
Effect of Adding Yeast Cultures to High-Grain Conditions on Production Performance, Rumen Fermentation Profile, Microbial Abundance, and Immunity in Goats


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental, Animals, Design, and Diet
2.2. Product Performance
2.3. Sample Collection of Blood and Ruminal Fluid
2.4. Laboratory Analyses
2.5. DNA Extraction and Amplification
2.6. Bioinformatics and Statistical Analysis
2.7. Correlation between Rumen Microbiota and Blood Parameters Variables
2.8. Statistical Analysis
3. Results
3.1. Product Performance
3.2. The Digestibility of Nutrients
3.3. Blood Parameters
3.4. Rumen Fermentation Characteristics
3.5. Statistics of 16SrRNA Sequencing Results of Rumen Microorganisms
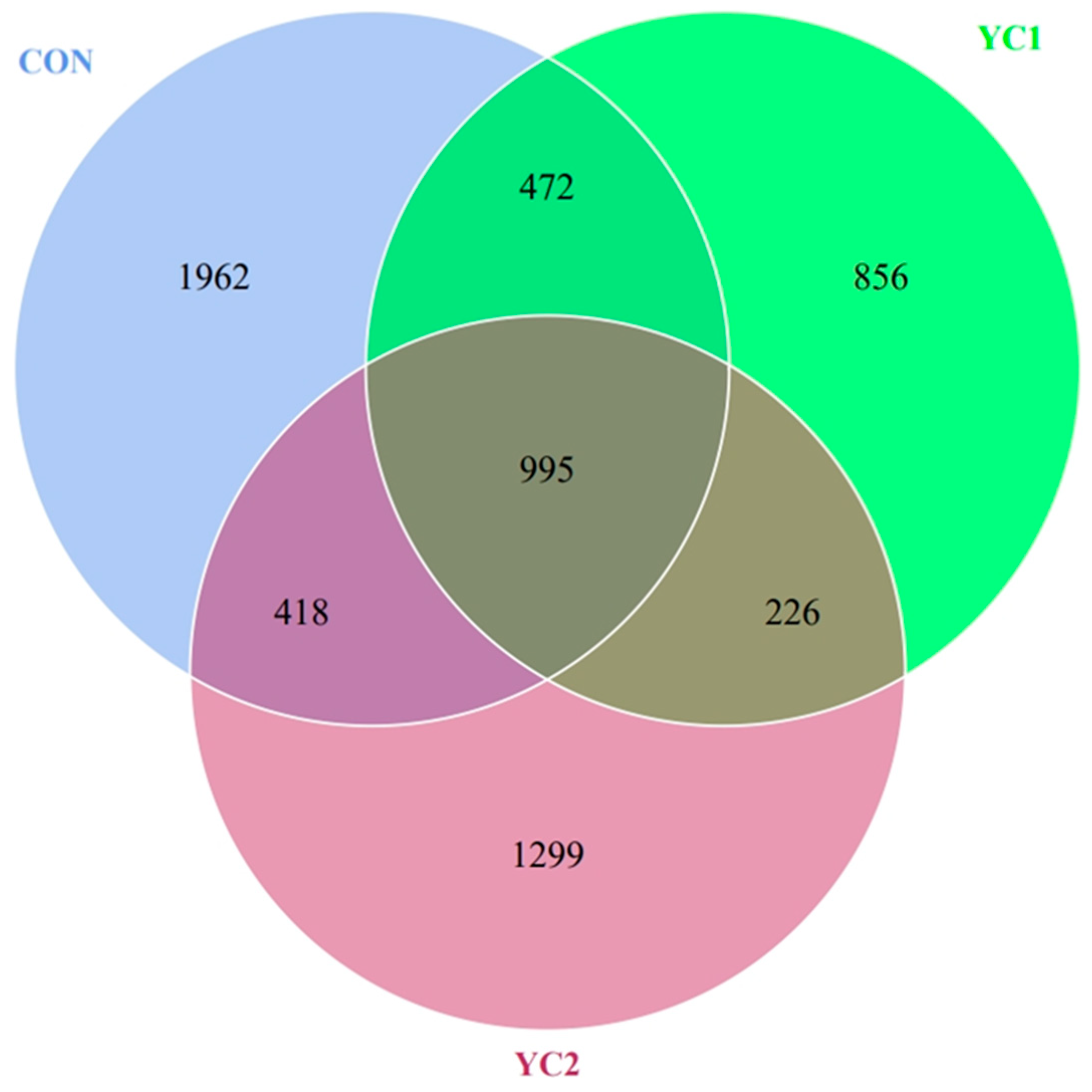
3.5.1. OTU Analysis
3.5.2. Alpha-Diversity Analysis
3.5.3. β Diversity Analysis
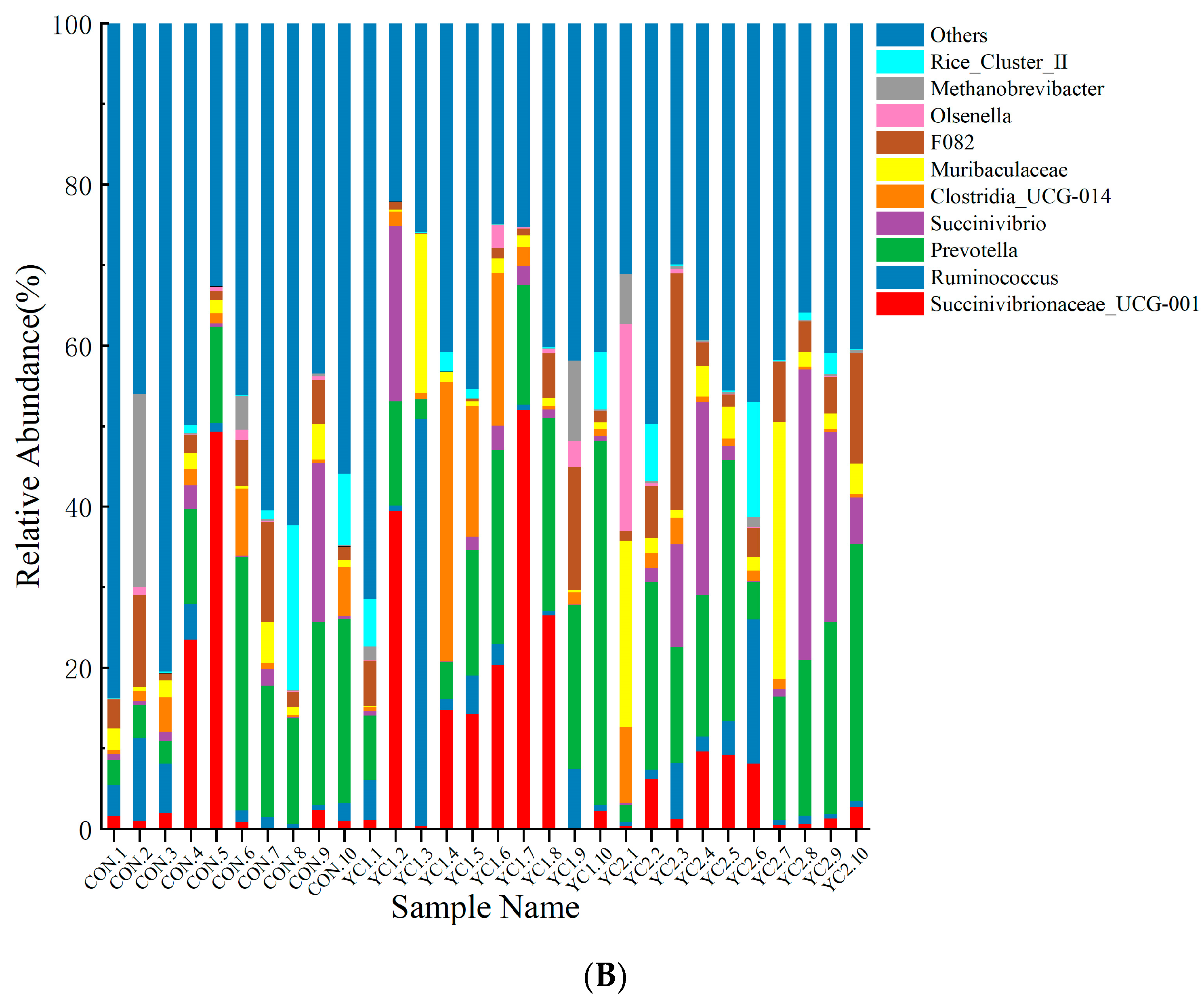
3.5.4. Microbiota Composition in Rumen
3.6. Correlation between Rumen Microbiota and Blood Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cavallini, D.; Mammi, L.M.E.; Palmonari, A.; García-González, R.; Chapman, J.D.; McLean, D.J.; Formigoni, A. Effect of an Immunomodulatory Feed Additive in Mitigating the Stress Responses in Lactating Dairy Cows to a High Concentrate Diet Challenge. Animals 2022, 12, 2129. [Google Scholar] [CrossRef] [PubMed]
- Khafipour, E.; Krause, D.O.; Plaizier, J.C. A grain-based subacute ruminal acidosis challenge causes translocation of lipopolysaccharide and triggers inflammation. J. Dairy Sci. 2009, 92, 1060–1070. [Google Scholar] [CrossRef] [PubMed]
- Tian, J. Effects of High Concentrate Diet on Rumen Fermentation and Epithelial Inflammation Related Gene Expression in Lactating Goats. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2018. [Google Scholar]
- Lu, Z.; Shen, H.; Shen, Z. High-Concentrate Diet-Induced Change of Cellular Metabolism Leads to Decreases of Immunity and Imbalance of Cellular Activities in Rumen Epithelium. Cell Physiol. Biochem. 2018, 45, 2145–2157. [Google Scholar] [CrossRef] [PubMed]
- Boyd, R.D.; Bauman, D.E.; Fox, D.G.; Scanes, C.G. Impact of metabolism modifiers on protein accretion and protein and energy requirements of livestock. J. Anim. Sci. 1991, 69, 56–75. [Google Scholar] [CrossRef]
- Reuben, R.C.; Elghandour, M.M.M.Y.; Alqaisi, O.; Cone, J.W.; Márquez, O.; Salem, A.Z.M. Influence of microbial probiotics on ruminant health and nutrition: Sources, mode of action and implications. J. Sci. Food Agric. 2022, 102, 1319–1340. [Google Scholar] [CrossRef] [PubMed]
- Wen, K.; Zhao, M.M.; Liu, L.; Khogali, M.K.; Geng, T.Y.; Wang, H.R.; Gong, D.Q. Thiamine modulates intestinal morphological structure and microbiota under subacute ruminal acidosis induced by a high-concentrate diet in Saanen goats. Animal 2021, 15, 100370. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Shah, A.M.; Wang, Z.; Peng, Q.; Hu, R.; Zou, H.; Tan, C.; Zhang, X.; Liao, Y.; Wang, Y.; et al. Effects of Land Transport Stress on Variations in Ruminal Microbe Diversity and Immune Functions in Different Breeds of Cattle. Animals 2019, 9, 599. [Google Scholar] [CrossRef] [PubMed]
