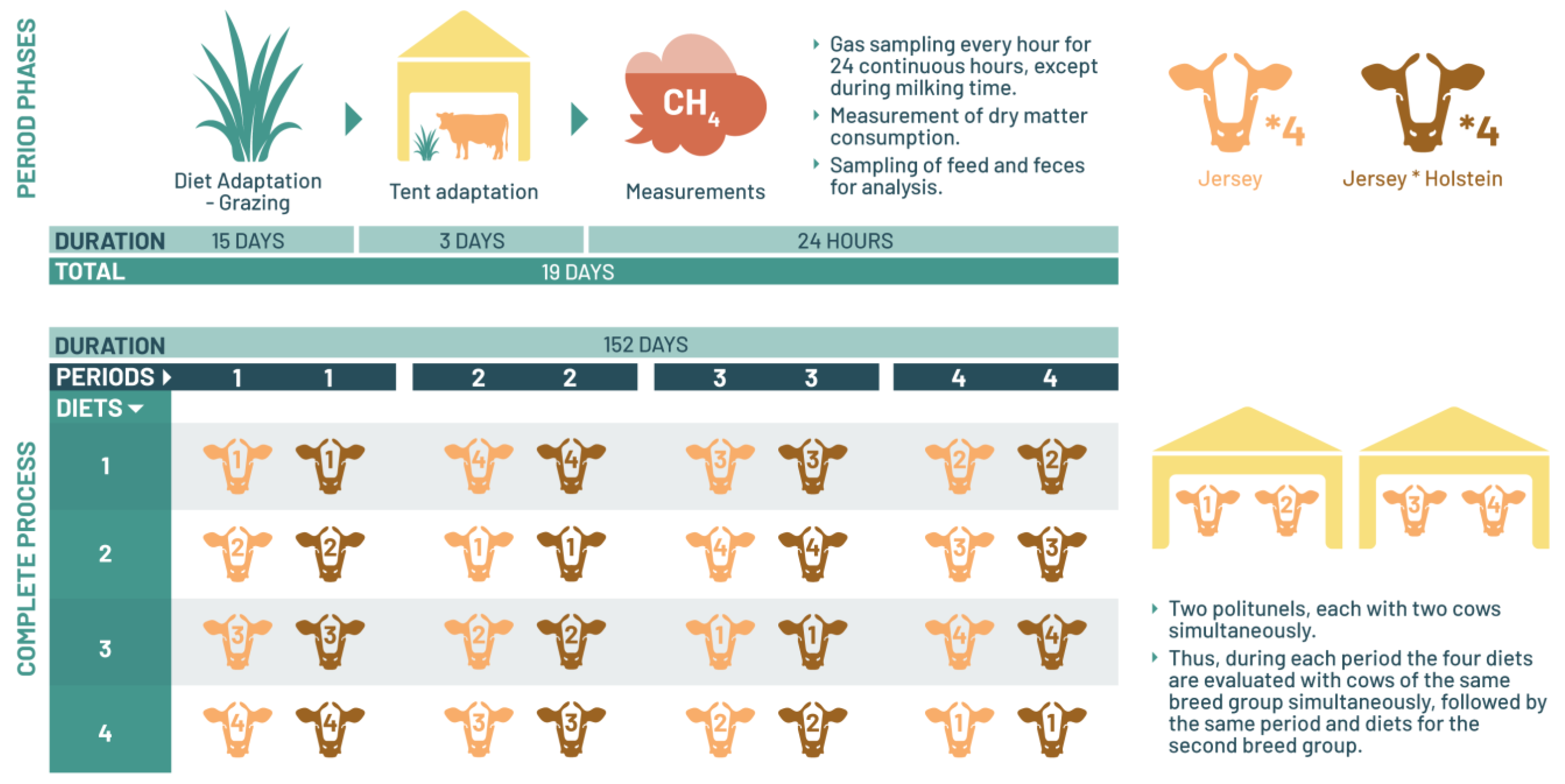
1. Introduction
Globally, methane (CH
4) is the second most important greenhouse gas (GHG) emitted by the agricultural sector, mainly because it has a global warming potential 28 times higher than that of carbon dioxide [
1]. However, its permanence in the atmosphere is lower [
2]. Additionally, developing countries emit 70% of the total global enteric CH
4, of which 25% comes from Latin America and the Caribbean [
3]. In livestock systems, CH
4 is generated as a product of enteric fermentation and is emitted to the environment mainly through belching, which represents a significant energy loss for the animal [
4].
