
Exploring the Microbial Community Structure in the Chicken House Environment by Metagenomic Analysis
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling of PM2.5 and DNA Extraction
2.2. Metagenomic Sequencing
2.3. Taxonomic Assignment and Functional Annotation
2.4. Gene Function Analysis
2.5. Statistical Analysis
3. Results and Discussion
3.1. Sequencing, Assembly, and Microbial Taxonomy
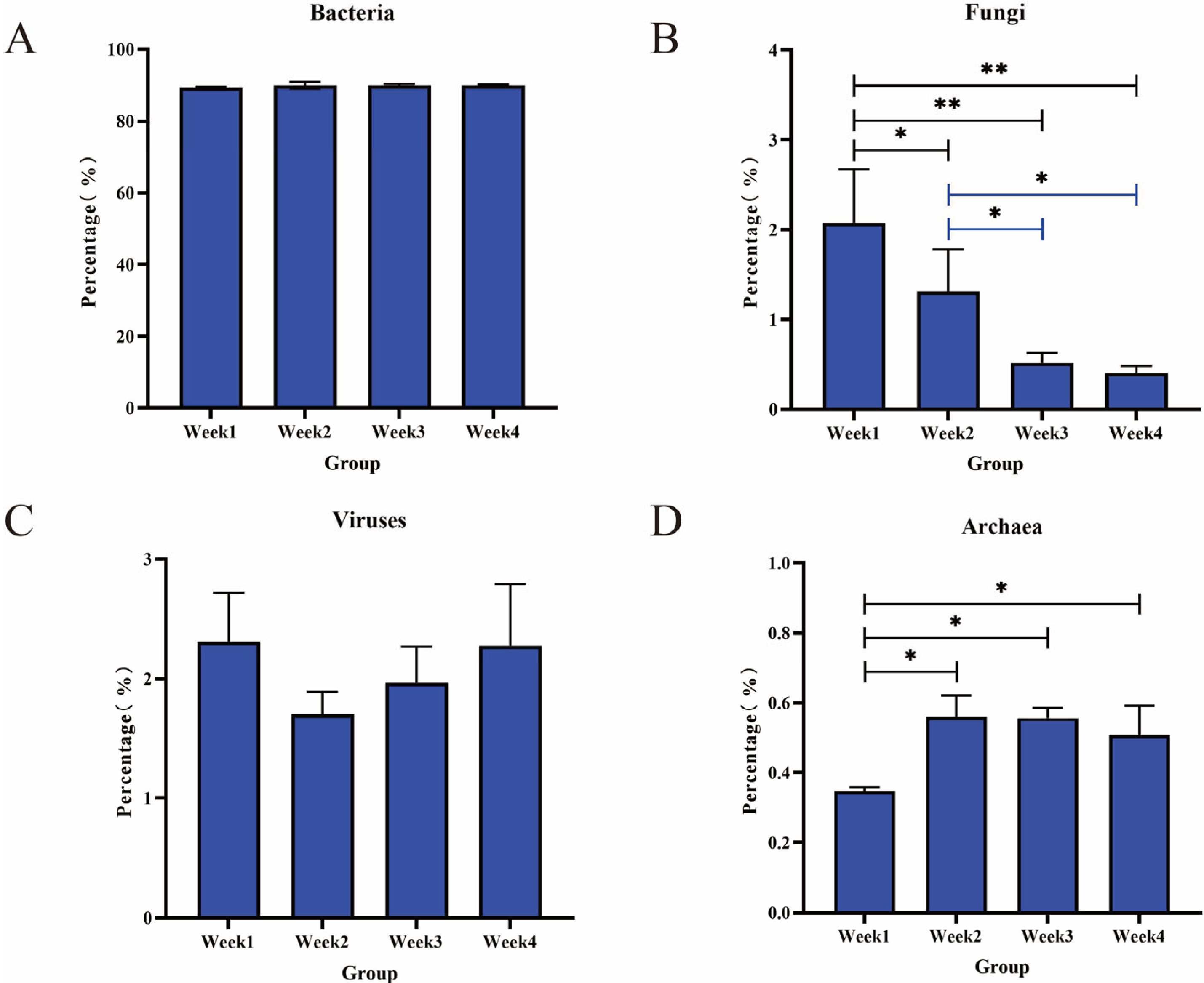
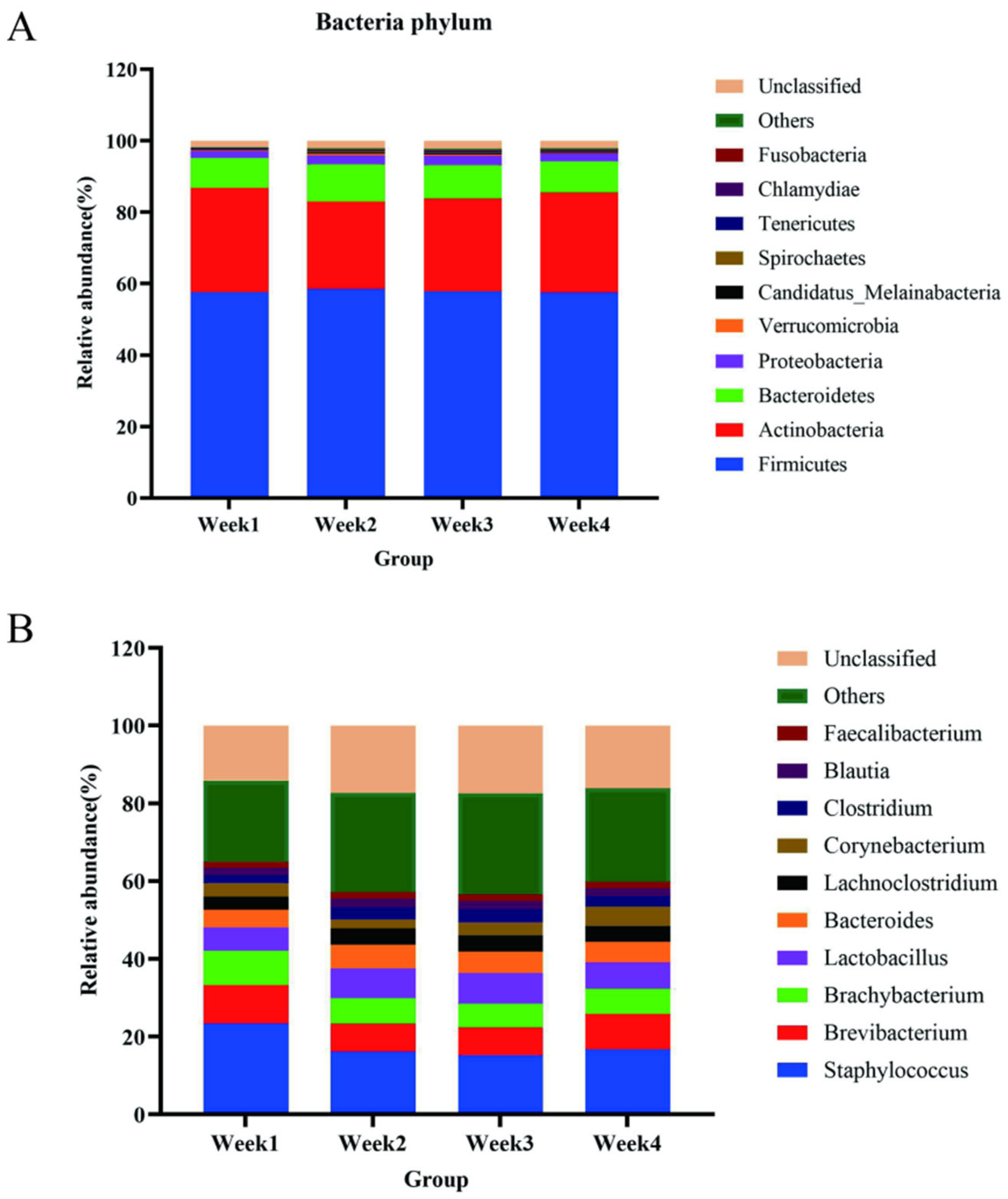
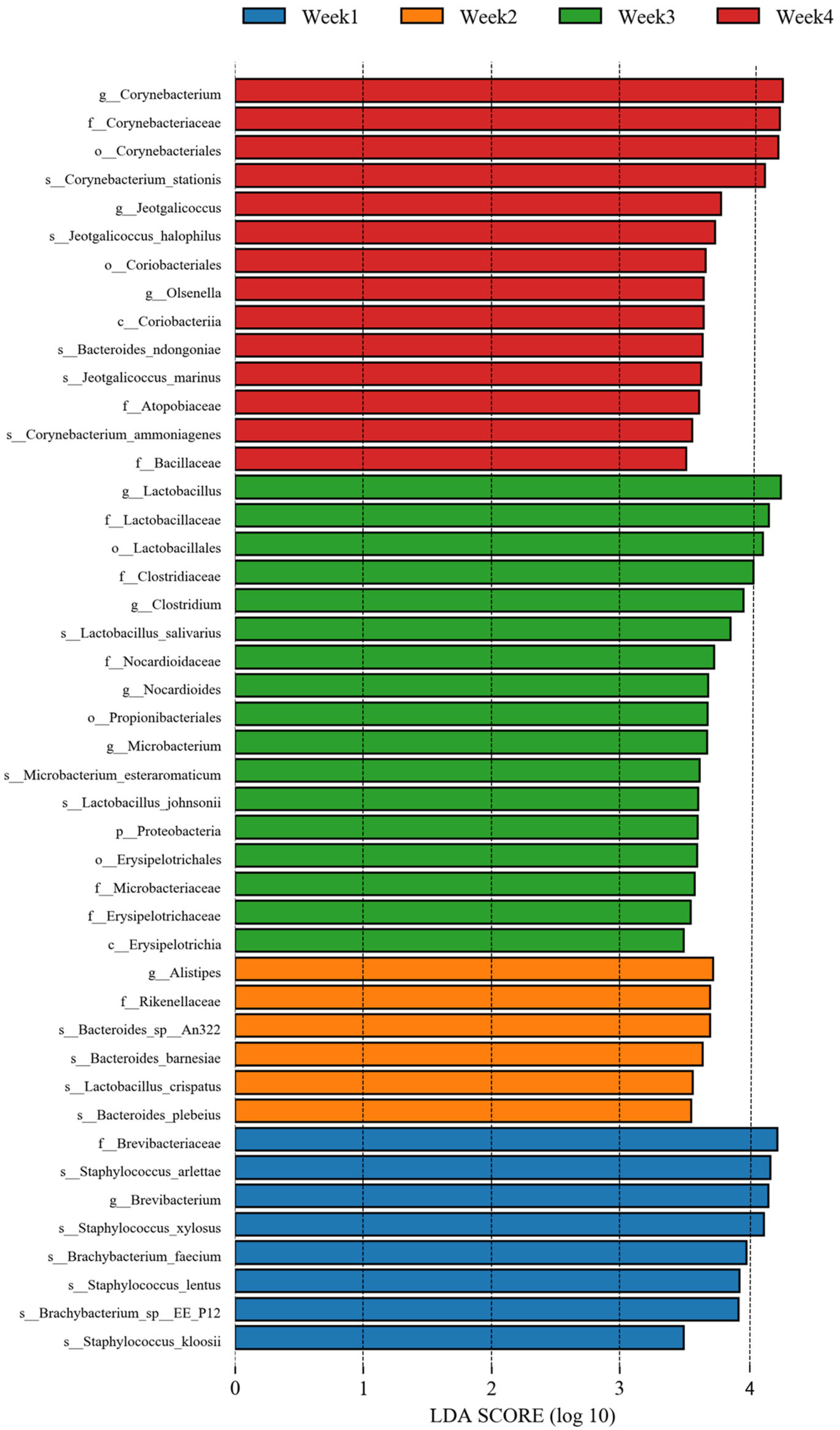
3.2. Bacterial Communities and Diversity Analysis
3.3. Opportunistic Pathogens Were Discovered in the PM2.5 Samples
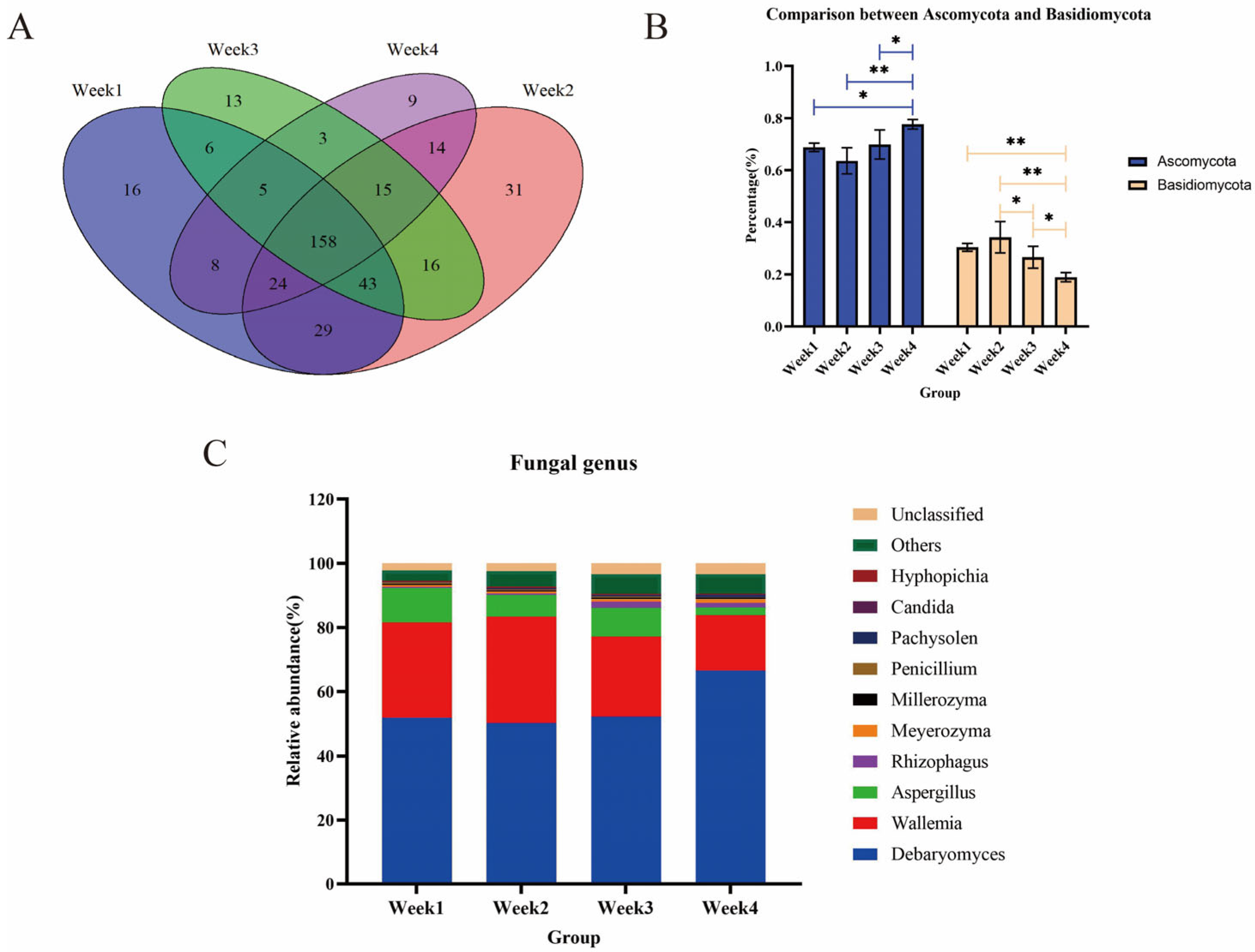
3.4. Fungal Communities and Diversity Analysis
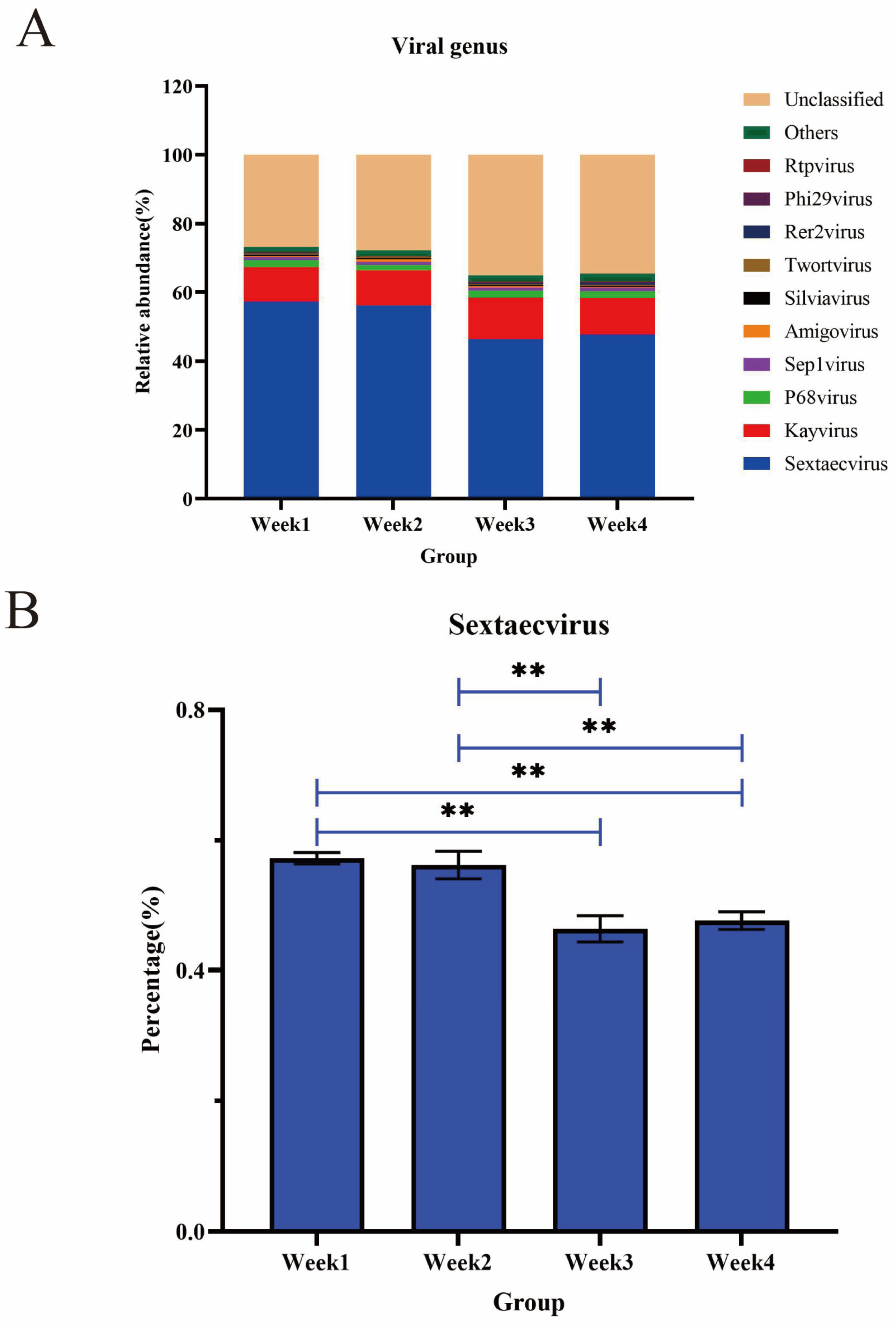
3.5. Diversity and Abundance of Viruses
3.6. Diversity and Abundance of Archaea
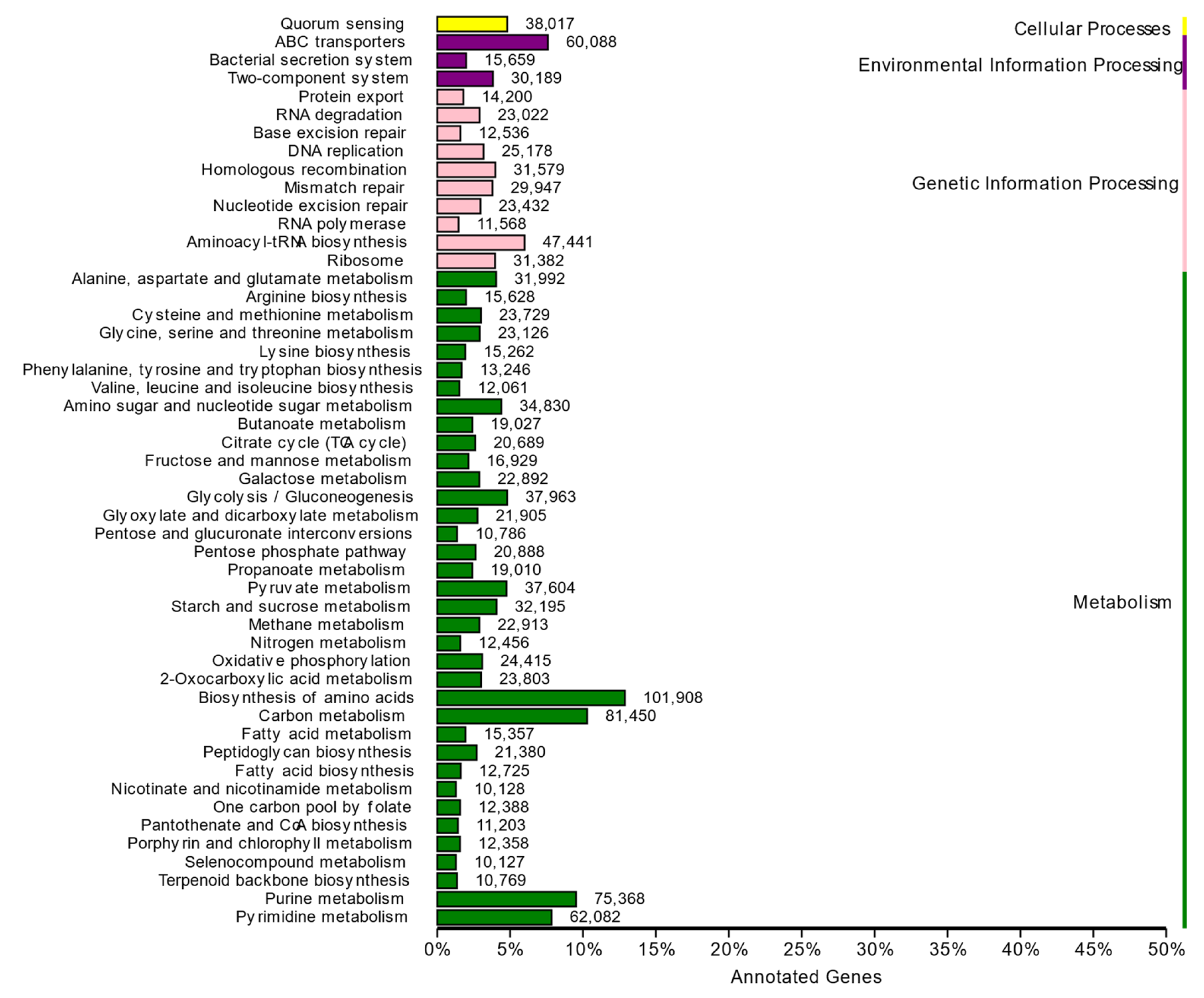
3.7. Functional Profile of the DNA Metagenome
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bai, W.; Li, Y.; Xie, W.; Ma, T.; Hou, J.; Zeng, X. Vertical variations in the concentration and community structure of airborne microbes in PM2.5. Sci. Total Environ. 2021, 760, 143396. [Google Scholar] [CrossRef]
- Carducci, A.; Donzelli, G.; Cioni, L.; Verani, M. Quantitative Microbial Risk Assessment in Occupational Settings Applied to the Airborne Human Adenovirus Infection. Int. J. Environ. Res. Public. Health 2016, 13, 733. [Google Scholar] [CrossRef]
- Song, L.; Wang, C.; Jiang, G.; Ma, J.; Li, Y.; Chen, H.; Guo, J. Bioaerosol is an important transmission route of antibiotic resistance genes in pig farms. Environ. Int. 2021, 154, 106559. [Google Scholar] [CrossRef]
- Che, F.X.; Li, J.S.; Chai, T.J. Principles and Applications of Aerobiology; Science Press: Beijing, China, 2004. [Google Scholar]