- Li, L.P.; Peng, K.L.; Xue, M.Y.; Zhu, S.L.; Liu, J.X.; Sun, H.Z. An Age Effect of Rumen Microbiome in Dairy Buffaloes Revealed by Metagenomics. Microorganisms 2022, 10, 1491. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Liu, H.; Hu, L.; Xu, S.; Ai, D.; Fu, H.; Chen, Y.; Zhao, J. Characteristics and sex differences of rumen microflora of yaks around the lake. Acta Anim. Nutr. 2020, 32, 234–243. [Google Scholar]
- Wu, D.; Vinitchaikul, P.; Deng, M.; Zhang, G.; Sun, L.; Wang, H.; Gou, X.; Mao, H.; Yang, S. Exploration of the effects of altitude change on bacteria and fungi in the rumen of yak (Bos grunniens). Arch. Microbiol. 2021, 203, 835–846. [Google Scholar] [CrossRef]
- Fliegerova, K.O.; Podmirseg, S.M.; Vinzelj, J.; Grilli, D.J.; Kvasnová, S.; Schierová, D.; Sechovcová, H.; Mrázek, J.; Siddi, G.; Arenas, G.N.; et al. The Effect of a High-Grain Diet on the Rumen Microbiome of Goats with a Special Focus on Anaerobic Fungi. Microorganisms 2021, 9, 157. [Google Scholar] [CrossRef] [PubMed]
- Granja-Salcedo, Y.T.; Ribeiro Junior, C.S.; de Jesus, R.B.; Gomez-Insuasti, A.S.; Rivera, A.R.; Messana, J.D.; Canesin, R.C.; Berchielli, T.T. Effect of different levels of concentrate on ruminal microorganisms and rumen fermentation in Nellore steers. Arch. Anim. Nutr. 2016, 70, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.; Zhang, H.; Wen, H.; Wan, H.; Wu, H.; Chen, Y.; Li, S.; Zhang, L.; Sun, X.; Li, B.; et al. Yeast Probiotic and Yeast Products in Enhancing Livestock Feeds Utilization and Performance: An Overview. J. Fungi 2022, 8, 1191. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Perricone, V.; Sandrini, S.; Irshad, N.; Savoini, G.; Comi, M.; Agazzi, A. Yeast-Derived Products: The Role of Hydrolyzed Yeast and Yeast Culture in Poultry Nutrition-A Review. Animals 2022, 12, 1426. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Marden, J.P.; Julien, C.; Monteils, V.; Auclair, E.; Moncoulon, R.; Bayourthe, C. How does live yeast differ from sodium bicarbonate to stabilize ruminal pH in high-yielding dairy cows? J. Dairy Sci. 2008, 91, 3528–3535. [Google Scholar] [CrossRef] [PubMed]
- Newbold, C.J.; Wallace, R.J.; Chen, X.B.; McIntosh, F.M. Different strains of Saccharomyces cerevisiae differ in their effects on ruminal bacterial numbers in vitro and in sheep. J. Anim. Sci. 1995, 73, 1811–1818. [Google Scholar] [CrossRef] [PubMed]
