
Behavioral Adaptations in Tropical Dairy Cows: Insights into Calving Day Predictions
 ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal, Housing, and Calving Management
2.2. Definition of Parturition
2.3. Data Preparation
Processing Sensor Data
2.4. Statistical Analysis
2.5. Development of Calving Prediction Model
2.6. Programming Packages
3. Results
3.1. Effect of Lactation Number on Behavior around Calving
3.2. Periparturient Activity Changes across Lactation Groups in Dairy Cows


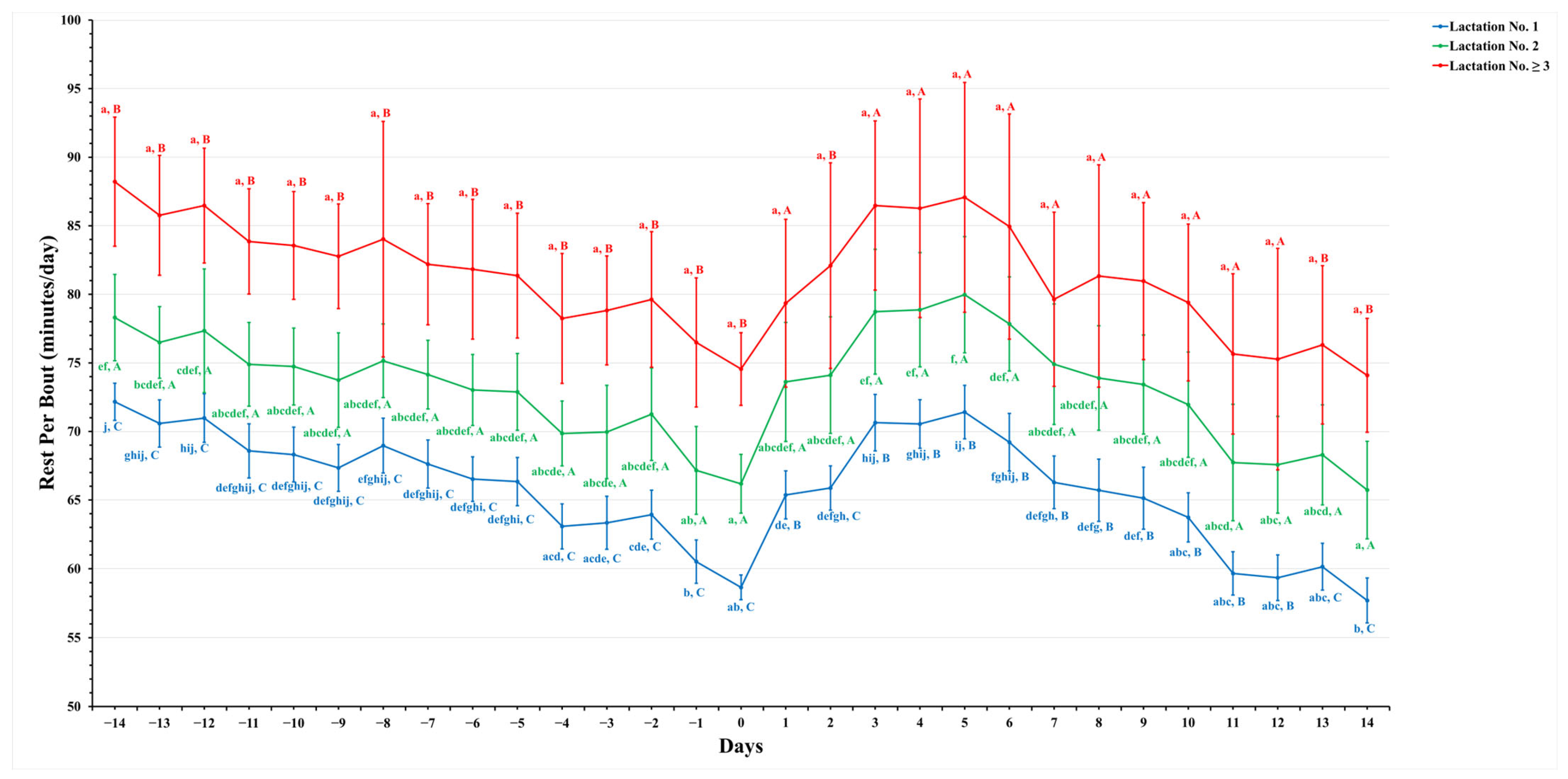

3.3. Lactation Groups and Temporal Dynamics Influence Activity Patterns

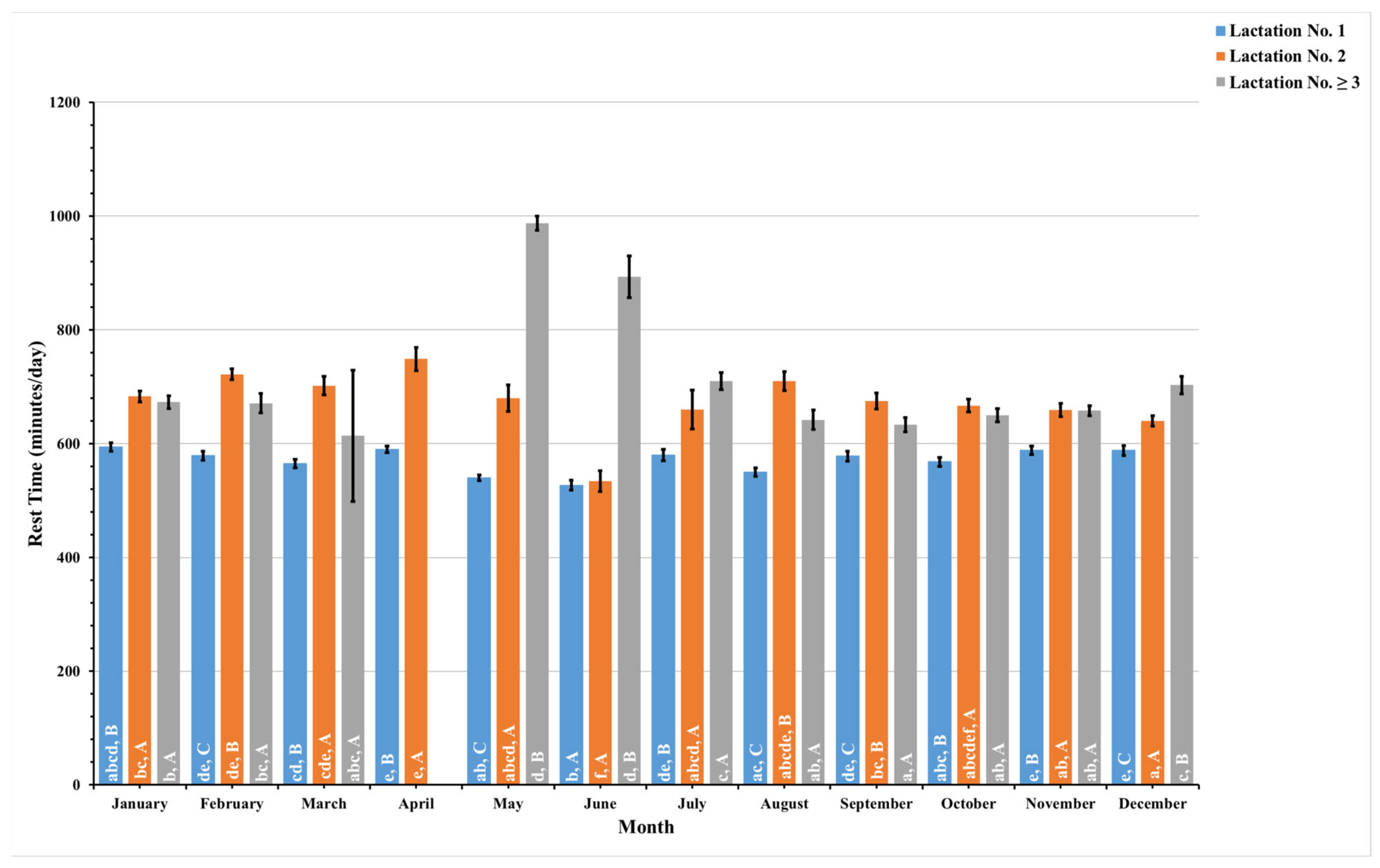


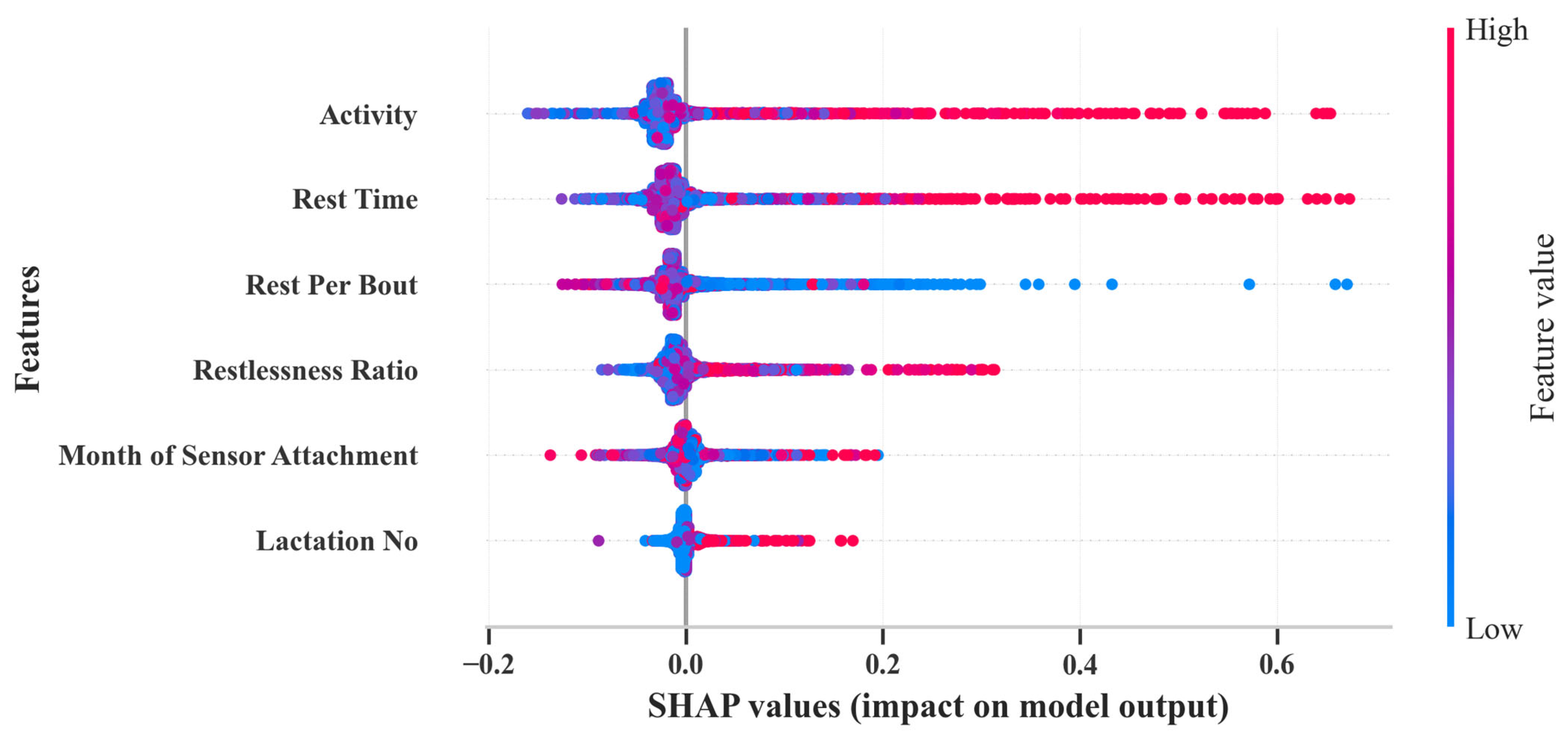
3.4. Machine Learning Model Evaluation
4. Discussion
4.1. Effect of Lactation Number on Behavior around Calving
4.2. Periparturient Activity Changes across Lactation Groups in Dairy Cows
4.3. Lactation Groups and Temporal Dynamics Influence Activity Patterns
4.4. Calving Prediction
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Independent Variables | N (9318) | Activity (Minutes/Day) | Rest Time (Minutes/Day) | Rest per Bout (Minutes/Day) | Restlessness Ratio | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 95% CI | 95% CI | 95% CI | 95% CI | ||||||||||
| Mean ± SE | Lower | Upper | Mean ± SE | Lower | Upper | Mean ± SE | Lower | Upper | Mean ± SE | Lower | Upper | ||
| Lact. 1 | 6314 (68.1%) | 218.7 ± 2.8 | 213.2 | 224.3 | 583.4 ± 5.3 | 573.0 | 593.9 | 66.4 ± 1.0 | 64.5 | 68.3 | 3.9 ± 0.1 | 3.7 | 4.0 |
| Lact. 2 | 1831 (19.7%) | 209.3 ± 4.9 | 199.7 | 218.9 | 659.5 ± 9.4 | 641.1 | 678.0 | 70.1 ± 1.7 | 66.7 | 73.4 | 3.0 ± 0.2 | 2.7 | 3.3 |
| Lact. ≥ 3 | 1146 (12.3%) | 206.6 ± 6.6 | 193.7 | 219.5 | 655.2 ± 12.5 | 630.7 | 679.7 | 75.5 ± 2.3 | 71.0 | 79.9 | 3.0 ± 0.2 | 2.6 | 3.4 |
| January | 969 (10.4%) | 226.3 ± 4.8 | 217.0 | 235.7 | 641.8 ± 9.3 | 623.6 | 660.0 | 75.1 ± 1.7 | 71.9 | 78.4 | 3.5 ± 0.2 | 3.2 | 3.8 |
| February | 937 (10.1%) | 203.2 ± 4.8 | 193.9 | 212.6 | 653.7 ± 9.3 | 635.6 | 672.1 | 69.7 ± 1.7 | 66.4 | 73.0 | 2.85 ± 0.2 | 2.5 | 3.2 |
| March | 862 (9.3%) | 217.4 ± 5.2 | 207.1 | 227.6 | 626.1 ± 10.2 | 606.1 | 646.1 | 66.2 ± 1.8 | 62.7 | 69.8 | 3.4 ± 0.2 | 3.0 | 3.7 |
| April | 924 (9.9%) | 216.6 ± 5.5 | 205.9 | 227.3 | 638.3 ± 10.7 | 617.4 | 659.2 | 69.0 ± 1.9 | 65.3 | 72.8 | 3.2 ± 0.2 | 2.9 | 3.6 |
| May | 943 (10.1%) | 226.5 ± 5.7 | 215.4 | 237.6 | 613.7 ± 11.0 | 592.1 | 635.2 | 65.1 ± 2.0 | 61.3 | 69.0 | 3.6 ± 0.2 | 3.2 | 4.0 |
