
Supplementation of the Combination of Quercetin and Vitamin E Alleviates the Effects of Heat Stress on the Uterine Function and Hormone Synthesis in Laying Hens

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Sample Collection
2.3. Egg Quality
2.4. Determination of Serum Biochemical Parameters, Progesterone, and Estradiol Levels
2.5. Determination of Calcium Level and Action of Ca2+-ATPase
2.6. Morphological Analysis of the Uterus Heat-Stressed Hens
2.7. Assessment of the Oxidative Status in the Shell Gland of Heat-Stressed Hens
2.8. Total RNA Extraction and qPCR
2.9. Statistical Analysis
3. Results
3.1. Effects of Quercetin (Q), Vitamin E (VE), and Combination of Quercetin and Vitamin E (Q + VE) on Production Performance of Heat-Stressed Hens
3.2. Effects of Dietary Quercetin (Q), Vitamin E (VE), and Combination of Quercetin and Vitamin E (Q + VE) on Eggshell Quality
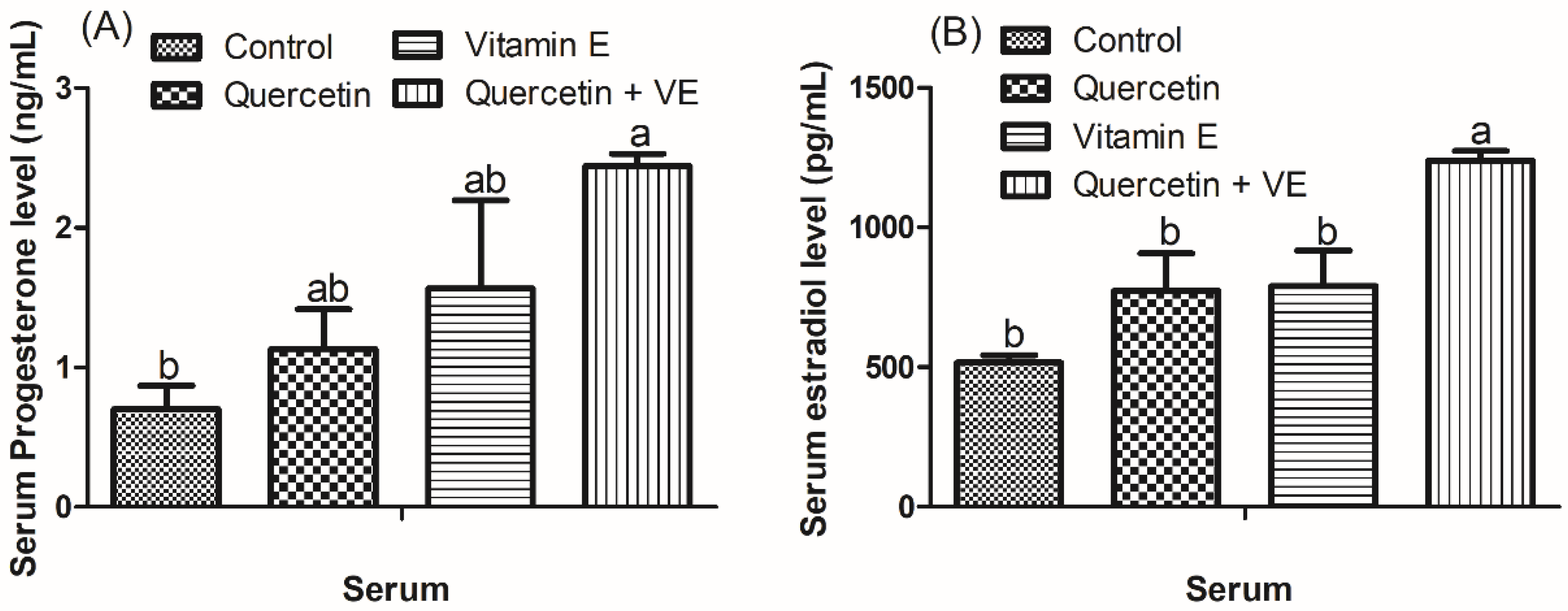
3.3. Effects of Dietary Quercetin (Q), Vitamin E (VE), and Combination of Quercetin and Vitamin E (Q + VE) on Serum Biochemical Parameters, Progesterone and Estradiol Levels
3.4. Effects of Dietary Quercetin (Q), Vitamin E (VE), and Combination of Quercetin and Vitamin E (Q + VE) on Calcium Concentration and Ca2+-ATPase Activity
3.5. Effects of Dietary Quercetin (Q), Vitamin E (VE), and Combination of Quercetin and Vitamin E (Q + VE) on Oxidative Status of the Uterus and Serum
3.6. Effects of Dietary Quercetin (Q), Vitamin E (VE), and Combination of Quercetin and Vitamin E (Q + VE) on Gene Expressions in the Uterus of Heat-Stressed Hens
3.7. Effects of Dietary Quercetin (Q), Vitamin E (VE), and Combination of Quercetin and Vitamin E (Q + VE) on Gene Expressions in the SYF, Hypothalamus, and Pituitary of Heat-Stressed Hens
3.8. Effects of Dietary Quercetin (Q), Vitamin E (VE), and Combination of Quercetin and Vitamin E (Q + VE) on the Morphological Analysis of the Uterus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tumova, E.; Gous, R.M. Interaction of hen production type, age, and temperature on laying pattern and egg quality. Poult. Sci. 2012, 91, 1269–1275. [Google Scholar] [CrossRef]
- Yardibi, H.; Turkay, G. The effects of vitamin E on antioxidant system and egg quality in heat stressed laying hens. Turk. J. Vet. Anim. Sci. 2014, 32, 319–325. [Google Scholar]
- Liu, M.; Lu, Y.; Gao, P.; Xie, X.; Li, D.; Yu, D.; Yu, M. Effect of curcumin on laying performance, egg quality, endocrine hormones, and immune activity in heat-stressed hens. Poult. Sci. 2020, 99, 2196–2202. [Google Scholar] [CrossRef]
- Mashaly, M.M.; Hendricks, G.L., 3rd; Kalama, M.A.; Gehad, A.E.; Abbas, A.O.; Patterson, P.H. Effect of heat stress on production parameters and immune responses of commercial laying hens. Poult. Sci. 2004, 83, 889–894. [Google Scholar] [CrossRef]
- Sangoh, P.H.; Jong, R.; Chaemin, P.; Byungsung, C.; Hyunseok, C.; Heechul, H.K.; Kang, S.O.; Yangho, C. Effects of extreme heat stress on growth performance, lymphoid organ, IgG and cecum microflora of broiler chickens. Int. J. Agric. Biol. 2013, 15, 1204–1208. [Google Scholar]