- Lesmeister, K.E.; Heinrichs, A.J.; Gabler, M.T. Effects of supplemental yeast (Saccharomyces cerevisiae) culture on rumen development, growth characteristics, and blood parameters in neonatal dairy calves. J. Dairy Sci. 2004, 87, 1832–1839. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Wu, T.; You, P.; Wang, H.; Burke, J.L.; Kang, K.; Yu, W.; Wang, M.; Li, B.; He, Y.; et al. Dietary Supplementation of Yeast Culture Into Pelleted Total Mixed Rations Improves the Growth Performance of Fattening Lambs. Front. Vet. Sci. 2021, 8, 657816. [Google Scholar] [CrossRef] [PubMed]
- Garcia Diaz, T.; Ferriani Branco, A.; Jacovaci, F.A.; Cabreira Jobim, C.; Bolson, D.C.; Pratti Daniel, J.L. Inclusion of live yeast and mannan-oligosaccharides in high grain-based diets for sheep: Ruminal parameters, inflammatory response and rumen morphology. PLoS ONE 2018, 13, e0193313. [Google Scholar] [CrossRef]
- Emu, Q.; Guan, H.; Zhu, J.; Zhang, L.; Fan, J.; Ji, Y.; Lin, Y.; Li, C.; Dan, X.; Aguo, Y.; et al. Grazing and Supplementation of Dietary Yeast Probiotics Shape the Gut Microbiota and Improve the Immunity of Black Fattening Goats (Capra hircus). Front. Microbiol. 2021, 12, 666837. [Google Scholar] [CrossRef]
- Nocek, J.E.; Holt, M.G.; Oppy, J. Effects of supplementation with yeast culture and enzymatically hydrolyzed yeast on performance of early lactation dairy cattle. J. Dairy Sci. 2011, 94, 4046–4056. [Google Scholar] [CrossRef] [PubMed]
- NY/T816-2004; Feed Standard of Meat-Producing Sheep and Goats. Industry Standards-Agriculture: Beijing, China, 2004.
- Wang, Y.; Li, Q.; Wang, L.; Liu, Y.; Yan, T. Effects of a High-Concentrate Diet on the Blood Parameters and Liver Transcriptome of Goats. Animals 2023, 13, 1559. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis of the Association of Official’s Analytical Chemists; Association of Official Analytical Chemists: Arlington, VA, USA, 2003. [Google Scholar]
- Van Soest, P.V.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Adeola, O. Digestion and balance techniques in pigs. In Swine Nutrition, 2nd ed.; Lewis, A.J., Sothern, L.L., Eds.; CRC Press: Boca Raton, FL, USA, 2000; 906p. [Google Scholar]
- Jang, K.B.; Kim, S.W. Evaluation of standardized ileal digestibility of amino acids in fermented soybean meal for nursery pigs using direct and difference procedures. Anim. Biosci. 2023, 36, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Li, Y.; Wang, L.; Wang, J.; Xu, Q.; Yan, T.; Xue, B. Evaluation of composition and individual variability of rumen microbiota in yaks by 16S rRNA high-throughput sequencing technology. Anaerobe 2015, 34, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4516–4522. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Huse, S.M.; Welch, D.M.; Morrison, H.G.; Sogin, M.L. Ironing out the wrinkles in the rare biosphere through improved OTU clustering. Environ. Microbiol. 2010, 12, 1889–1898. