
Effects of Acetic Acid and Morphine in Shore Crabs, Carcinus maenas: Implications for the Possibility of Pain in Decapods


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Collection and Care
2.2. Experimental Treatments
2.2.1. First Treatment
2.2.2. Second Treatment
2.3. Statistical Methods
2.3.1. First Observation Period
2.3.2. Second Observation Period
2.4. Ethical Considerations
3. Results
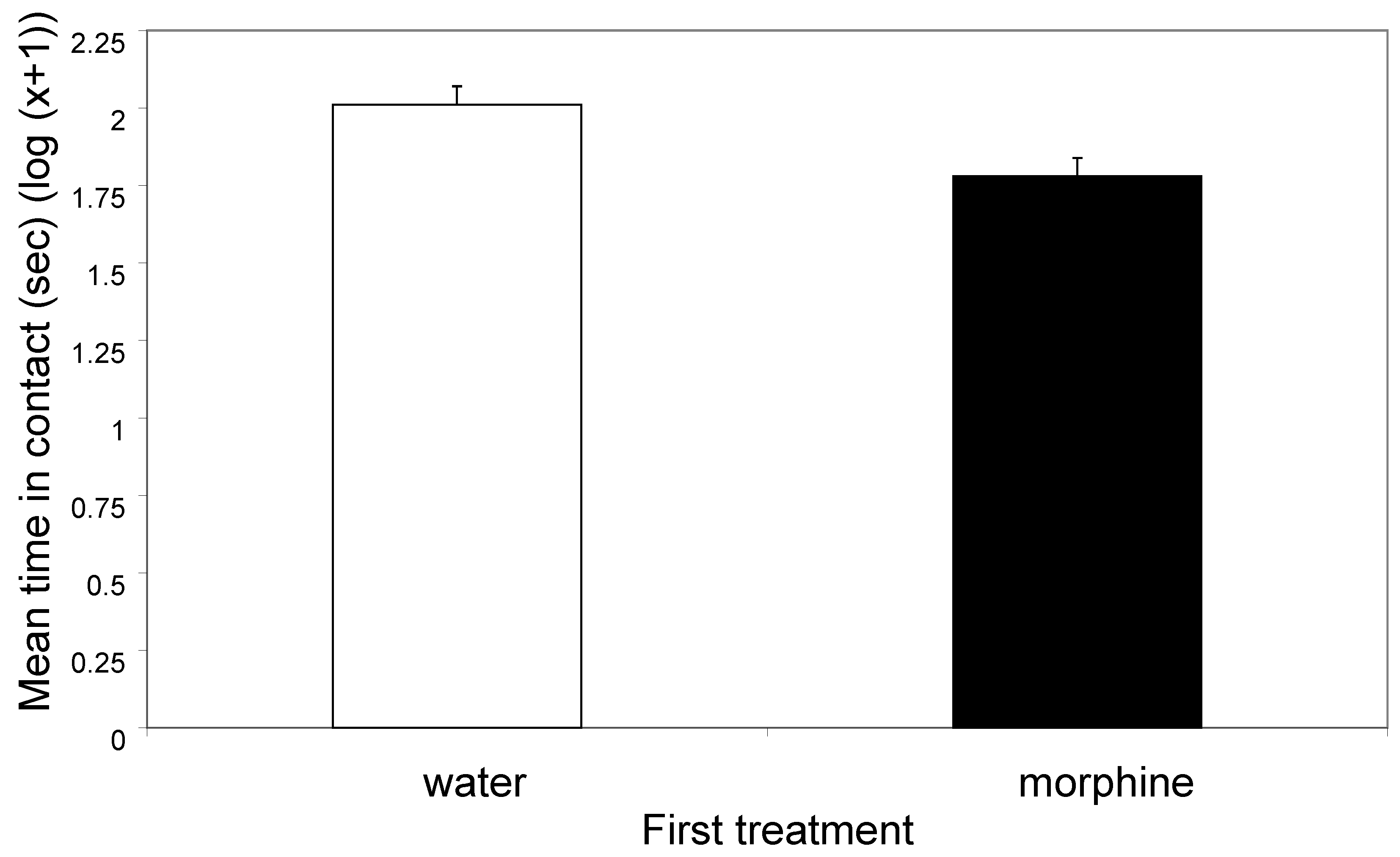
3.1. First Observation Period
3.2. Second Observation Period
4. Discussion
4.1. Effects of Morphine on Behaviour
4.2. Effects of Acetic Acid
4.3. Effects of Morphine on Responses to Acetic Acid
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gregory, N.S.; Harris, A.L.; Robinson, C.R.; Dougherty, P.M.; Fuchs, P.N.; Sluka, K.A. An overview of animal models of pain: Disease models and outcome measures. J. Pain 2013, 14, 1255–1269. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Park, J.; Kim, M.S.; Seol, G.H.; Min, S.S. Analgesic effects of eucalyptus essentialoil in mice. Korean J. Pain 2019, 32, 79–86. [Google Scholar] [CrossRef]
- Sneddon, L.U. The evidence for pain in fish: The use of morphine as an analgesic. Appl. Anim. Behav. Sci. 2003, 83, 153–162. [Google Scholar] [CrossRef]
- Elwood, R.W. Potential pain in fish and decapods: Similar experimental approaches and similar results. Front. Vet. Sci. 2021, 8, 631151. [Google Scholar] [CrossRef]
- Barr, S.; Laming, P.R.; Dick, J.T.; Elwood, R.W. Nociception or pain in a decapod crustacean? Anim. Behav. 2008, 75, 745–751. [Google Scholar] [CrossRef]
- Puri, S.; Faulkes, Z. Do decapod crustaceans have nociceptors for extreme pH? PLoS ONE 2010, 5, e10244. [Google Scholar] [CrossRef]
- Elwood, R.W.; Dalton, N.; Riddell, G. Aversive responses by shore crabs to acetic acid but not to capsaicin. Behav. Process. 2017, 140, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Barr, S.; Elwood, R.W. The Effects of Caustic Soda and Benzocaine on Directed Grooming to the Eyestalk in the Glass Prawn, Palaemon elegans, Are Consistent with the Idea of Pain in Decapods. Animals 2024, 14, 364. [Google Scholar] [CrossRef] [PubMed]