| June | 444 (4.8%) | 223.26 ± 6.7 | 210.1 | 236.4 | 619.7 ± 13.2 | 593.8 | 645.5 | 56.9 ± 2.3 | 52.3 | 61.5 | 3.8 ± 0.2 | 3.3 | 4.3 |
| July | 304 (3.3%) | 181.1 ± 7.8 | 165.8 | 196.5 | 614.7 ± 15.5 | 584.3 | 645.1 | 61.6 ± 2.7 | 56.3 | 67.0 | 2.8 ± 0.3 | 2.2 | 3.4 |
| August | 476 (5.1%) | 193.3 ± 6.4 | 180.7 | 205.8 | 625.8 ± 12.6 | 601.1 | 65.4 | 66.5 ± 2.2 | 62.1 | 70.8 | 3.09 ± 0.2 | 2.6 | 3.6 |
| September | 663 (7.1%) | 204.4 ± 5.6 | 193.4 | 215.5 | 623.8 ± 11.0 | 602.3 | 645.4 | 78.4 ± 2.0 | 74.5 | 82.2 | 3.4 ± 0.2 | 3.0 | 3.8 |
| October | 808 (8.7%) | 203.58 ± 4.9 | 193.9 | 213.3 | 610.8 ± 9.7 | 591.9 | 629.8 | 84.3 ± 1.7 | 80.9 | 87.7 | 3.3 ± 0.2 | 2.9 | 3.6 |
| November | 1116 (12.0%) | 209.8 ± 4.5 | 200.9 | 218.6 | 652.1 ± 8.8 | 634.9 | 669.4 | 74.5 ± 1.6 | 71.4 | 77.6 | 3.1 ± 0.2 | 2.8 | 3.4 |
| December | 872 (9.4%) | 232.9 ± 4.8 | 223.5 | 242.3 | 671.97 ± 9.4 | 653.5 | 690.4 | 80.3 ± 1.7 | 77.0 | 83.6 | 3.37 ± 0.2 | 3.0 | 3.7 |
| Day −14 | 287 (3.1%) | 209.5 ± 5.0 | 199.8 | 219.2 | 744.4 ± 9.9 | 724.9 | 763.9 | 76.4 ± 1.7 | 73.0 | 79.8 | 2.1 ± 0.2 | 1.7 | 2.5 |
| Day −13 | 292 (3.1%) | 180.5 ± 4.9 | 170.8 | 190.1 | 708.45 ± 9.9 | 689.1 | 727.8 | 74.5 ± 1.7 | 71.1 | 77.9 | 2.0 ± 0.2 | 1.6 | 2.4 |
| Day −12 | 295 (3.2%) | 179.6 ± 4.9 | 169.9 | 189.2 | 701.1 ± 9.9 | 681.8 | 720.4 | 75.2 ± 1.7 | 71.8 | 78.6 | 2.1 ± 0.2 | 1.7 | 2.5 |
| Day −11 | 295 (3.2%) | 180.7 ± 4.9 | 171.1 | 190.4 | 696.2 ± 9.8 | 676.9 | 715.5 | 73.0 ± 1.7 | 69.6 | 76.4 | 2.1 ± 0.2 | 1.7 | 2.5 |
| Day −10 | 295 (3.2%) | 181.7 ± 4.9 | 172.0 | 191.3 | 701.7 ± 9.8 | 682.4 | 720.9 | 72.9 ± 1.7 | 69.5 | 76.2 | 2.0 ± 0.2 | 1.6 | 2.4 |
| Day −9 | 295 (3.2%) | 176.8 ± 4.9 | 167.2 | 186.5 | 703.9 ± 9.8 | 684.6 | 723.2 | 71.9 ± 1.7 | 68.5 | 75.3 | 2.0 ± 0.2 | 1.6 | 2.4 |
| Day −8 | 295 (3.2%) | 178.2 ± 4.9 | 168.6 | 187.9 | 703.0 ± 9.8 | 683.7 | 722.3 | 73.4 ± 1.7 | 70.0 | 76.8 | 2.0 ± 0.2 | 1.6 | 2.4 |
| Day −7 | 295 (3.2%) | 175.7 ± 4.9 | 166.1 | 185.3 | 699.9 ± 9.8 | 680.6 | 719.1 | 72.1 ± 1.7 | 68.7 | 75.4 | 2.0 ± 0.2 | 1.6 | 2.4 |
| Day −6 | 295 (3.2%) | 183.6 ± 4.9 | 174.0 | 193.2 | 687.4 ± 9.8 | 668.2 | 706.7 | 71.1 ± 1.7 | 67.7 | 74.5 | 2.2 ± 0.2 | 1.8 | 2.6 |
| Day −5 | 295 (3.2%) | 181.1 ± 4.9 | 171.5 | 190.7 | 698.9 ± 9.8 | 679.7 | 718.2 | 70.9 ± 1.7 | 67.5 | 74.3 | 2.2 ± 0.2 | 1.8 | 2.6 |
| Day −4 | 295 (3.2%) | 186.6 ± 4.9 | 177.0 | 196.2 | 676.64 ± 9.8 | 657.4 | 695.9 | 67.8 ± 1.7 | 64.2 | 71.1 | 2.4 ± 0.2 | 2.0 | 2.8 |