- Ashiq, S.; Hussain, M.; Ahmad, B. Natural occurrence of mycotoxins in medicinal plants: A review. Fungal Genet. Biol. 2014, 66, 1–10. [Google Scholar] [CrossRef]
- Shu, Y.; Wang, S.P.; Wu, L.Y.; Zeng, Y.; Zhou, Y.H.; Hu, X.X.; Gao, S.F.; Liu, H.Z. Review of Pathogenic Microbial Aerosol Detection and Prevention on Livestock Farms. Hunan Agric. Sci. 2014, 19, 36–38. [Google Scholar] [CrossRef]
- Yang, Y.; Zhou, R.; Chen, B.; Zhang, T.; Hu, L.; Zou, S. Characterization of airborne antibiotic resistance genes from typical bioaerosol emission sources in the urban environment using metagenomic approach. Chemosphere 2018, 213, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Hamscher, G.; Pawelzick, H.T.; Sczesny, S.; Nau, H.; Hartung, J. Antibiotics in dust originating from a pig-fattening farm: A new source of health hazard for farmers? Environ. Health Perspect. 2003, 111, 1590–1594. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.R.; Han, M.F.; Wang, C.; Yang, N.Y.; Wang, Y.C.; Duan, E.H.; Hsi, H.C.; Deng, J.G. A short review of bioaerosol emissions from gas bioreactors: Health threats, influencing factors and control technologies. Chemosphere 2020, 253, 126737. [Google Scholar] [CrossRef] [PubMed]
- Xi, M.; Shen, D.; Dai, P.; Han, G.; Li, C. TBHQ alleviates pyroptosis and necroptosis in chicken alveolar epithelial cells induced by fine particulate matter from broiler houses. Poult. Sci. 2022, 101, 101593. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Zhang, C.; Chen, S.; Shi, L.; Li, D.C.; Lv, N.; Cui, L.; Chen, Y.; Zheng, Y. Particulate Matter 2.5 Induced Developmental Cardiotoxicity in Chicken Embryo and Hatchling. Front. Pharmacol. 2020, 11, 841. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Chen, M.; Gao, Y.; Wang, Z.; Jin, F. Tussilagone protects acute lung injury from PM2.5 via alleviating Hif-1α/NF-κB-mediated inflammatory response. Environ. Toxicol. 2022, 37, 1198–1210. [Google Scholar] [CrossRef]