- Dyuizen, I.V.; Kotsyuba, E.P.; Lamash, N.E. Changes in the nitric oxide system in the shore crab Hemigrapsus sanguineus (Crustacea, decapoda) CNS induced by a nociceptive stimulus. J. Exp. Biol. 2012, 215, 2668–2676. [Google Scholar] [CrossRef]
- Sherwin, C.M. Can invertebrates suffer? Or how robust is argument-by-analogy? Anim. Welf. 2001, 10, 103–108. [Google Scholar] [CrossRef]
- Oliverio, A.; Castellano, C.; Puglisi-Allegra, S. Psychobiology of Opioids. Int. Rev. Neurobiol. 1984, 25, 277–337. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, F.R.; Castellano, C. Behavioral effects of morphine in mice: Role of experimental housing. Pharmacol. Biochem. Behav. 1989, 34, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Maldonaldo, H.; Miralto, A. Effect of morphine and naloxone on a defensive response of the mantis shrimp (Squilla mantis). J. Comp. Physiol. 1982, 147, 455–459. [Google Scholar] [CrossRef]
- Lozada, M.; Romano, A.; Maldonado, H. Effect of morphine and naloxone on a defensive response of the crab Chasmagnathus granulatus. Pharm. Biochem. Behav. 1988, 30, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Tomsic, T.M.V.; Maldonado, H. Habituation to a danger stimulus in two semiterrestrial crabs: Ontogenic, ecological and opioid modulation correlates. J. Comp. Physiol. A 1993, 173, 621–633. [Google Scholar] [CrossRef]
- Barr, S.; Elwood, R.W. No evidence of morphine analgesia to noxious shock in the shore crab, Carcinus maenas. Behav. Process. 2011, 86, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Patterson, L.; Dick, J.T.A.; Elwood, R.W. Claw loss and feeding ability in the edible crab, Cancer pagurus: Implications of fishery practice. Appl. Anim. Behav. Sci. 2009, 116, 302–305. [Google Scholar] [CrossRef]
- Soulsbury, C.; Bateson, M.; Braithwaite, V.; Cotter, S.; Elwood, R.W.; Whiting, M.; Wilkinson, A.; Collins, L.M. The welfare and ethics of research involving wild animals: A primer. Methods Ecol. Evol. 2020, 11, 1164–1181. [Google Scholar] [CrossRef]
- Crump, A.; Fischer, B.; Arnott, G.; Birch, J.; Briffa, M.; Browning, H.; Coates, C.; Elwood, R.; Khan, N.; Thakrar, U.; et al. Guidelines for Protecting and Promoting Decapod Crustacean Welfare in Research; Insect Welfare Research Society: Indianapolis, IN, USA, 2024; Available online: https://www.insectwelfare.com/research-guidelines (accessed on 14 February 2024).