- Deeb, N.; Cahaner, A. Genotype-by-environment interaction with broiler genotypes differing in growth rate. Growth rate and water consumption of broiler progeny from weight-selected versus nonselected parents under normal and high ambient temperatures. Poult. Sci. 2002, 81, 293–301. [Google Scholar] [CrossRef]
- Zhang, M.; Zou, X.T.; Li, H.; Dong, X.Y.; Zhao, W. Effect of dietary γ-aminobutyric acid on laying performance, egg quality, immune activity and endocrine hormone in heat-stressed Roman hens. Anim. Sci. J. 2012, 83, 141–147. [Google Scholar] [CrossRef]
- Settar, P.; Yalcin, S.; Turkmut, L.; Ozkan, S.; Cahanar, A. Season by genotype interaction related to broiler growth rate and heat tolerance. Poult. Sci. 1999, 78, 1353–1358. [Google Scholar] [CrossRef]
- Lara, L.J.; Rostagno, M.H. Impact of Heat Stress on Poultry Production. Animals 2013, 3, 356–369. [Google Scholar] [CrossRef]
- Lin, H.; Mertens, K.; Kemps, B.; Govaerts, T.; De Ketelaere, B.; De Baerdemaeker, J.; Decuypere, E.; Buyse, J. New approach of testing the effect of heat stress on eggshell quality: Mechanical and material properties of eggshell and membrane. Br. Poult. Sci. 2004, 45, 476–482. [Google Scholar] [CrossRef]
- Lin, H.; Decuypere, E.; Buyse, J. Acute heat stress induces oxidative stress in broiler chickens. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2006, 144, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Qi, L.; Dai, H.; Hu, C.; Lv, Z.; Wei, Q.; Shi, F. Dietary stevioside supplementation improves laying performance and eggshell quality through increasing estrogen synthesis, calcium level and antioxidant capacity of reproductive organs in aged breeder hens. Anim. Feed. Sci. Technol. 2020, 269, 114682. [Google Scholar] [CrossRef]
- Li, Q.; Zhao, X.; Wang, S.; Zhou, Z. Letrozole induced low estrogen levels affected the expressions of duodenal and renal calcium-processing gene in laying hens. Gen. Comp. Endocrinol. 2018, 255, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Rivas, P.A.; Chauhan, S.S.; Ha, M.; Fegan, N.; Dunshea, F.R.; Warner, R.D. Effects of heat stress on animal physiology, metabolism, and meat quality: A review. Meat Sci. 2020, 162, 108025. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Y.; Liu, H.N.; Suo, Y.L.; Hu, L.L.; Feng, X.A.; Zhang, L.; Jin, F. Effect of quercetin on performance and egg quality during the late laying period of hens. Br. Poult. Sci. 2013, 54, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Dalia, A.M.; Loh, T.C.; Sazili, A.Q.; Jahromi, M.F.; Samsudin, A.A. Effects of vitamin E, inorganic selenium, bacterial organic selenium, and their combinations on immunity response in broiler chickens. BMC Vet. Res. 2018, 24, 249. [Google Scholar] [CrossRef] [PubMed]
- Amevor, F.K.; Cui, Z.; Du, X.; Ning, Z.; Shu, G.; Jin, N.; Deng, X.; Tian, Y.; Zhang, Z.; Kang, X.; et al. Combination of quercetin and vitamin E supplementation promotes yolk precursor synthesis and follicle development in aging breeder hens via liver-blood-ovary signal axis. Animals 2021, 11, 1915. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Zhao, X.; Amevor, F.K.; Du, X.; Wang, Y.; Li, D.; Shu, G.; Tian, Y.; Zhao, X. Therapeutic application of quercetin in aging-related diseases: SIRT1 as a potential mechanism. Front. Immunol. 2022, 13, 943321. [Google Scholar] [CrossRef] [PubMed]