The carbon footprint is the sum of CH
4, nitrous oxide (N
2O), carbon dioxide (CO
2) and other gases emitted directly or indirectly during the process of obtaining a product [
5]. However, it has been stated that the carbon footprint per gram of protein in milk or meat from ruminants is very high compared to other meats, such as pork, poultry, or rabbit, or other animal products such as yogurt or eggs [
6]. For the case of ruminants, the amount of gases in the carbon footprint is directly related to the type of feed consumed, [
7] and indirectly a reduction in the carbon footprint of milk production has been established when animal performance and farm profitability are improved [
8,
9].
In recent years, the livestock sector, as well as other economic sectors, has made great efforts in the search for and implementation of strategies to reduce GHG emissions, particularly of CH
4. The sector aims to achieve sustainable production systems that contribute to the national commitments made before the United National Framework Convention on Climate Change (UNFCCC). Thus, several options have been evaluated to mitigate enteric CH
4 emissions [
10], among which are feeding management, feed composition, forage quality, modifications of the microbial community, chemical manipulation, and animal crossbreeding [
11,
12,
13].
Despite knowledge of these strategies, the Colombian dairy sector resists change due to historically low profit margins. Milk prices hardly compensate for the cost of raw materials such as fertilizers, corn, and soybean used in the production of concentrates [
14]. Adoption of GHG mitigation strategies will be low if they cannot also increase productivity in dairy systems, reduce dependence on external inputs, increase producers’ profits, and make sustainable use of natural resources. In this sense, GHG mitigation actions must allow for the partial or total replacement of grains in concentrate feed formulations with nutrition sources that cost less and are locally available [
15].
In the search for nutritional alternatives, research has been conducted on cassava (
Manihot esculenta Crantz) as a supplement for ruminants. This tropical and subtropical shrubby plant, belonging to the Euphorbiaceae family, is characterized by a high tolerance for poor soils and adverse climatic conditions [
16]. Supplementation with cassava in ruminant feed has been used for its nutritional value [
17,
18]. Among other qualities, roots have a high content of non-structural carbohydrates (75 to 85%) and low levels of crude protein (CP; 2–3% CP), while leaves and green stems contain higher CP values (25%) [
16,
19]. Additionally, multiple authors found antimethanogenic compounds such as tannins and saponins within cassava roots, stems, and leaves [
1,
20,
21].
Despite cassava’s importance—Colombia produces approximately 2.1 million tons per year on 187.2 thousand hectares [
22]—no field-based studies have been carried out on the effect of cassava intake on the combination of productive parameters, carbon footprint, and enteric CH
4 emissions in cattle or the relationship of these parameters with the most common cattle breeds and crosses of specialized dairy cows.
Furthermore, there appears to be a correlation between animal production parameters and genetic characteristics, although the nature of this relationship is complex. Extensive research is essential to comprehensively understand these correlations over time, considering various production systems and environmental conditions [
23]. Notably, crossbreeding has been suggested as a potential avenue for enhancing performance and mitigating CH
4 emissions. Therefore, it is crucial to assess the CH
4 emissions of different breeds and crosses commonly employed in specialized dairy farming to ascertain the genetic connections between methane production and existing selection traits [
24].
Considering the aforementioned factors, this study represents the inaugural investigation conducted under field conditions with specialized dairy cows supplemented with cassava in Colombia. The primary aim was to assess the impact of supplementation with cassava leaves and roots on CH4 emissions, the carbon footprint, and various key productivity parameters.
4. Discussion
Cassava leaves generated the highest protein intake among the treatments consumed by the animals in this study. Cassava foliage has been reported to be an effective source of bypass protein for feedlot steers [
42] and goats [
43]. However, the potential of cassava foliage as a source of protein in ruminant feeds has not been fully exploited, probably due to the risk of toxicity. The contents of the hydrogen cyanide (HCN) precursors linamarin and lotaustralin can range from 364 to 964 g/kg depending on variety, age, and plant part [
19,
44]. Sun drying or ensiling of cassava foliage can reduce HCN toxicity to harmless levels (50 mg/kg) [
45], while also improving palatability and prolonging storage time [
16]. However, Thang et al. [
46] state that high energy intake is needed in cattle diets to counteract the negative effect of HCN. Suharti et al. [
19] recommend supplementing cassava leaves with cyanide-degrading bacteria. These latter two alternatives have shown positive results.