- Nathaniel, T.I.; Panksepp, J.; Huber, R. Effects of a single and repeated morphine treatment on conditioned and unconditioned behavioral sensitization in crayfish. Behav. Brain Res. 2010, 207, 310–320. [Google Scholar] [CrossRef]
- de Souza Valente, C. Anaesthesia of decapod crustaceans. Vet. Anim. Sci. 2022, 16, 100252. [Google Scholar] [CrossRef]
- Elwood, R.W. Pain and Suffering in Invertebrates? ILAR J. 2011, 52, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Elwood, R.W. Discrimination between nociceptive reflexes and more complex responses consistent with pain in crustaceans. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20190368. [Google Scholar] [CrossRef] [PubMed]
- Crump, A.; Browning, H.; Schnell, A.; Burn, C.; Birch, J. Sentience in decapod crustaceans: A general framework and review of the evidence. Anim. Sentience 2022, 7, 1. [Google Scholar] [CrossRef]
- Maginnis, T.L. The costs of autotomy and regeneration in animals: A review and framework for future research. Behav. Ecol. 2006, 17, 857–872. [Google Scholar] [CrossRef]
- Fiorito, G. Is there ‘pain’ in invertebrates? Behav. Process. 1986, 12, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Patterson, L.; Dick, J.T.A.; Elwood, R.W. Physiological stress responses in the edible crab, Cancer pagurus to the fishery practice of de-clawing. Mar. Biol. 2007, 152, 265–272. [Google Scholar] [CrossRef]
- Sy, C.; Airriess, C.N. Dopamine–Stimulated Limb Autotomy in the Dungeness Crab, Cancer magister. J. Crust. Biol. 2002, 22, 697–703. [Google Scholar] [CrossRef]
- Elwood, R.W. Behavioural Indicators of Pain and Suffering in Arthropods and Might Pain Bite Back? Animals 2023, 13, 2602. [Google Scholar] [CrossRef]
- Eisner, T.; Camazine, S. Spider leg autotomy induced by prey venom injection: An adaptive response to “pain”? Proc. Natl. Acad. Sci. USA 1983, 80, 3382–3385. [Google Scholar] [CrossRef]
- McVean, A. The nervous control of autotomy in Carcinus maenas. J. Exp. Biol. 1974, 60, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Findlay, I.; McVean, A.R. The nervous control of limb autotomy in the hermit crab Pagurus bernhardus (L.) and the role of the cuticular stress detector, CSD1. J. Exp. Biol. 1977, 70, 93–104. [Google Scholar] [CrossRef]
- Darnell, M.Z.; Patricia, R.Y.; Backwell, P.R.Y.; Munguia, P. Frequency and latency of autotomy of a sexually selected fiddler crab appendage. J. Exp. Mar. Biol. Ecol. 2020, 523, 151255. [Google Scholar] [CrossRef]
- Farnsworth, K.D.; Elwood, R.W. Why it hurts: With freedom comes the biological need for pain. Anim. Cogn. 2023, 26, 1259–1275. [Google Scholar] [CrossRef]
- Rotllant, G.; Llonch, P.; García del Arco, J.A.; Chic, Ò.; Flecknell, P.; Sneddon, L.U. Methods to Induce Analgesia and Anesthesia in Crustaceans: A Supportive Decision Tool. Biology 2023, 12, 387. [Google Scholar] [CrossRef]
- Weary, D.M.; Neil, L.; Flower, F.C.; Fraser, D. Identifying and preventing pain in animals. Appl. Anim. Behav. Sci. 2006, 100, 64–76. [Google Scholar] [CrossRef]
- Munro, G. Pharmacological assessment of the rat formalin test utilizing the clinically used analgesic drugs gabapentin, lamotrigine, morphine, duloxetine, tramadol and ibuprofen: Influence of low and high formalin concentrations. Eur. J. Pharmacol. 2009, 605, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Magee, B.; Elwood, R.W. Shock avoidance by discrimination learning in the shore crab (Carcinus maenas) is consistent with a key criterion for pain. J. Exp. Biol. 2013, 216, 353–358. [Google Scholar] [CrossRef]