- Wang, Z.; Wu, Y.; Pei, C.; Wang, M.; Wang, X.; Shi, S.; Huang, D.; Wang, Y.; Li, S.; Xiao, W.; et al. Astragaloside IV pre-treatment attenuates PM2.5-induced lung injury in rats: Impact on autophagy, apoptosis and inflammation. Phytomedicine 2022, 96, 153912. [Google Scholar] [CrossRef] [PubMed]
- Arteaga, V.; Mitchell, D.; Armitage, T.; Tancredi, D.; Schenker, M.; Mitloehner, F. Cage Versus Noncage Laying-Hen Housings: Respiratory Exposures. J. Agromed. 2015, 20, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Bai, Y.; Lou, C.; Wu, B. Serum metabolome responses induced by long-term inoculation of suspended PM2.5 in chicken. Poult. Sci. 2024, 103, 103283. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Guo, M.; Li, Z.; Yu, X.; Huang, G.; Li, Z.; Zhang, X.; Liu, L. Associations between air pollutant and pneumonia and asthma requiring hospitalization among children aged under 5 years in Ningbo, 2015–2017. Front. Public Health 2022, 10, 1017105. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Routledge, M.N. The contribution of PM2.5 to cardiovascular disease in China. Environ. Sci. Pollut. Res. Int. 2020, 27, 37502–37513. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Gong, Y.Y.; Martinelli, I.; Angelici, L.; Favero, C.; Bertazzi, P.A.; Mannucci, P.M.; Ariëns, R.A.; Routledge, M.N. Fibrin clot structure is affected by levels of particulate air pollution exposure in patients with venous thrombosis. Environ. Int. 2016, 92–93, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Stafoggia, M.; Renzi, M.; Forastiere, F.; Ljungman, P.; Davoli, M.; De’ Donato, F.; Gariazzo, C.; Michelozzi, P.; Scortichini, M.; Solimini, A.; et al. Short-term effects of particulate matter on cardiovascular morbidity in Italy: A national analysis. Eur. J. Prev. Cardiol. 2022, 29, 1202–1211. [Google Scholar] [CrossRef]