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M.; et al. The Ribosomal Database Project: Improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 2009, 37, D141–D145. [Google Scholar] [CrossRef] [PubMed]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An effective distance metric for microbial community comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Desnoyers, M.; Giger-Reverdin, S.; Bertin, G.; Duvaux-Ponter, C.; Sauvant, D. Meta-analysis of the influence of Saccharomyces cerevisiae supplementation on ruminal parameters and milk production of ruminants. J. Dairy Sci. 2009, 92, 1620–1632. [Google Scholar] [CrossRef] [PubMed]
- Poppy, G.D.; Rabiee, A.R.; Lean, I.J.; Sanchez, W.K.; Dorton, K.L.; Morley, P.S. A meta-analysis of the effects of feeding yeast culture produced by anaerobic fermentation of Saccharomyces cerevisiae on milk production of lactating dairy cows. J. Dairy Sci. 2012, 95, 6027–6041. [Google Scholar] [CrossRef] [PubMed]
- Ogbuewu, I.P.; Mbajiorgu, C.A. Meta-analysis of Saccharomyces cerevisiae on enhancement of growth performance, rumen fermentation and haemato-biochemical characteristics of growing goats. Heliyon 2023, 9, e14178. [Google Scholar] [CrossRef]
- Dai, H.; Ma, N.; Chang, G.; Aabdin, Z.U.; Shen, X. Long-term high-concentrate diet feeding induces apoptosis of rumen epithelial cells and inflammation of rumen epithelium in dairy cows. Anim. Biotechnol. 2022, 33, 289–296. [Google Scholar] [CrossRef]
- Ma, N.; Abaker, J.A.; Wei, G.; Chen, H.; Shen, X.; Chang, G. A high-concentrate diet induces an inflammatory response and oxidative stress and depresses milk fat synthesis in the mammary gland of dairy cows. J. Dairy Sci. 2022, 105, 5493–5505. [Google Scholar] [CrossRef]
- Jiang, Y.; Dai, P.; Dai, Q.; Ma, J.; Wang, Z.; Hu, R.; Zou, H.; Peng, Q.; Wang, L.; Xue, B. Effects of the higher concentrate ratio on the production performance, ruminal fermentation, and morphological structure in male cattle-yaks. Vet. Med. Sci. 2022, 8, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Ogata, T.; Makino, H.; Ishizuka, N.; Iwamoto, E.; Masaki, T.; Ikuta, K.; Kim, Y.H.; Sato, S. Long-term high-grain diet altered the ruminal pH, fermentation, and composition and functions of the rumen bacterial community, leading to enhanced lactic acid production in Japanese Black beef cattle during fattening. PLoS ONE 2019, 14, e0225448. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Kong, L.; Ren, S.; Aschenbach, J.R.; Shen, H. Acid tolerance of lactate-utilizing bacteria of the order Bacteroidales contributes to prevention of ruminal acidosis in goats adapted to a high-concentrate diet. Anim. Nutr. 2023, 14, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Halfen, J.; Carpinelli, N.; Del Pino, F.A.B.; Chapman, J.D.; Sharman, E.D.; Anderson, J.L.; Osorio, J.S. Effects of yeast culture supplementation on lactation performance and rumen fermentation profile and microbial abundance in mid-lactation Holstein dairy cows. J. Dairy Sci. 2021, 104, 11580–11592. [Google Scholar] [CrossRef] [PubMed]