- Barr, S.; Elwood, R.W. Trade-Offs between Avoidance of Noxious Electric Shock and Avoidance of Bright Light in Shore Crabs Are Consistent with Predictions of Pain. Animals 2024, 14, 770. [Google Scholar] [CrossRef]
- Okada, S.; Hirano, N.; Abe, T.; Nagayama, T. Aversive operant conditioning alters the phototactic orientation of the marbled crayfish. J. Exp. Biol. 2021, 224 Pt 6, jeb242180. [Google Scholar] [CrossRef] [PubMed]
- Appel, M.; Elwood, R.W. Motivational trade-offs and potential pain experience in hermit crabs. Appl. Anim. Behav. Sci. 2009, 119, 120–124. [Google Scholar] [CrossRef]
- Elwood, R.W.; Appel, M. Pain experience in hermit crabs? Anim. Behav. 2009, 77, 1243–1246. [Google Scholar] [CrossRef]
- Magee, B.; Elwood, R.W. Trade-offs between predator avoidance and electric shock avoidance in hermit crabs demonstrate a non-reflexive response to noxious stimuli consistent with prediction of pain. Behav. Process. 2016, 130, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Fossat, P.; Bacqué-Cazenave, J.; De Deurwaerdère, P.; Delbecque, J.-P.; Cattaert, D. Anxiety-like behavior in crayfish is controlled by serotonin. Science 2014, 344, 1293–1297. [Google Scholar] [CrossRef] [PubMed]
- Fossat, P.; Bacqué-Cazenave, J.; De Deurwaerdere, P.; Cattaert, D.; Delbecque, J.-P. Serotonin, but not dopamine, controls the stress response and anxiety-like behavior in the crayfish Procambarus clarkii. J. Exp. Biol. 2015, 218, 2745–2752. [Google Scholar] [CrossRef] [PubMed]
- Elwood, R.W.; Adams, L. Electric shock causes physiological stress responses in shore crabs, consistent with prediction of pain. Biol. Lett. 2015, 11, 20150800. [Google Scholar] [CrossRef] [PubMed]
- Appel, M.; Elwood, R.W. Gender differences, responsiveness and memory of a potentially painful event in hermit crabs. Anim. Behav. 2009, 78, 1373–1379. [Google Scholar] [CrossRef]
- Britsih Veterinary Association. BVA Policy Position on the Recognition of Animals as Sentient Beings. Available online: https://www.bva.co.uk/media/4052/bva-policy-position-on-the-recognition-of-animals-as-sentient-beings.pdf (accessed on 9 May 2024).
- Animal Welfare (Sentiece) Act 2022. Available online: https://www.legislation.gov.uk/ukpga/2022/22/enacted (accessed on 3 March 2024).
- Comstock, G. Pain in Pleocyemata, but not in Dendrobranchiata? Anim. Sentience 2022, 7, 13. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barr, S.; Elwood, R.W. Effects of Acetic Acid and Morphine in Shore Crabs, Carcinus maenas: Implications for the Possibility of Pain in Decapods. Animals 2024, 14, 1705. https://doi.org/10.3390/ani14111705
Barr S, Elwood RW. Effects of Acetic Acid and Morphine in Shore Crabs, Carcinus maenas: Implications for the Possibility of Pain in Decapods. Animals. 2024; 14(11):1705. https://doi.org/10.3390/ani14111705
Chicago/Turabian StyleBarr, Stuart, and Robert W. Elwood. 2024. "Effects of Acetic Acid and Morphine in Shore Crabs, Carcinus maenas: Implications for the Possibility of Pain in Decapods" Animals 14, no. 11: 1705. https://doi.org/10.3390/ani14111705
APA StyleBarr, S., & Elwood, R. W. (2024). Effects of Acetic Acid and Morphine in Shore Crabs, Carcinus maenas: Implications for the Possibility of Pain in Decapods. Animals, 14(11), 1705. https://doi.org/10.3390/ani14111705