- Amevor, F.K.; Cui, Z.; Ning, Z.; Du, X.; Jin, N.; Shu, G.; Deng, X.; Zhu, Q.; Tian, Y.; Li, D.; et al. Synergistic effects of quercetin and vitamin E on egg production, egg quality, and immunity in aging breeder hens. Poult. Sci. 2021, 100, 101481. [Google Scholar] [CrossRef]
- Liu, Y.J.; Zhao, L.H.; Mosenthin, R.; Zhang, J.Y.; Ji, C.; Ma, Q.G. Protective effect of vitamin E on laying performance, antioxidant capacity, and immunity in laying hens challenged with Salmonella Enteritidis. Poult. Sci. 2019, 98, 5847–5854. [Google Scholar] [CrossRef]
- Jiang, W.; Zhang, L.; Shan, A. The effect of vitamin E on laying performance and egg quality in laying hens fed corn dried distillers grains with solubles. Poult. Sci. 2013, 92, 2956–2964. [Google Scholar] [CrossRef] [PubMed]
- Habibian, M.; Ghazi, S.; Moeini, M.M.; Abdolmohammadi, A. Effects of dietary selenium and vitamin E on immune response and biological blood parameters of broilers reared under thermoneutral or heat stress conditions. Int. J. Biometeorol. 2014, 58, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Sonam, K.; Guleria, S. Synergistic antioxidant activity of natural products. Annal. Pharmacol. Pharm. 2017, 2, 1086. [Google Scholar]
- McDowell, L.R. Vitamins in Animal Nutrition: Comparative Aspects to Human Nutrition; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Cui, Z.; Jin, N.; Amevor, F.K.; Shu, G.; Du, X.; Kang, X.; Ning, Z.; Deng, X.; Tian, Y.; Zhu, Q.; et al. Dietary supplementation of salidroside alleviates liver lipid metabolism disorder and inflammatory response to promote hepatocyte regeneration via PI3K/AKT/Gsk3-beta pathway. Poult. Sci. 2022, 101, 102034. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hu, Z.; Lu, C.; Bai, K.; Zhang, L.; Wang, T. Effect of various levels of dietary curcumin on meat quality and antioxidant profile of breast muscle in broilers. J. Agric. Food Chem. 2015, 63, 3880–3886. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.X.; Maria, T.C.; Zhou, B.; Xiao, F.L.; Wang, M.; Mao, Y.J.; Li, Y. Quercetin improves immune function in Arbor Acre broilers through activation of NF-κB signaling pathway. Poult. Sci. 2020, 99, 906–913. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ding, X.; Bai, S.; Wang, J.; Zeng, Q.; Peng, H.; Su, Z.; Xuan, Y.; Fraley, G.S.; Zhang, K. Effects of maternal dietary vitamin E on the egg characteristics, hatchability and offspring quality of prolonged storage eggs of broiler breeder hens. J. Anim. Physiol. Anim. Nutr. 2018, 104, 1384–1391. [Google Scholar] [CrossRef]
- Cui, Z.; Ning, Z.; Deng, X.; Du, X.; Amevor, F.K.; Liu, L.; Kang, X.; Tian, Y.; Wang, Y.; Li, D.; et al. Integrated proteomic and metabolomic analyses of chicken ovary revealed the crucial role of lipoprotein lipase on lipid metabolism and steroidogenesis during sexual maturity. Front. Physiol. 2022, 13, 885030. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using RT- PCR and the 2 (-Delta C (T)) methods. Methods 2001, 25, 402–440. [Google Scholar] [CrossRef]