| Day −3 | 294 (3.2%) | 187.2 ± 4.9 | 177.6 | 196.8 | 668.1 ± 9.8 | 648.8 | 687.3 | 68.0 ± 1.7 | 64.6 | 71.4 | 2.5 ± 0.2 | 2.1 | 2.9 |
| Day −2 | 294 (3.2%) | 192.8 ± 4.9 | 183.2 | 202.4 | 666.5 ± 9.8 | 647.3 | 685.7 | 68.9 ± 1.7 | 65.5 | 72.3 | 2.6 ± 0.2 | 2.2 | 3.0 |
| Day −1 | 292 (3.1%) | 210.3 ± 4.9 | 200.7 | 220.0 | 683.8 ± 9.8 | 664.6 | 703.1 | 65.5 ± 1.7 | 62.1 | 68.9 | 2.9 ± 0.2 | 2.5 | 3.3 |
| Day 0 | 363 (3.9%) | 296.2 ± 4.6 | 287.2 | 305.2 | 674.3 ± 9.2 | 656.3 | 692.2 | 63.4 ± 1.6 | 60.2 | 66.6 | 5.0 ± 0.2 | 4.6 | 5.4 |
| Day 1 | 341 (3.7%) | 331.6 ± 4.7 | 322.4 | 340.8 | 525.7 ± 9.4 | 507.4 | 544.1 | 70.4 ± 1.7 | 67.2 | 73.6 | 7.26 ± 0.2 | 6.9 | 7.6 |
| Day 2 | 344 (3.7%) | 315.4 ± 4.7 | 306.2 | 324.6 | 528.3 ± 9.3 | 510.0 | 546.6 | 71.0 ± 1.6 | 67.8 | 74.2 | 6.7 ± 0.2 | 6.3 | 7.1 |
| Day 3 | 346 (3.7%) | 281.9 ± 4.7 | 272.7 | 291.1 | 558.5 ± 9.3 | 540.3 | 576.8 | 75.6 ± 1.6 | 72.4 | 78.8 | 5.2 ± 0.2 | 4.8 | 5.6 |
| Day 4 | 346 (3.7%) | 251.3 ± 4.7 | 242.1 | 260.4 | 579.8 ± 9.3 | 561.5 | 598.1 | 75.6 ± 1.6 | 72.3 | 78.6 | 4.2 ± 0.2 | 3.8 | 4.6 |
| Day 5 | 347 (3.7%) | 224.9 ± 4.7 | 215.7 | 234.0 | 583.4 ± 9.3 | 565.2 | 601.7 | 76.4 ± 1.6 | 73.2 | 79.7 | 3.7 ± 0.2 | 3.3 | 4.1 |
| Day 6 | 347 (3.7%) | 209.5 ± 4.7 | 200.3 | 218.7 | 568.5 ± 9.3 | 550.2 | 586.8 | 74.3 ± 1.6 | 71.1 | 77.5 | 3.6 ± 0.2 | 3.3 | 4.0 |
| Day 7 | 345 (3.7%) | 209.3 ± 4.7 | 200.1 | 218.5 | 564.6 ± 9.3 | 546.3 | 582.9 | 71.6 ± 1.6 | 68.4 | 74.8 | 3.7 ± 0.2 | 3.3 | 4.1 |
| Day 8 | 347 (3.7%) | 208.9 ± 4.7 | 199.7 | 218.1 | 568.5 ± 9.3 | 550.2 | 586.7 | 70.8 ± 1.6 | 67.4 | 74.0 | 3.6 ± 0.2 | 3.3 | 4.0 |
| Day 9 | 346 (3.7%) | 213.3 ± 4.7 | 204.1 | 222.4 | 565.3 ± 9.3 | 547.0 | 583.6 | 70.4 ± 1.6 | 67.2 | 73.6 | 3.68 ± 0.2 | 3.3 | 4.1 |
| Day 10 | 347 (3.7%) | 206.9 ± 4.7 | 197.7 | 216.1 | 574.5 ± 9.3 | 556.2 | 592.8 | 68.9 ± 1.6 | 65.7 | 72.1 | 3.4 ± 0.2 | 3.0 | 3.8 |
| Day 11 | 346 (3.7%) | 205.9 ± 4.7 | 196.8 | 215.1 | 575.0 ± 9.3 | 556.7 | 593.3 | 65.0 ± 1.6 | 61.8 | 68.2 | 3.7 ± 0.2 | 3.3 | 4.0 |
| Day 12 | 346 (3.7%) | 194.6 ± 4.7 | 185.5 | 203.8 | 580.1 ± 9.3 | 561.8 | 598.3 | 64.8 ± 1.6 | 61.6 | 68.0 | 3.8 ± 0.2 | 3.4 | 4.1 |
| Day 13 | 346 (3.7%) | 195.3 ± 4.7 | 186.2 | 204.5 | 582.8 ± 9.3 | 564.5 | 601.1 | 65.7 ± 1.6 | 62.3 | 68.9 | 3.2 ± 0.2 | 2.9 | 3.6 |
| Day 14 | 347 (3.7%) | 185.2 ± 4.7 | 176.1 | 194.4 | 579.8 ± 9.3 | 561.5 | 598.1 | 63.2 ± 1.6 | 60.0 | 66.4 | 3.1 ± 0.2 | 2.8 | 3.5 |
References