- Wu, B.; Qin, L.; Wang, M.; Zhou, T.; Dong, Y.; Chai, T. The composition of microbial aerosols, PM2.5, and PM10 in a duck house in Shandong province, China. Poult. Sci. 2019, 98, 5913–5924. [Google Scholar] [CrossRef]
- Yang, W.; Guo, M.; Liu, G.; Yu, G.; Wang, P.; Wang, H.; Chai, T. Detection and analysis of fine particulate matter and microbial aerosol in chicken houses in Shandong Province, China. Poult. Sci. 2018, 97, 995–1005. [Google Scholar] [CrossRef]
- Xu, Y.; Huang, Y.; Guo, L.; Zhang, S.; Wu, R.; Fang, X.; Xu, H.; Nie, Q. Metagenomic analysis reveals the microbiome and antibiotic resistance genes in indigenous Chinese yellow-feathered chickens. Front. Microbiol. 2022, 13, 930289. [Google Scholar] [CrossRef] [PubMed]
- Rajeoni, A.H.; Ghalyanchilangeroudi, A.; Khalesi, B.; Madadi, M.S.; Hosseini, H. The tracheal virome of broiler chickens with respiratory disease complex in Iran: The metagenomics study. Iran. J. Microbiol. 2021, 13, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Kim, Y.I.; Pascua, P.N.; Choi, Y.K. Avian Influenza A Viruses: Evolution and Zoonotic Infection. Semin. Respir. Crit. Care Med. 2016, 37, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Trojánek, Z.; Kovarík, A.; Spanová, A.; Marosiová, K.; Horák, D.; Rittich, B. Application of magnetic polymethacrylate-based microspheres for the isolation of DNA from raw vegetables and processed foods of plant origin. J. Food Process. Preserv. 2018, 42, e13384. [Google Scholar] [CrossRef]
- Li, D.; Liu, C.M.; Luo, R.; Sadakane, K.; Lam, T.W. MEGAHIT: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Lomsadze, A.; Borodovsky, M. Ab initio gene identification in metagenomic sequences. Nucleic Acids Res. 2010, 38, e132. [Google Scholar] [CrossRef] [PubMed]