- Carpinelli, N.A.; Halfen, J.; Trevisi, E.; Chapman, J.D.; Sharman, E.D.; Anderson, J.L.; Osorio, J.S. Effects of peripartal yeast culture supplementation on lactation performance, blood biomarkers, rumen fermentation, and rumen bacteria species in dairy cows. J. Dairy Sci. 2021, 104, 10727–10743. [Google Scholar] [CrossRef] [PubMed]
- Sivinski, S.E.; Meier, K.E.; Mamedova, L.K.; Saylor, B.A.; Shaffer, J.E.; Sauls-Hiesterman, J.A.; Yoon, I.; Bradford, B.J. Effect of Saccharomyces cerevisiae fermentation product on oxidative status, inflammation, and immune response in transition dairy cattle. J. Dairy Sci. 2022, 105, 8850–8865. [Google Scholar] [CrossRef] [PubMed]
- Dias, A.L.G.; Freitas, J.A.; Micai, B.; Azevedo, R.A.; Greco, L.F.; Santos, J.E.P. Effect of supplemental yeast culture and dietary starch content on rumen fermentation and digestion in dairy cows. J. Dairy Sci. 2018, 101, 201–221. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Xu, S.; Liu, H.; Xu, T.; Hu, L.; Zhao, N.; Han, X.; Zhang, X. Yak rumen microbial diversity at different forage growth stages of an alpine meadow on the Qinghai-Tibet Plateau. PeerJ 2019, 7, e7645. [Google Scholar] [CrossRef] [PubMed]
- Petersen, C.; Round, J.L. Defining dysbiosis and its influence on host immunity and disease. Cell Microbiol. 2014, 16, 1024–1033. [Google Scholar] [CrossRef]
- Mei, S.; He, G.; Chen, Z.; Zhang, R.; Liao, Y.; Zhu, M.; Xu, D.; Shen, Y.; Zhou, B.; Wang, K.; et al. Probiotic-Fermented Distillers Grain Alters the Rumen Microbiome, Metabolome, and Enzyme Activity, Enhancing the Immune Status of Finishing Cattle. Animals 2023, 13, 3774. [Google Scholar] [CrossRef]
- Wang, Y.; Nan, X.; Zhao, Y.; Jiang, L.; Wang, H.; Zhang, F.; Hua, D.; Liu, J.; Yao, J.; Yang, L.; et al. Dietary Supplementation of Inulin Ameliorates Subclinical Mastitis via Regulation of Rumen Microbial Community and Metabolites in Dairy Cows. Microbiol. Spectr. 2021, 9, e0010521. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Sherlock, D.N.; Elolimy, A.A.; Vailati-Riboni, M.; Yoon, I.; Loor, J.J. Impact of a Saccharomyces cerevisiae fermentation product during an intestinal barrier challenge in lactating Holstein cows on ileal microbiota and markers of tissue structure and immunity. J. Anim. Sci. 2023, 101, skad309. [Google Scholar] [CrossRef] [PubMed]
- Li, D. Effects of Compound Bacteria Culture and Yeast β-Glucan on Antioxidant Function and Inflammatory Factors in Mutton Sheep; Inner Mongolia Agricultural University: Hohhot, China, 2021. [Google Scholar]
- Kowalik, B.; Skomiał, J.; Pająk, J.J.; Taciak, M.; Majewska, M.; Bełżecki, G. Population of ciliates, rumen fermentation indicators and biochemical parameters of blood serum in heifers fed diets supplemented with yeast (Saccharomyces cerevisiae) preparation. Anim. Sci. Pap. Rep. 2012, 30, 329–338. [Google Scholar]