- Mao, A.; Huang, E.; Wang, X.; Liu, K. Deep learning-based animal activity recognition with wearable sensors: Overview, challenges, and future directions. Comput. Electron. Agric. 2023, 211, 108043. [Google Scholar] [CrossRef]
- Eerdekens, A.; Deruyck, M.; Fontaine, J.; Martens, L.; De Poorter, E.; Plets, D.; Joseph, W. A framework for energy-efficient equine activity recognition with leg accelerometers. Comput. Electron. Agric. 2021, 183, 106020. [Google Scholar] [CrossRef]
- Ouellet, V.; Vasseur, E.; Heuwieser, W.; Burfeind, O.; Maldague, X.; Charbonneau, É. Evaluation of calving indicators measured by automated monitoring devices to predict the onset of calving in Holstein dairy cows. J. Dairy Sci. 2016, 99, 1539–1548. [Google Scholar] [CrossRef] [PubMed]
- Heinrichs, A.; Heinrichs, B.; Cavallini, D.; Fustini, M.; Formigoni, A. Limiting total mixed ration availability alters eating and rumination patterns of lactating dairy cows. JDS Commun. 2021, 2, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Von Keyserlingk, M.; Rushen, J.; de Passillé, A.M.; Weary, D.M. Invited review: The welfare of dairy cattle—Key concepts and the role of science. J. Dairy Sci. 2009, 92, 4101–4111. [Google Scholar] [CrossRef] [PubMed]
- Matamala, F.; Strappini, A.; Sepúlveda-Varas, P. Dairy cow behaviour around calving: Its relationship with management practices and environmental conditions. Austral J. Vet. Sci. 2021, 53, 9–22. [Google Scholar] [CrossRef]
- Leliveld, L.M.; Riva, E.; Mattachini, G.; Finzi, A.; Lovarelli, D.; Provolo, G. Dairy cow behavior is affected by period, time of day and housing. Animals 2022, 12, 512. [Google Scholar] [CrossRef] [PubMed]
- Giannone, C.; Bovo, M.; Ceccarelli, M.; Torreggiani, D.; Tassinari, P. Review of the Heat Stress-Induced Responses in Dairy Cattle. Animals 2023, 13, 3451. [Google Scholar] [CrossRef]
- Tao, S.; Rivas, R.M.O.; Marins, T.N.; Chen, Y.-C.; Gao, J.; Bernard, J.K. Impact of heat stress on lactational performance of dairy cows. Theriogenology 2020, 150, 437–444. [Google Scholar] [CrossRef]
- Hut, P.; Scheurwater, J.; Nielen, M.; van den Broek, J.; Hostens, M. Heat stress in a temperate climate leads to adapted sensor-based behavioral patterns of dairy cows. J. Dairy Sci. 2022, 105, 6909–6922. [Google Scholar] [CrossRef]
- Huzzey, J.; Von Keyserlingk, M.; Weary, D. Changes in feeding, drinking, and standing behavior of dairy cows during the transition period. J. Dairy Sci. 2005, 88, 2454–2461. [Google Scholar] [CrossRef]
- Proudfoot, K.L.; Huzzey, J.M. A first time for everything: The influence of parity on the behavior of transition dairy cows. JDS Commun. 2022, 3, 467–471. [Google Scholar] [CrossRef]