- Ghasemian, M.; Jahanian, R. Dietary mannan-oligosaccharides supplementation could affect performance, immunocompetence, serum lipid metabolites, intestinal bacterial populations, and ileal nutrient digestibility in aged laying hens. Anim. Feed Sci. Technol. 2016, 213, 81–89. [Google Scholar] [CrossRef]
- Miao, L.P.; Zhou, M.Y.; Zhang, X.Y.; Yuan, C.; Dong, X.Y.; Zou, X.T. Effect of excess dietary fluoride on laying performance and antioxidant capacity of laying hens. Poult. Sci. 2017, 96, 2200–2205. [Google Scholar] [CrossRef] [PubMed]
- Vimalraj, S. Alkaline phosphatase: Structure, expression and its function in bone mineralization. Gene 2020, 754, 144855. [Google Scholar] [CrossRef] [PubMed]
- Dobson, H.; Smith, R.F. Stress and reproduction in farm animals. J. Reprod. Fertil. Suppl. 1995, 49, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Wang, Z.; Cao, J.; Chen, Y.; Dong, Y. A novel and compact review on the role of oxidative stress in female reproduction. Reprod. Biol. Endocrinol. 2018, 16, 80. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.X.; Chaudhry, M.T.; Yao, J.Y.; Wang, S.N.; Zhou, B.; Wang, M.; Han, C.Y.; You, Y.; Li, Y. Effects of phyto-oestrogen quercetin on productive performance, hormones, reproductive organs and apoptotic genes in laying hens. J. Anim. Physiol. Anim. Nutr. 2018, 102, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Ulusoy, H.G.; Sanlier, N. A minireview of quercetin: From its metabolism to possible mechanisms of its biological activities. Crit. Rev. Food Sci. Nutr. 2020, 60, 3290–3303. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Qi, L.; Lv, Z.; Jin, S.; Wei, X.; Shi, F. Dietary stevioside supplementation alleviates lipopolysaccharide-induced intestinal mucosal damage through anti-inflammatory and antioxidant effects in broiler chickens. Antioxidants 2019, 8, 575. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Cheng, L.; Li, X.; Tang, S.; Zhang, X.; Gong, Y. Transcriptional regulation of CYP19A1 expression in chickens: ESR1, ESR2 and NR5A2 form a functional network. Gen. Comp. Endocrinol. 2022, 315, 113939. [Google Scholar] [CrossRef] [PubMed]
- Kakuta, H.; Iguchi, T.; Sato, T. The Involvement of Granulosa Cells in the Regulation by Gonadotropins of Cyp17a1 in Theca Cells. In Vivo 2018, 32, 1387–1401. [Google Scholar] [CrossRef]
- Wang, C.; Ma, W. Hypothalamic and pituitary transcriptome profiling using RNA-sequencing in high-yielding and low-yielding laying hens. Sci. Rep. 2019, 9, 10285. [Google Scholar] [CrossRef]
- Kim, D.; Ocon-Grove, O.; Johnson, A.L. Bone morphogenetic protein 4 supports the initial differentiation of hen (Gallus gallus) granulosa cells. Biol. Reprod. 2013, 88, 161. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lin, X.; Zhang, S.; Guo, C.; Li, J.; Mi, Y.; Zhang, C. Lycopene ameliorates oxidative stress in the aging chicken ovary via activation of Nrf2/HO-1 pathway. Aging 2018, 10, 2016–2036. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Yao, J.; Zhou, S.; Mi, Y.; Li, J.; Zhang, C. Improvement of eggshell quality by dietary N-carbamylglutamate supplementation in laying chickens. Poult. Sci. 2020, 99, 4085–4095. [Google Scholar] [CrossRef] [PubMed]