- Zhang, J. Study on the Immune Enhancement Effect of Yeast Cell Wall Extract on Newcastle Disease Vaccine. Master’s Thesis, Southwest University, Chongqing, China, 2022. [Google Scholar] [CrossRef]
- Oshima, T.; Koseki, J.; Chen, X.; Matsumoto, T.; Miwa, H. Acid modulates the squamous epithelial barrier function by modulating the localization of claudins in the superficial layers. Lab. Invest. 2012, 92, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Su, M.; Wang, C.; Li, D.; Li, Q.; Liu, Z.; Qi, X.; Wu, Y.; Zhao, Y.; Li, T.; et al. Yeast culture repairs rumen epithelial injury by regulating microbial communities and metabolites in sheep. Front. Microbiol. 2023, 14, 1305772. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Xue, F.; Nan, X.; Tang, Z.; Wang, K.; Beckers, Y.; Jiang, L.; Xiong, B. Illumina Sequencing Approach to Characterize Thiamine Metabolism Related Bacteria and the Impacts of Thiamine Supplementation on Ruminal Microbiota in Dairy Cows Fed High-Grain Diets. Front. Microbiol. 2017, 8, 1818. [Google Scholar] [CrossRef]
- Bampidis, V.A.; Lymberopoulos, A.G.; Christodoulou, V.; Belibasaki, S. Impacts of supplemental dietary biotin on lameness in sheep. Anim. Feed. Sci. Technol. 2006, 134, 162–169. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Group | ||
|---|---|---|---|
| CON | YC1 | YC2 | |
| Ingredients | |||
| Corn | 50.70 | 50.20 | 50.20 |
| Soybean meal | 10.00 | 10.00 | 10.00 |
| DDGS | 6.00 | 6.00 | 6.00 |
| Alfalfa hay | 20.00 | 20.00 | 20.00 |
| Corn husk | 10.00 | 10.00 | 10.00 |
| CaCO3 | 0.80 | 0.80 | 0.80 |
| NaCl | 0.40 | 0.40 | 0.40 |
| Premix | 0.55 | 0.55 | 0.55 |
| NaHCO3 | 1.00 | 1.00 | 1.00 |
| MgO | 0.50 | 0.50 | 0.50 |
| Antioxidant | 0.05 | 0.05 | 0.05 |
| Yeast culture | 0.00 | 0.50 | 0.50 |
| Chemical composition | |||
| Dry matter | 89.05 | 89.05 | 89.05 |
| Crude protein | 14.20 | 14.20 | 14.21 |
| Crude ash | 6.44 | 6.44 | 6.44 |
| Crude fat | 4.31 | 4.31 | 4.31 |
| Metabolizable energy, M Cal/kg | 2.57 | 2.57 | 2.57 |
| Neutral detergent fiber | 48.05 | 48.05 | 48.05 |
| Acid detergent fiber | 12.12 | 12.12 | 12.12 |
| Calcium | 0.56 | 0.56 | 0.56 |
| Phosphorus | 0.38 | 0.38 | 0.38 |
| Concentrate:roughage | 80:20 | 80:20 | 80:20 |
| Group | p-Value | |||
|---|---|---|---|---|
| CON | YC1 | YC2 | ||
| Initial body weight, kg | 25.75 ± 0.42 | 25.42 ± 0.40 | 25.72 ± 0.96 | 0.828 |
| BW at 30 days, kg | 30.99 ± 0.23 | 31.95 ± 0.39 | 32.20 ± 0.86 | 0.297 |
| ADG, g/d | 174.67 ± 6.82 b | 213.33 ± 4.77 a | 216.67 ± 9.33 a | <0.001 |
| ADFI, kg/d | 1.11 ± 0.02 b | 1.15 ± 0.05 a | 1.17 ± 0.06 a | 0.009 |
| F/G | 6.35 ± 0.26 a | 5.39 ± 0.21 b | 5.41 ± 0.16 b | 0.005 |
| Item | Group | p-Value | ||
|---|---|---|---|---|
| CON | YC1 | YC2 | ||