In contrast to cassava leaves, the nutritional content of cassava root is characterized by low amounts of protein (2–3%), macronutrients such as fiber (2%), some vitamins, and minerals. However, it has a high calcium, vitamin C, and starch contents [
47,
48]. The content of this last nutrient in the present investigation was 18% above that reported in the literature (67–81%); this range is a function of the variety of cassava evaluated [
49]. Cassava roots also provide the diet with thiamine, riboflavin, carotenoids, minerals, and nicotinic acid, all essential for proper animal metabolism [
47,
50,
51]. Several reports indicate that cassava supplements can replace corn-, cereal-, or tuber-based supplements, maintaining nutritional intake without negative effects on digestibility [
19,
51,
52]. These reports agree with the results of this study, where no differences were observed in DM or organic matter intake, but differences were observed in CP, NDF, and ADF intake when cows received cassava root in their diet (
Table 3).
Regarding DM digestibility, this study found no differences between treatments, although the structural carbohydrate content was different between treatments (
Table 3). Digestibility may not have been affected because important system components such as protein and energy supplied the requirements for the correct functioning of ruminal microorganisms. Authors such as Thang et al. [
46] and Suharti et al. [
19] concluded that the use of cassava with higher levels of protein and metabolizable energy improved microbial protein synthesis and fiber fermentation in diets of cattle that were fed low-quality pasture. However, the inclusion of 25% cassava leaf meal could reduce nutrient digestibility [
53]. Additionally, Lunsin et al. [
54] reported that supplementation with cassava root hay can improve fiber digestibility, as this type of supplementation significantly improves total bacterial populations of
Ruminococcus flavefaciens and
Fibrobacter succinogens. Similarly, Suharti et al. [
19] found that supplementation with bitter cassava leaves increased the total bacterial population without an effect on the population of ruminal protozoa.
The F1 breed exhibited higher total daily milk production compared to the Jersey breed, reflecting inherent genetic disparities between the two. This observation aligns with findings from a study by Coffey et al. [
55], where a 305-day assessment revealed that Holstein cows outperformed both the Friesian (4591 kg) and Jersey (4230 kg) breeds, recording a superior production of 5217 kg. Interestingly, our study did not reveal a correlation between dry matter intake and milk yield, contrasting with the results of a meta-analysis by Hristov et al. [
56], which showed a moderate linear relationship (R2 = 0.47). This discrepancy might be attributed to the confinement conditions imposed on the animals during our experiment.
The addition of cassava root and leaf did not affect the milk production or milk quality of Jersey and F1 cows (
Table 4), confirming other results reported in the literature [
57]. Although there were differences in the consumption of NDF, a precursor of milk fat, the production of acetic acid in the rumen may not have differed between diets. Pertiwi et al. [
58] concluded that supplementation with 34.5% cassava root husk in Holstein cows positively affected CP and total solids. Their results for fat and milk volume were similar to those of this study. The protein content in milk depends on the energy present in the diet. Therefore, protein contents in milk lower than 3%, values that were not obtained in the present study, are typical of low-energy diets [
59]. Likewise, the MUN values obtained range between 14 and 16 mg/dL, indicating an optimal use of nitrogen [
60].
In the present study, cassava leaf and root treatments did not significantly change CH
4 emissions (
Table 5), despite differences between digestible intakes of ADF and NDF. The NDF is directly and inversely related to dry matter digestibility, CH
4 emissions, intake level, and feeding frequency [
3,
61]. Likewise, the higher contribution of starch present in the root (mainly amylose: 16–18%, and amylopectin: 82–84%) [
62] was expected to have an effect on the reduction of CH
4 emissions [
63]. This effect is expected because the digestion of starch in the rumen is decreased, but the digestion of starch in the small intestine is increased, along with a reduced proportion of acetate and butyrate with respect to propionate production [
64]. A negative effect of the content of secondary compounds on CH
4 emissions would also be expected. Why cassava diets improved some digestibility characteristics but did not change CH
4 emissions remains an open question. Future research could test different cassava sources and higher feed quantities, compared to those used in this study.