- Sack, M.N.; Fyhrquist, F.Y.; Saijonmaa, O.J.; Fuster, V.; Kovacic, J.C. Basic biology of oxidative stress and the cardiovascular system: Part 1 of a 3-part series. J. Am. Coll. Cardiol. 2017, 70, 196–211. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, K.Z.; Beck, M.M.; Scheideler, S.E.; Forman, M.F.; Anderson, K.P.; Kachman, S.D. Acute high environmental temperature and calcium-estrogen relationship in the hen. Poult. Sci. 1996, 75, 1555–1562. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.T.; Fujita, M.; Yamamoto, S.; Iwasaki, K.; Ikawa, R.; Oyama, H.; Horikawa, H. Effects of glucose in drinking water on the changes in whole blood viscosity and plasma osmolality of broiler chickens during high temperature exposure. Poult. Sci. 1998, 77, 644–647. [Google Scholar] [CrossRef] [PubMed]
- Onagbesan, O.M.; Peddie, M.J. Calcium-dependent stimulation of estrogen secretion by FSH from theca cells of the domestic hen (Gallus domesticus). Gen. Comp. Endocrinol. 1989, 75, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Zhao, F.; Tian, Z.M.; Zhang, H.X.; Ruan, D.; Li, Y.; Wang, S.; Zheng, C.T.; Lin, Y.C. Dietary calcium deficiency in laying ducks impairs eggshell quality by suppressing shell biomineralization. J. Exp. Biol. 2015, 218, 3336–3343. [Google Scholar] [CrossRef]
- Jing, X.; Wang, Y.; Song, F.; Xu, X.; Liu, M.; Wei, Y.; Zhu, H.; Liu, Y.; Wei, J.; Xu, X. A Comparison between Vitamin D3 and 25-Hydroxyvitamin D3 on Laying Performance, Eggshell Quality and Ultrastructure, and Plasma Calcium Levels in Late Period Laying Hens. Animals 2022, 12, 2824. [Google Scholar] [CrossRef]
- Long, L.; Wu, S.G.; Yuan, F.; Zhang, H.J.; Wang, J.; Qi, G.H. Effects of dietary octacosanol supplementation on laying performance, egg quality, serum hormone levels, and expression of genes related to the reproductive axis in laying hens. Poult. Sci. 2017, 96, 894–903. [Google Scholar] [CrossRef]
- Bar, A.; Vax, E.; Hunziker, W.; Halevy, O.; Striem, S. The role of gonadal hormones in gene expression of calbindin (Mr28,000) in the laying hen. Gen. Comp. Endocrinol. 1996, 103, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Wistedt, A.; Ridderstråle, Y.; Wall, H.; Holm, L. Exogenous estradiol improves shell strength in laying hens at the end of the laying period. Acta Vet. Scand. 2014, 56, 34. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′–3′) | Product Length (bp) | Annealing Temperature (°C) |
|---|---|---|---|
| FSHR | F: GAGGAGGTGAAGAAGATGCGGATG R: TGAGCCACTCTGTTGCCATACC | 105 | 60 |
| CYP17A1 | F: CTTCAGGTGTTTCTCTTCCTCCTC R: CTGTGGTTTCATGGCTGGATC | 131 | 59.82 |
| CYP19A1 | F: TCATCGCCTCCATCGTCTAC R: TCTTACTGCGCGTCTTCTGG | 240 | 57.79 |
| 17β-HSD | F: CCCTCACCCAGCCCGACTTC R: GCCGTTGGTGGAGGTGTTACAG | 179 | 58 |
| ERβ | F: GGCAAGCAGCACGGTGGAC R: CTTCTGCCACTCCTCCCTTTGC | 129 | 59 |
