, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.H.; Huang, J.Q.; Deng, J.; Lei, X.G. Avian selenogenome: Response to dietary Se and vitamin E deficiency and supplementation. Poult. Sci. 2019, 98, 4247–4254. [Google Scholar] [CrossRef] [PubMed]
- Maxie, M.G. (Ed.) Cardiovascular system. In Jubb, Kennedy and Palmer’s Pathology of Domestic Animals, 6th ed.; Elsevier: London, UK, 2016; pp. 2–99. [Google Scholar]
- Castroviejo, J. El urogallo "Tetrao urogallus. L." en España; Monografías de la Estación Biológica de Doñana 3; Consejo Superior de Investigaciones Científicas: Madrid, Spain, 1975. [Google Scholar]
- Osbaldiston, G.W.; Wise, D.B. Spondylolisthesis and leg weakness in the chicken—A common etiology. Vet. Rec. 1967, 80, 320–322. [Google Scholar]
- Randall, C.J. Diseases and Disorders of the Domestic Fowl and Turkey, 2nd ed.; Mosby-Wolfe: London, UK, 1997. [Google Scholar]
- Akbas, Y.; Yalcin, S.; Ozkan, S.; Kirkpinar, F.; Takma, C.; Gevrekçi, Y.; Güler, H.C.; Turkmut, L. Heritability estimates of tibial dyschondroplasia, valgus-varus, foot-pad dermatitis and hock burn in broiler. Arch. Geflügelkd. Sonderh. 2009, 73, 1–6. [Google Scholar]
- Hulan, H.W.; De Groote, G.; Fontaine, G.; De Munter, G.; McRae, K.B.; Proudfoot, F.G. The effect of different totals and ratios of dietary calcium and phosphorus on the performance and incidence of leg abnormalities of male and female broiler chickens. Poult. Sci. J. 1985, 64, 1157–1169. [Google Scholar] [CrossRef] [PubMed]
- Haye, U.; Simons, P.C.M. Twisted legs in broilers. Br. Poult. Sci. 1978, 19, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.E.; Sunde, M.L.; Stahl, J.L.; Hanson, L.E. Zinc deficiency in pheasant chicks fed practical diets. Avian Dis. 1984, 28, 1102–1109. [Google Scholar] [CrossRef] [PubMed]
- Poland, G.; Raftery, A. (Eds.) BSAVA Manual of Backyard Poultry Medicine and Surgery, 1st ed.; BSAVA: London, UK, 2019; p. 222. [Google Scholar]
- Maxie, M.G. (Ed.) Muscle and tendon. In Jubb, Kennedy and Palmer´s Pathology of Domestic Animals, 6th ed.; Elsevier: London, UK, 2016; pp. 165–248. [Google Scholar]
- Liukkonen-Anttila, T. Nutritional and Genetic Adaptation of Galliform Birds: Implications for Hand-Rearing and Restocking; Oulu University Press: Oulu, Finland, 2001. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
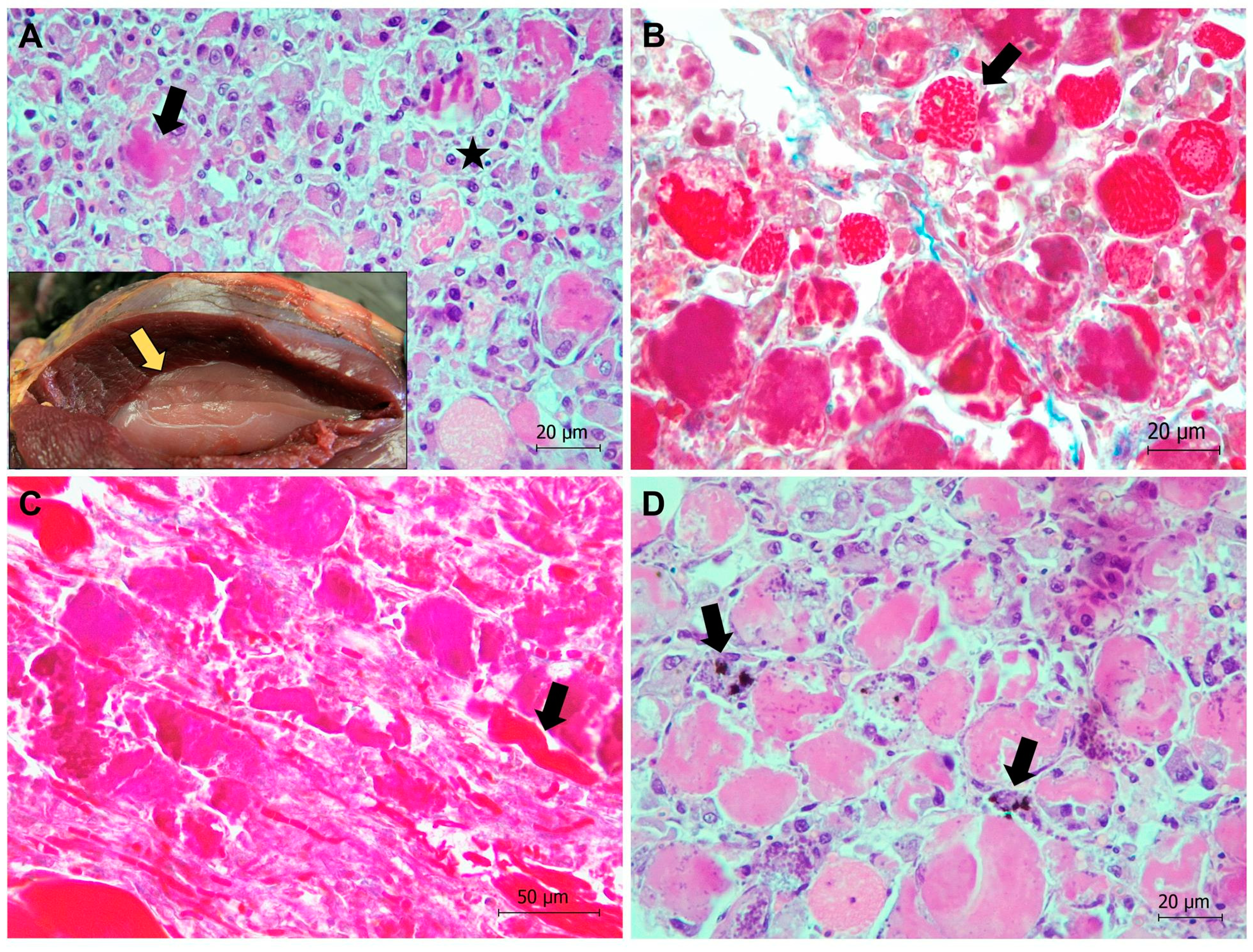{kind=link}