- Schirmann, K.; Weary, D.; Heuwieser, W.; Chapinal, N.; Cerri, R.; Von Keyserlingk, M. Rumination and feeding behaviors differ between healthy and sick dairy cows during the transition period. J. Dairy Sci. 2016, 99, 9917–9924. [Google Scholar] [CrossRef]
- Von Keyserlingk, M.A.; Weary, D. Feeding behaviour of dairy cattle: Meaures and applications. Can. J. Anim. Sci. 2010, 90, 303–309. [Google Scholar] [CrossRef]
- Wankhade, P.R.; Manimaran, A.; Kumaresan, A.; Jeyakumar, S.; Ramesha, K.; Sejian, V.; Rajendran, D.; Varghese, M.R. Metabolic and immunological changes in transition dairy cows: A review. Vet. World 2017, 10, 1367. [Google Scholar] [CrossRef]
- Borchers, M.; Chang, Y.; Proudfoot, K.; Wadsworth, B.; Stone, A.; Bewley, J. Machine-learning-based calving prediction from activity, lying, and ruminating behaviors in dairy cattle. J. Dairy Sci. 2017, 100, 5664–5674. [Google Scholar] [CrossRef]
- Iqbal, M.W.; Draganova, I.; Morel, P.C.; Morris, S.T. Validation of an Accelerometer Sensor-Based Collar for Monitoring Grazing and Rumination Behaviours in Grazing Dairy Cows. Animals 2021, 11, 2724. [Google Scholar] [CrossRef]
- Araújo, S.O.; Peres, R.S.; Ramalho, J.C.; Lidon, F.; Barata, J. Machine Learning Applications in Agriculture: Current Trends, Challenges, and Future Perspectives. Agronomy 2023, 13, 2976. [Google Scholar] [CrossRef]
- Zehner, N.; Niederhauser, J.J.; Schick, M.; Umstatter, C. Development and validation of a predictive model for calving time based on sensor measurements of ingestive behavior in dairy cows. Comput. Electron. Agric. 2019, 161, 62–71. [Google Scholar] [CrossRef]
- Clark, C.; Lyons, N.; Millapan, L.; Talukder, S.; Cronin, G.; Kerrisk, K.; Garcia, S. Rumination and activity levels as predictors of calving for dairy cows. Animal 2015, 9, 691–695. [Google Scholar] [CrossRef]
- Keceli, A.S.; Catal, C.; Kaya, A.; Tekinerdogan, B. Development of a recurrent neural networks-based calving prediction model using activity and behavioral data. Comput. Electron. Agric. 2020, 170, 105285. [Google Scholar] [CrossRef]
- Giaretta, E.; Marliani, G.; Postiglione, G.; Magazzù, G.; Pantò, F.; Mari, G.; Formigoni, A.; Accorsi, P.; Mordenti, A. Calving time identified by the automatic detection of tail movements and rumination time, and observation of cow behavioural changes. Animal 2021, 15, 100071. [Google Scholar] [CrossRef]