- Steinegger, M.; Söding, J. MMseqs2 enables sensitive protein sequence searching for the analysis of massive data sets. Nat. Biotechnol. 2017, 35, 1026–1028. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef]
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The Carbohydrate-Active EnZymes database (CAZy): An expert resource for Glycogenomics. Nucleic Acids Res. 2009, 37, D233–D238. [Google Scholar] [CrossRef]
- Chen, L.; Yang, J.; Yu, J.; Yao, Z.; Sun, L.; Shen, Y.; Jin, Q. VFDB: A reference database for bacterial virulence factors. Nucleic Acids Res. 2005, 33, D325–D328. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.Y.; Li, J.Q.; Wu, S.F.; Zhu, Y.P.; Chen, Y.W.; He, C.F. Integrated nr database in protein annotation system and its localization. Comput. Eng. 2006, 32, 71–74. [Google Scholar]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Nageshwar Reddy, D. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787–8803. [Google Scholar] [CrossRef] [PubMed]
- Qian, C.; Castañeda-Gulla, K.; Sattlegger, E.; Mutukumira, A.N. Enterotoxigenicity and genetic relatedness of Staphylococcus aureus in a commercial poultry plant and poultry farm. Int. J. Food Microbiol. 2022, 363, 109454. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.; Porter, R.; McComb, B.; Lipper, R.; Olson, S.; Nohner, S.; Shivaprasad, H.L. Clostridial dermatitis and cellulitis: An emerging disease of turkeys. Avian Dis. 2010, 54, 788–794. [Google Scholar] [CrossRef] [PubMed]
- Cleveland, K.O.; Gelfand, M.S. Crying “FOWL”: Streptococcus gallinaceous infective endocarditis and spinal infection in a chicken farmer. Am. J. Med. Sci. 2022, 364, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Zhang, J.; Tang, J.; Li, M.; Zhao, X.; Zhu, H.; Yu, X.; Li, Y.; Feng, T.; Zhang, X. Analyses of Aerosol Concentrations and Bacterial Community Structures for Closed Cage Broiler Houses at Different Broiler Growth Stages in Winter. J. Food Prot. 2018, 81, 1557–1564. [Google Scholar] [CrossRef]