| Dry matter | 68.07 ± 1.20 b | 73.45 ± 1.47 a | 72.54 ± 1.55 a | 0.002 |
| Crude protein | 70.34 ± 2.02 b | 72.78 ± 1.22 ab | 74.22 ± 1.60 a | 0.009 |
| Acid detergent fiber | 44.79 ± 1.91 b | 52.20 ± 0.84 a | 52.43 ± 1.63 a | 0.012 |
| Neutral detergent fiber | 69.47 ± 1.76 b | 74.78 ± 1.16 a | 73.48 ± 0.60 a | 0.016 |
| Ether extract | 71.43 ± 2.27 b | 78.38 ± 1.42 a | 72.58 ± 2.26 b | 0.009 |
| Items | Group | p-Value | ||
|---|---|---|---|---|
| CON | YC1 | YC2 | ||
| Glutamic pyruvic transaminase, ALT, U/L | 21.99 ± 1.63 | 19.93 ± 1.26 | 20.85 ± 1.63 | 0.634 |
| Glutamic oxaloacetic transaminase, AST, U/L | 66.47 ± 6.26 | 66.99 ± 6.24 | 68.43 ± 5.44 | 0.972 |
| Urea, mmol/L | 5.50 ± 0.40 b | 6.11 ± 0.45 b | 7.61 ± 0.29 a | 0.005 |
| Creatinine, CREA, umol/L | 55.34 ± 5.34 | 60.64 ± 5.65 | 65.98 ± 3.10 | 0.314 |
| Alkaline phosphatase, ALP, U/L | 307.3 ± 87.41 | 512.10 ± 145.27 | 348.2 ± 107.02 | 0.428 |
| Lactate dehydrogenase, LDH, U/L | 278.05 ± 18.89 | 291.41 ± 22.47 | 296.88 ± 25.35 | 0.830 |
| Creatine kinase, CK, U/L | 109.89 ± 11.22 | 110.07 ± 14.77 | 110.67 ± 11.24 | 0.998 |
| Total protein, TP, g/L | 55.34 ± 3.72 | 54.13 ± 2.65 | 59.81 ± 2.63 | 0.393 |
| Albumin, ALB, g/L | 27.43 ± 1.72 | 26.12 ± 1.21 | 30.35 ± 1.52 | 0.143 |
| Total cholesterol, TC, mmol/L | 1.54 ± 0.12 | 1.45 ± 0.12 | 1.68 ± 0.14 | 0.466 |
| Triglyceride, TG, mmol/L | 0.25 ± 0.03 b | 0..43 ± 0.07 a | 0.28 ± 0.04 b | 0.044 |
| Low-density lipoprotein cholesterol, LDL-C, mmol/L | 0.54 ± 0.05 | 0.57 ± 0.05 | 0.57 ± 0.05 | 0.904 |
| High-density lipoprotein cholesterol, HDL-C, mmol/L | 1.51 ± 0.10 ab | 1.33 ± 0.11 b | 1.72 ± 0.13 a | 0.065 |
| Lipopolysaccharide, LPS, ng/L | 249.37 ± 5.89 | 241.71 ± 7.59 | 263.88 ± 11.30 | 0.197 |
| Histamine, HIS, ng/mL | 9.59 ± 0.26 | 9.94 ± 0.21 | 10.26 ± 0.38 | 0.286 |
| Interleukin-1β, IL-1β, pg/mL | 78.60 ± 10.46 a | 36.05 ± 4.17 b | 34.07 ± 15.96 b | <0.001 |
| Interleukin-6, IL-6, pg/mL | 92.84 ± 8.95 a | 39.99 ± 4.53 b | 37.69 ± 3.94 b | <0.001 |
| Interleukin-10, IL-10, pg/mL | 20.53 ± 1.94 b | 41.66 ± 1.48 a | 25.70 ± 2.87 b | <0.001 |
| Tumor Necrosis Factor-α, TNF-α, pg/mL | 157.76 ± 12.31 a | 91.84 ± 5.81 b | 97.19 ± 5.21 b | <0.001 |
| Immunoglobulin M, IgM, μg/mL | 1733.44 ± 73.71 b | 1926.57 ± 87.40 ab | 2052.36 ± 80.37 a | 0.031 |
| Immunoglobulin A, IgA, μg/mL | 152.45 ± 21.18 b | 206.60 ± 8.11 a | 199.92 ± 11.17 a | 0.028 |
| Immunoglobulin G, IgG, mg/mL | 5.24 ± 0.43 b | 7.01 ± 0.23 a | 7.52 ± 0.26 a | <0.001 |
| Major acute phase protein, MAP, mg/L | 112.33 ± 9.79 | 107.59 ± 5.50 | 97.78 ± 6.79 | 0.395 |
| Heat shock protein 70, HP-70, pg/mL | 375.41 ± 32.46 | 317.08 ± 24.80 | 337.57 ± 22.65 | 0.315 |
| Item | Group | p-Value | ||
|---|---|---|---|---|
| CON | YC1 | YC2 | ||
| pH | 7.05 ± 0.08 | 7.08 ± 0.06 | 7.06 ± 0.07 | 0.655 |