- Shahinfar, S.; Page, D.; Guenther, J.; Cabrera, V.; Fricke, P.; Weigel, K. Prediction of insemination outcomes in Holstein dairy cattle using alternative machine learning algorithms. J. Dairy Sci. 2014, 97, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Boonkum, W.; Misztal, I.; Duangjinda, M.; Pattarajinda, V.; Tumwasorn, S.; Sanpote, J. Genetic effects of heat stress on milk yield of Thai Holstein crossbreds. J. Dairy Sci. 2011, 94, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Mee, J.F. Managing the dairy cow at calving time. Vet. Clin. Food Anim. Pract. 2004, 20, 521–546. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.B. Behaviour around the time of calving in dairy cows. Appl. Anim. Behav. Sci. 2012, 139, 195–202. [Google Scholar] [CrossRef]
- Miedema, H.M.; Cockram, M.S.; Dwyer, C.M.; Macrae, A.I. Changes in the behaviour of dairy cows during the 24 h before normal calving compared with behaviour during late pregnancy. Appl. Anim. Behav. Sci. 2011, 131, 8–14. [Google Scholar] [CrossRef]
- Schuenemann, G.; Nieto, I.; Bas, S.; Galvão, K.; Workman, J. Assessment of calving progress and reference times for obstetric intervention during dystocia in Holstein dairy cows. J. Dairy Sci. 2011, 94, 5494–5501. [Google Scholar] [CrossRef]
- Henriksen, J.; Munksgaard, L. Validation of AfiTagII, a device for automatic measuring of lying behaviour in Holstein and Jersey cows on two different bedding materials. Animal 2019, 13, 617–621. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Shahinfar, S.; Khansefid, M.; Haile-Mariam, M.; Pryce, J. Machine learning approaches for the prediction of lameness in dairy cows. Animal 2021, 15, 100391. [Google Scholar] [CrossRef] [PubMed]
- Warner, D.; Vasseur, E.; Lefebvre, D.M.; Lacroix, R. A machine learning based decision aid for lameness in dairy herds using farm-based records. Comput. Electron. Agric. 2020, 169, 105193. [Google Scholar] [CrossRef]
- Bonestroo, J.; van der Voort, M.; Hogeveen, H.; Emanuelson, U.; Klaas, I.C.; Fall, N. Forecasting chronic mastitis using automatic milking system sensor data and gradient-boosting classifiers. Comput. Electron. Agric. 2022, 198, 107002. [Google Scholar] [CrossRef]
- Friedman, J.H. Greedy function approximation: A gradient boosting machine. Ann. Stat. 2001, 29, 1189–1232. [Google Scholar] [CrossRef]
- Becker, C.; Aghalari, A.; Marufuzzaman, M.; Stone, A. Predicting dairy cattle heat stress using machine learning techniques. J. Dairy Sci. 2021, 104, 501–524. [Google Scholar] [CrossRef]