- Tang, Q.; Huang, K.; Liu, J.; Shen, D.; Dai, P.; Li, Y.; Li, C. Seasonal variations of microbial assemblage in fine particulate matter from a nursery pig house. Sci. Total Environ. 2020, 708, 134921. [Google Scholar] [CrossRef]
- Astley, R.; Miller, F.C.; Mursalin, M.H.; Coburn, P.S.; Callegan, M.C. An Eye on Staphylococcus aureus Toxins: Roles in Ocular Damage and Inflammation. Toxins 2019, 11, 356. [Google Scholar] [CrossRef]
- Zhao, Y.; Tang, J.; Yang, D.; Tang, C.; Chen, J. Staphylococcal enterotoxin M induced inflammation and impairment of bovine mammary epithelial cells. J. Dairy Sci. 2020, 103, 8350–8359. [Google Scholar] [CrossRef]
- Lu, C.P. Veterinary Microbiology; China Agriculture Press: Beijing, China, 2001. [Google Scholar]
- Huang, Y.; Peng, C.; Cheng, L.; Li, W. Diagnosis and Control of Pseudo Tuberculosis in Muscovy Ducks. China Poult. 1997, 12, 19–20. [Google Scholar] [CrossRef]
- Jain, U.; Ver Heul, A.M.; Xiong, S.; Gregory, M.H.; Demers, E.G.; Kern, J.T.; Lai, C.W.; Muegge, B.D.; Barisas, D.A.G.; Leal-Ekman, J.S.; et al. Debaryomyces is enriched in Crohn’s disease intestinal tissue and impairs healing in mice. Science 2021, 371, 1154–1159. [Google Scholar] [CrossRef] [PubMed]
- Cannon, R.D.; Lamping, E.; Holmes, A.R.; Niimi, K.; Baret, P.V.; Keniya, M.V.; Tanabe, K.; Niimi, M.; Goffeau, A.; Monk, B.C. Efflux-Mediated Antifungal Drug Resistance. Clin. Microbiol. Rev. 2009, 22, 291–321. [Google Scholar] [CrossRef] [PubMed]
- Meng, K.; Yuan, X.Y.; Yu, J.; Zhang, Y.; Ai, W.; Wang, Y. Isolation and Identification of a Fowl Adenovirus Serotype 4 Strain from Chicken Broiler and Prevention and Control Effect of Tissue Inactivated Vaccine. Viruses 2021, 11, 627. [Google Scholar] [CrossRef] [PubMed]