In the literature, cassava leaves are reported to contain contents of between 2.06 and 4.36% in tannins as DM in condensed leaves [
20,
21] and between 1.58 and 1.65 mg/100 g of saponins in bitter cassava leaf meal [
65]. These substances lead to decreased CH
4 production and increased efficiency of microbial protein synthesis [
66,
67]. However, this study’s results agree with cattle trials where an increase in the level of starch or consumption of secondary compounds in the diet did not affect CH
4 production per unit DM intake of concentrate or per unit milk produced [
19,
68]. The molecular weight, chemical structure, and/or amount per kg DM of some of these compounds may explain the lack of effects on CH
4 emissions [
69,
70]. According to Binsulong et al. [
52], using cassava instead of rice straw in the ration of Holstein Friesian crossbred bulls decreased net CH
4 emissions due to a reduction in fiber content in the diet.
Net CH
4 emissions were lower in Jersey than in F1 breeds (
Table 5). Authors such as Van Wyngaard et al. [
71] reported values of 21.8 kg CH
4/kg DMI and 6.85% CH
4 conversion rate (Ym) for multiparous Jersey cows that were 100 (±45.8 SD) days postpartum and had a body weight of 408 (±32.5 SD) kg, whose diet was mainly based on kikuyu grass. These values are slightly lower than those for Jersey cows in our study. In other research using the same technique and kikuyu grass as the base diet, Donneys [
72] found that Holstein and Holstein * Simental breed cows of 533 (±81 SD) kg live weight consumed 9.65 kg DM/d and produced 21.65 g CH
4/kg DMI, with a CH
4 conversion rate (Ym) of 6.89%.
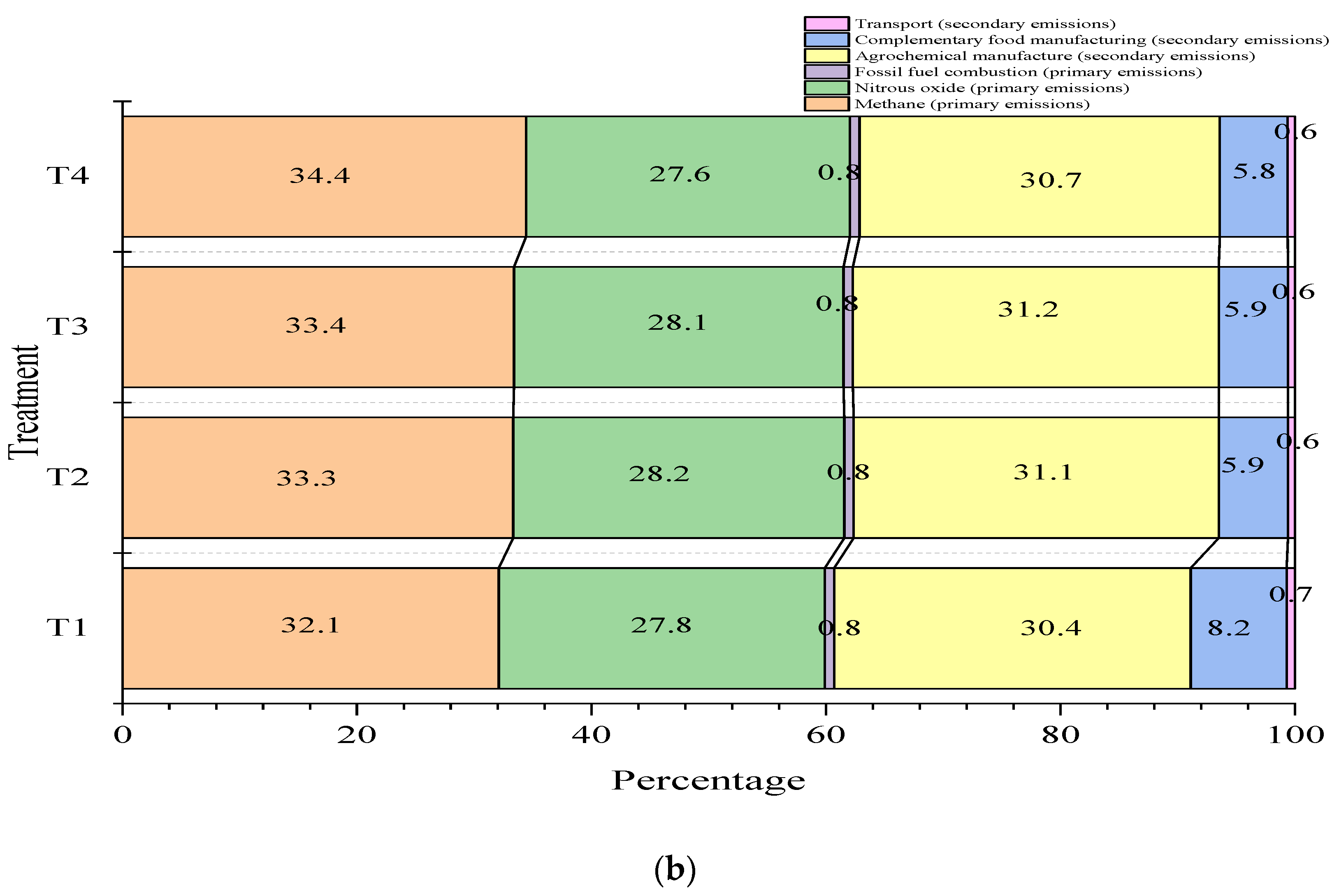
The carbon footprint corrected for fat and protein content in milk was 1.5 kg CO
2eq per kg of FPCM for the four treatments evaluated (
Figure 2 and
Table 6). These values are within the range reported by Mazzetto et al. [
7] for several countries in different continents for non-cassava diets (0.74 and 5.99 kg CO
2eq per kg FPCM). In Colombia, some studies report values for non-cassava diets between 2.1 and 4.2 kg CO
2eq per kg FPCM and between 9.0 and 18.3 CO
2eq per kg of meat for intensive or extensive dual-purpose production systems, respectively [
73]. According to Mazzetto et al. [
7], the distribution of GHGs in the carbon footprint depends on the type of production system. For example, CH
4 emissions predominate in pasture-based livestock farming, while N