- Harris, C.R.; Millman, K.J.; Van Der Walt, S.J.; Gommers, R.; Virtanen, P.; Cournapeau, D.; Wieser, E.; Taylor, J.; Berg, S.; Smith, N.J. Array programming with NumPy. Nature 2020, 585, 357–362. [Google Scholar] [CrossRef]
- Reback, J.; McKinney, W.; Van Den Bossche, J.; Augspurger, T.; Cloud, P.; Klein, A.; Hawkins, S.; Roeschke, M.; Tratner, J.; She, C. pandas-dev/pandas: Pandas 1.0.5; Zenodo: Geneva, Switzerland, 2020. [Google Scholar]
- Microsoft Corporation. Microsoft Excel (Student and Home Edition) [Software]; Microsoft: Redmond, WA, USA, 2021. [Google Scholar]
- Waskom, M.L. Seaborn: Statistical data visualization. J. Open Source Softw. 2021, 6, 3021. [Google Scholar] [CrossRef]
- Hunter, J.D. Matplotlib: A 2D graphics environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- Santos, M.R.; Guedes, A.; Sanchez-Gendriz, I. SHapley Additive exPlanations (SHAP) for Efficient Feature Selection in Rolling Bearing Fault Diagnosis. Mach. Learn. Knowl. Extr. 2024, 6, 316–341. [Google Scholar] [CrossRef]
- Zhu, X.; Zhang, M.; Wen, Y.; Shang, D. Machine learning advances the integration of covariates in population pharmacokinetic models: Valproic acid as an example. Front. Pharmacol. 2022, 13, 994665. [Google Scholar] [CrossRef]
- Blackie, N.; Scaife, J.; Bleach, E. Lying behaviour and activity of early lactation Holstein dairy cattle measured using an activity monitor. Cattle Pract. 2006, 14, 139. [Google Scholar]
- Hwang, W.Y.; Choi, S.Y.; An, H.J. Concept analysis of transition to motherhood: A methodological study. Korean J. Women Health Nurs. 2022, 28, 8. [Google Scholar] [CrossRef] [PubMed]
- Titler, M.; Maquivar, M.; Bas, S.; Rajala-Schultz, P.J.; Gordon, E.; McCullough, K.; Federico, P.; Schuenemann, G.M. Prediction of parturition in Holstein dairy cattle using electronic data loggers. J. Dairy Sci. 2015, 98, 5304–5312. [Google Scholar] [CrossRef]
- Miedema, H.M.; Cockram, M.S.; Dwyer, C.M.; Macrae, A.I. Behavioural predictors of the start of normal and dystocic calving in dairy cows and heifers. Appl. Anim. Behav. Sci. 2011, 132, 14–19. [Google Scholar] [CrossRef]
- Creutzinger, K.; Dann, H.; Krawczel, P.D.; Habing, G.; Proudfoot, K. The effect of stocking density and a blind on the behavior of Holstein dairy cattle in group maternity pens. Part I: Calving location, locomotion, and separation behavior. J. Dairy Sci. 2021, 104, 7109–7121. [Google Scholar] [CrossRef]