- Saengkerdsub, S.; Ricke, S.C. Ecology and characteristics of methanogenic archaea in animals and humans. Crit. Rev. Microbiol. 2014, 40, 97–116. [Google Scholar] [CrossRef] [PubMed]
- Timmis, K.N. Handbook of Hydrocarbon and Lipid Microbiology; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Sogodogo, E.; Drancourt, M.; Grine, G. Methanogens as emerging pathogens in anaerobic abscesses. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Lepp, P.W.; Brinig, M.M.; Ouverney, C.C.; Palm, K.; Armitage, G.C.; Relman, D.A. Methanogenic Archaea and human periodontal disease. Proc. Natl. Acad. Sci. USA 2004, 101, 6176–6181. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, P.D.; Shanahan, F.; Marchesi, J.R. Human methanogen diversity and incidence in healthy and diseased colonic groups using mcrA gene analysis. BMC Microbiol. 2008, 8, 79. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Lian, Y.; Lin, D.; Huang, D.; Yao, Y.; Ju, F.; Wang, M. Insights into microbial contamination in multi-type manure-amended soils: The profile of human bacterial pathogens, virulence factor genes and antibiotic resistance genes. J. Hazard. Mater. 2022, 437, 129356. [Google Scholar] [CrossRef]
- Cross, A.S. What is a virulence factor? Crit. Care 2008, 12, 196. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Opportunistic Pathogens | Relative Abundance | Number of Species |
|---|---|---|
| Staphylococcus | 15.999% | 138 |
| Clostridium | 2.486% | 456 |
| Enterococcus | 0.497% | 80 |
| Escherichia | 0.294% | 31 |
| Campylobacter | 0.158% | 176 |
| Streptococcus | 0.097% | 137 |
| Acinetobacter | 0.090% | 60 |
| Mycobacterium | 0.024% | 207 |
| Listeria | 0.019% | 23 |
| Salmonella | 0.014% | 4 |