2O emissions are highest in intensive or confined livestock farming. In the present study, CH
4 emissions from enteric fermentation are slightly predominant (35.9%), followed by N
2O (27%) from manure management, concentrate production, and fertilizer use.
According to Uddin et al. [
74], cows have a higher carbon footprint when their diets are low in NDF (19% DM) than when the NDF content is high (24% DM; 1.49 vs. 1.35 kg CO
2eq/kg FPCM). However, in the present study the cassava root diet provided 8.5% less NDF (460 g/kg DM) than other supplements evaluated and showed a tendency to have lowest carbon footprint with respect to the rest of the treatments evaluated (<3.8%). This can be explained by the differences between the feed resources used [
74]. Likewise, the literature reports differences between the carbon footprints of milk for different breeds of cows, where Jersey cows produce 4.4% less carbon than Holstein cows (1.41 vs. 1.47 kg CO
2eq/kg FPCM, respectively). Uddin et al. [
74] attribute this difference to the milk produced by Jersey cows, which has higher fat and protein contents than that of Holstein cows. However, in the present study, this difference was not observed. On the contrary, Jersey cows had a higher carbon footprint than Jersey * Holstein crossbred cows.
In this study we were able to use measured emission factors, as opposed to default emission factors, to calculate the carbon footprint. This aspect is important for several reasons including improved accuracy related to the conditions and management practices of the studied system. These measured emission factors considered important factors, such as the local environment, feed sources, and animal breeds, that can significantly affect CH4 emissions. Using measured values leads to more accurate and representative assessments of the carbon footprint.
Default emission factors are often more generalized and may not accurately reflect the specific conditions of this study. The accurate, measured emission factors presented here can be used for developing better accounting (i.e., national GHG inventories), effective policies, and mitigation strategies in Colombia (i.e., nationally determined contributions, NDC). In summary, by using measured emission factors for calculating the carbon footprints of livestock systems in this study, we provide a more accurate, region-specific, and informative basis for understanding and addressing CH4 emissions.


 ,
, 



{kind=link}
{kind=link}
{kind=link}