| ID | Date | Age | Sex | Cause of Death | Microbiology | Classification of Death |
|---|---|---|---|---|---|---|
| 1 | 4 July 2015 | 6 days | N.d. | Acute enteritis/septicemia | Culture: Enterococcus spp. (intestine, liver) | I |
| 2 | 29 April 2016 | 5 days | N.d. | Septicemia | Culture: Escherichia coli (intestine, liver) | I |
| 3 | 31 July 2017 | 47 days | Female | Septicemia | Culture: Escherichia coli (intestine, liver, kidney) | I |
| 4 | 25 June 2019 | 4 days | N.d. | Sacculitis | Culture: Escherichia coli (sacculum) | I |
| 5 | 25 June 2019 | 3 days | N.d. | Septicemia | Culture: Escherichia coli (intestine, liver) | I |
| 6 | 25 June 2019 | 3 days | Male | Acute typhlitis | Culture: Escherichia coli (intestine) | I |
| 7 | 12 July 2019 | 8 days | Male | Acute enteritis | Culture: Escherichia coli (intestine) | I |
| 8 | 3 July 2020 | 8 days | Male | Acute enteritis/septicemia | Culture: Escherichia coli (intestine, liver) | I |
| 9 | 3 July 2020 | 8 days | N.d. | Septicemia | Culture: Escherichia coli (fecal content, liver) | I |
| 10 | 16 July 2020 | 15 days | Male | Acute enteritis/septicemia | Culture: Escherichia coli (intestine, liver) | I |
| 11 | 2 July 2021 | 15 days | Male | Acute typhlitis | Culture: Escherichia coli, Enterococcus gallinarum, Clostridium perfringens (intestine) | I |
| 12 | 12 July 2022 | 6 days | Female | Necrotic hemorrhagic enteritis/septicemia | Culture: Clostridium perfringens (intestine, liver) | I |
| 13 | 20 July 2022 | 26 days | Female | Acute enteritis/septicemia | Culture: Escherichia coli (intestine, liver, heart, brain) | I |
| 14 | 21 July 2022 | 6 days | Female | Necrotic hepatitis | - | I |
| 15 | 15 September 2022 | 55 days | Female | Heart apex rupture/cardiomyopathy | - | NI |
| 16 | 10 November 2022 | 2 months | Male | Valgus leg deformity | - | NI-euthanasia |
| ID | Date | Age | Sex | Cause of Death | Microbiology | Classification of Death |
|---|---|---|---|---|---|---|
| 1 | 14 April 2004 | 3 years | Male | Exertional myopathy | - | NI |
| 2 | 12 July 2008 | Adult | Male | Predation-Exertional myopathy | - | NI |
| 3 | 19 June 2012 | 1 year | Male | Aspergillosis | PCR: Aspergillus fumigatus (lungs, air sacs) | I |
| 4 | 12 November 2013 | 2 years | Male | Duodenal obstruction with a hazelnut and coelomitis | - | NI |