| Virulence Factor ID | VF’s Name | Species Name | VF’s Function | Abundance |
|---|---|---|---|---|
| VF0272 | FbpABC | Neisseria meningitidis | Encodes a periplasmic-binding protein-dependent iron transport system necessary for the utilization of iron bound to transferrin or iron chelates; FbpA is the periplasmic Fe3+ binding protein | 0.48% |
| VF0268 | HitABC | Haemophilus influenzae | The HitABC(fbpABC) operon encodes a periplasmic-binding protein-dependent iron transport system necessary for the utilization of iron bound to transferrin or iron chelates | 0.36% |
| VF0573 | Colibactin | Klebsiella pneumonia | Inducing DNA damage | 0.27% |
| VF0467 | Acinetobactin | Acinetobacter baumannii | High-affinity catechol-hydroxamate siderophore competing with host cells for iron | 0.22% |
| VF0361 | Capsule | Enterococcus faecalis | Contributes to host immune evasion | 0.22% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lou, C.; Chen, Z.; Bai, Y.; Chai, T.; Guan, Y.; Wu, B. Exploring the Microbial Community Structure in the Chicken House Environment by Metagenomic Analysis. Animals 2024, 14, 55. https://doi.org/10.3390/ani14010055
Lou C, Chen Z, Bai Y, Chai T, Guan Y, Wu B. Exploring the Microbial Community Structure in the Chicken House Environment by Metagenomic Analysis. Animals. 2024; 14(1):55. https://doi.org/10.3390/ani14010055
Chicago/Turabian StyleLou, Cheng, Zhuo Chen, Yu Bai, Tongjie Chai, Yuling Guan, and Bo Wu. 2024. "Exploring the Microbial Community Structure in the Chicken House Environment by Metagenomic Analysis" Animals 14, no. 1: 55. https://doi.org/10.3390/ani14010055
APA StyleLou, C., Chen, Z., Bai, Y., Chai, T., Guan, Y., & Wu, B. (2024). Exploring the Microbial Community Structure in the Chicken House Environment by Metagenomic Analysis. Animals, 14(1), 55. https://doi.org/10.3390/ani14010055