| ERα | F: ACGGCACCAACGAGGAGATCC R: CTTCCCGTTCACCTGGCACTTC | 175 | 60.67 |
| GNRHR | F: TGTGCTGTGTGCAACGACTA R: CAGGCCTGGCAACTCTTTCT | 167 | 57 |
| GNRH1 | F: CTGCAGGACGAGATGTGCAA R: AGGTCTGAAAGGCGAACAGG | 175 | 60.67 |
| CABP-28 K | F: ACATCCAGGGAGAGGTTTCCT R: GTGGGACATGGTGCCTTGAG | 208 | 60.20 |
| CDH6 | F: ATCGTCGCCTTCTTCGAGTT R: ATCCCATCCTCCGTTGTCCT | 150 | 59 |
| PR | F: GTGATGGCATGGGACATAGCTC R: TGGCGTAGACCTTGCGGATAA | 90 | 58 |
| HPRT1 | F: GTGATGGCATGGGACATAGCTC R: TGGCGTAGACCTTGCGGATAA | 90 | 58 |
| β-actin | F: ATCCGGACCCTCCATTGTC R: AGCCATGCCAATCTCGTCTT | 120 | 60 |
| Item 1 | Control | Quercetin | Vitamin E | Q + VE |
|---|---|---|---|---|
| GLU (mmol/L) | 16.15 ± 2.09 a | 14.51 ± 2.68 ab | 13.15 ± 1.79 bc | 11.99 ± 1.76 c |
| ALP (U/L) | 295.93 ± 65.02 a | 234.09 ± 66.30 b | 179.15 ± 79.94 bc | 132.99 ± 42.57 c |
| Parameters 1 | Treatments | ||||
|---|---|---|---|---|---|
| Tissue | Control | Q | VE | Q + VE | |
| SOD | Serum | 17.53 ± 3.09 c | 20.48 ± 12.14 b | 16.77 ± 2.53 c | 33.53 ± 10.07 a |
| Uterus | 24.20 ± 5.61 c | 32.23 ± 7.61 ab | 30.14 ± 7.14 ab | 35.01 ± 6.22 a | |
| TAOC | Serum | 14.81 ± 4.13 b | 20.18 ± 6.18 a | 19.70 ± 4.76 a | 24.27 ± 6.19 a |
| Uterus | 11.58 ± 2.49 c | 19.21 ± 6.28 ab | 19.49 ± 6.12 ab | 22.08 ± 11.27 a | |
| GSH-Px | Serum | 29.87 ± 7.58 c | 39.60 ± 5.13 b | 36.86 ± 6.35 b | 47.73 ± 6.81 a |
| Uterus | 18.78 ± 3.65 c | 23.83 ± 7.68 b | 24.63 ± 5.23 ab | 29.21 ± 4.20 a | |
| CAT | Serum | 16.12 ± 3.64 c | 22.01 ± 4.99 b | 22.68 ± 8.05 b | 29.20 ± 8.11 a |
| Uterus | 29.25 ± 10.62 c | 43.96 ± 3.66 b | 49.49 ± 5.40 ab | 54.34 ± 5.59 a | |
| MDA | Serum | 1.80 ± 0.59 a | 1.23 ± 0.52 b | 1.11 ± 0.53 c | 1.20 ± 0.64 b |
| Uterus | 1.86 ± 0.53 a | 0.99 ± 0.61 b | 0.91 ± 0.43 b | 0.57 ± 0.17 c | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, X.; Amevor, F.K.; Du, X.; Wu, Y.; Xu, D.; Wei, S.; Shu, G.; Feng, J.; Zhao, X. Supplementation of the Combination of Quercetin and Vitamin E Alleviates the Effects of Heat Stress on the Uterine Function and Hormone Synthesis in Laying Hens. Animals 2024, 14, 1554. https://doi.org/10.3390/ani14111554
Cao X, Amevor FK, Du X, Wu Y, Xu D, Wei S, Shu G, Feng J, Zhao X. Supplementation of the Combination of Quercetin and Vitamin E Alleviates the Effects of Heat Stress on the Uterine Function and Hormone Synthesis in Laying Hens. Animals. 2024; 14(11):1554. https://doi.org/10.3390/ani14111554
Chicago/Turabian StyleCao, Xueqing, Felix Kwame Amevor, Xiaxia Du, Youhao Wu, Dan Xu, Shuo Wei, Gang Shu, Jing Feng, and Xiaoling Zhao. 2024. "Supplementation of the Combination of Quercetin and Vitamin E Alleviates the Effects of Heat Stress on the Uterine Function and Hormone Synthesis in Laying Hens" Animals 14, no. 11: 1554. https://doi.org/10.3390/ani14111554
APA StyleCao, X., Amevor, F. K., Du, X., Wu, Y., Xu, D., Wei, S., Shu, G., Feng, J., & Zhao, X. (2024). Supplementation of the Combination of Quercetin and Vitamin E Alleviates the Effects of Heat Stress on the Uterine Function and Hormone Synthesis in Laying Hens. Animals, 14(11), 1554. https://doi.org/10.3390/ani14111554