| TVFA, mmol/l | 40.36 ± 5.17 | 39.63 ± 5.49 | 36.86 ± 3.41 | 0.871 |
| Acetic acid, mmol/L | 25.70 ± 3.54 | 25.18 ± 23.04 | 23.04 ± 2.36 | 0.820 |
| Propionic acid, mmol/L | 9.80 ± 1.88 | 11.25 ± 2.14 | 10.01 ± 1.08 | 0.824 |
| Butyric acid, mmol/L | 2.88 ± 0.54 | 1.60 ± 0.26 | 2.03 ± 0.50 | 0.136 |
| Isobutyric acid, mmol/L | 0.66 ± 0.10 | 0.55 ± 0.08 | 0.60 ± 0.07 | 0.715 |
| Isovaleric acid, mmol/L | 0.89 ± 0.11 | 0.72 ± 0.12 | 0.81 ± 0.06 | 0.542 |
| Pentanoic acid, mmol/L | 0.49 ± 0.07 | 0.36 ± 0.08 | 0.38 ± 0.04 | 0.306 |
| Acetic acid/Propionic acid | 2.94 ± 0.27 | 2.83 ± 0.60 | 2.34 ± 0.20 | 0.565 |
| Microbial protein, μg/L | 16.64 ± 2.24 b | 28.13 ± 1.90 a | 28.60 ± 5.37 a | 0.039 |
| NH3-N, mg/dl | 70.90 ± 13.10 | 67.09 ± 6.92 | 60.23 ± 1.66 | 0.656 |
| Item | Group | p-Value | ||
|---|---|---|---|---|
| CON | YC1 | YC2 | ||
| Chao1 | 802.11 ± 57.60 a | 522.66 ± 44.54 b | 719.69 ± 47.48 a | 0.002 |
| Dominance | 0.04 ± 0.00 b | 0.10 ± 0.02 a | 0.06 ± 0.01 ab | 0.051 |
| goods_coverage | 0.99 ± 0.00 | 0.99 ± 0.00 | 0.99 ± 0.00 | 0.087 |
| observed_otus | 761.6 ± 52.94 a | 500.40 ± 43.23 b | 677.11 ± 43.42 a | 0.002 |
| pielou_e | 0.71 ± 0.03 a | 0.61 ± 0.03 b | 0.68 ± 0.03 ab | 0.071 |
| Shannon | 6.81 ± 0.31 a | 5.49 ± 0.34 b | 6.39 ± 0.31 ab | 0.020 |
| Simpson | 0.96 ± 0.01 a | 0.90 ± 0.02 b | 0.94 ± 0.01 ab | 0.051 |
| Item | Group | p-Value | ||
|---|---|---|---|---|
| CON | YC1 | YC2 | ||
| Phylum | ||||
| Bacteroidota | 30.27 ± 3.74 b | 31.38 ± 4.64 b | 52.03 ± 4.25 a | 0.002 |
| Firmicutes | 36.44 ± 6.44 a | 33.78 ± 6.94 a | 15.11 ± 1.97 b | 0.033 |
| Fibrobacterota | 0.24 ± 0.06 b | 0.10 ± 0.03 b | 0.91 ± 0.21 a | <0.001 |
| genus | ||||
| Rikenellaceae_RC9_gut_group | 3.13 ± 0.69 a | 0.71 ± 0.23 b | 4.17 ± 0.1.18 a | 0.012 |
| Prevotellaceae_UCG-001 | 0.96 ± 0.20 b | 0.81 ± 0.19 b | 2.19 ± 0.45 a | 0.006 |
| Prevotellaceae_UCG-003 | 0.41 ± 0.20 b | 0.30 ± 0.07 b | 1.26 ± 0.28 a | 0.004 |
| Fibrobacter | 0.24 ± 0.06 b | 0.10 ± 0.03 b | 0.91 ± 0.21 a | <0.001 |
| Prevotellaceae_UCG-010 | 0.45 ± 0.11 a | 0.13 ± 0.05 b | 0.22 ± 0.04 b | 0.018 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, P.; Wang, L. Effect of Adding Yeast Cultures to High-Grain Conditions on Production Performance, Rumen Fermentation Profile, Microbial Abundance, and Immunity in Goats. Animals 2024, 14, 1799. https://doi.org/10.3390/ani14121799
Qi P, Wang L. Effect of Adding Yeast Cultures to High-Grain Conditions on Production Performance, Rumen Fermentation Profile, Microbial Abundance, and Immunity in Goats. Animals. 2024; 14(12):1799. https://doi.org/10.3390/ani14121799
Chicago/Turabian StyleQi, Pei, and Lizhi Wang. 2024. "Effect of Adding Yeast Cultures to High-Grain Conditions on Production Performance, Rumen Fermentation Profile, Microbial Abundance, and Immunity in Goats" Animals 14, no. 12: 1799. https://doi.org/10.3390/ani14121799