| 5 | 26 January 2017 | 1.5 years | Female | Neurogenic shock (stress) | - | NI |
| 6 | 3 February 2017 | 7 months | Female | Cardiomyopathy | - | NI |
| 7 | 3 February 2017 | 7 months | Female | Heart apex rupture/cardiomyopathy | - | NI |
| 8 | 26 January 2018 | 2.5 years | Female | Neurogenic shock (stress) | - | NI |
| 9 | 27 June 2018 | 1 year | Male | Neurogenic shock (stress) | - | NI |
| 10 | 15 March 2019 | 3.5 years | Female | Perforation of the proventriculus and heart with a briar branch | - | NI |
| 11 | 1 March 2020 | 8 years | Female | Heart apex rupture/cardiomyopathy | - | NI |
| 12 | 20 June 2022 | 7 years | Male | Necrotic hepatitis | Culture: Clostridium perfringens (intestine, liver, heart) | I |
| 13 | 6 July 2022 | 10 years | Male | Necrotic hepatitis | - | I |
| 14 | 9 November 2022 | 5 years | Female | Neurogenic shock (stress) | - | NI |
| 15 | 17 November 2022 | 8 years | Female | Purulent myocarditis | - | I |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Rodríguez, A.; Herrero-García, G.; de Garnica García, M.G.; García Esgueva, Á.; Balsera, R.; Oleaga, Á.; Fernández, D.; Amado, J.; Royo, L.J.; García Iglesias, M.J.; et al. Mortality Causes in Captive Cantabrian capercaillie (Tetrao urogallus cantabricus) in Spain. Animals 2023, 13, 1255. https://doi.org/10.3390/ani13071255
García-Rodríguez A, Herrero-García G, de Garnica García MG, García Esgueva Á, Balsera R, Oleaga Á, Fernández D, Amado J, Royo LJ, García Iglesias MJ, et al. Mortality Causes in Captive Cantabrian capercaillie (Tetrao urogallus cantabricus) in Spain. Animals. 2023; 13(7):1255. https://doi.org/10.3390/ani13071255
Chicago/Turabian StyleGarcía-Rodríguez, Alberto, Gloria Herrero-García, María Gracia de Garnica García, Álvaro García Esgueva, Ramón Balsera, Álvaro Oleaga, Daniel Fernández, Javier Amado, Luis José Royo, María José García Iglesias, and et al. 2023. "Mortality Causes in Captive Cantabrian capercaillie (Tetrao urogallus cantabricus) in Spain" Animals 13, no. 7: 1255. https://doi.org/10.3390/ani13071255
APA StyleGarcía-Rodríguez, A., Herrero-García, G., de Garnica García, M. G., García Esgueva, Á., Balsera, R., Oleaga, Á., Fernández, D., Amado, J., Royo, L. J., García Iglesias, M. J., & Balseiro, A. (2023). Mortality Causes in Captive Cantabrian capercaillie (Tetrao urogallus cantabricus) in Spain. Animals, 13(7), 1255. https://doi.org/10.3390/ani13071255