- Proudfoot, K.; Veira, D.; Weary, D.; Von Keyserlingk, M. Competition at the feed bunk changes the feeding, standing, and social behavior of transition dairy cows. J. Dairy Sci. 2009, 92, 3116–3123. [Google Scholar] [CrossRef]
- Steensels, M.; Bahr, C.; Berckmans, D.; Halachmi, I.; Antler, A.; Maltz, E. Lying patterns of high producing healthy dairy cows after calving in commercial herds as affected by age, environmental conditions and production. Appl. Anim. Behav. Sci. 2012, 136, 88–95. [Google Scholar] [CrossRef]
- Neave, H.; Lomb, J.; Von Keyserlingk, M.; Behnam-Shabahang, A.; Weary, D. Parity differences in the behavior of transition dairy cows. J. Dairy Sci. 2017, 100, 548–561. [Google Scholar] [CrossRef] [PubMed]
- Tucker, C.B.; Jensen, M.B.; de Passillé, A.M.; Hänninen, L.; Rushen, J. Invited review: Lying time and the welfare of dairy cows. J. Dairy Sci. 2021, 104, 20–46. [Google Scholar] [CrossRef]
- Jensen, M.B.; Proudfoot, K.L. Effect of group size and health status on behavior and feed intake of multiparous dairy cows in early lactation. J. Dairy Sci. 2017, 100, 9759–9768. [Google Scholar] [CrossRef]
- Von Keyserlingk, M.; Olenick, D.; Weary, D. Acute behavioral effects of regrouping dairy cows. J. Dairy Sci. 2008, 91, 1011–1016. [Google Scholar] [CrossRef] [PubMed]
- Rice, C.A.; Eberhart, N.L.; Krawczel, P.D. Prepartum lying behavior of Holstein dairy cows housed on pasture through parturition. Animals 2017, 7, 32. [Google Scholar] [CrossRef] [PubMed]
- Antanaitis, R.; Anskienė, L.; Palubinskas, G.; Džermeikaitė, K.; Bačėninaitė, D.; Viora, L.; Rutkauskas, A. Ruminating, Eating, and Locomotion Behavior Registered by Innovative Technologies around Calving in Dairy Cows. Animals 2023, 13, 1257. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, N.E.; Wicks, H.C.; Carson, A.F.; McCoy, M.A. Influence of post-calving regrouping strategy on welfare and performance parameters in dairy heifers. Appl. Anim. Behav. Sci. 2008, 114, 319–329. [Google Scholar] [CrossRef]
- Gargiulo, J.I.; Eastwood, C.R.; Garcia, S.C.; Lyons, N.A. Dairy farmers with larger herd sizes adopt more precision dairy technologies. J. Dairy Sci. 2018, 101, 5466–5473. [Google Scholar] [CrossRef]
- Tucker, C.; Dalley, D.; Burke, J.-L.; Clark, D. Milking cows once daily influences behavior and udder firmness at peak and mid lactation. J. Dairy Sci. 2007, 90, 1692–1703. [Google Scholar] [CrossRef]
- Ito, K.; Chapinal, N.; Weary, D.; Von Keyserlingk, M. Associations between herd-level factors and lying behavior of freestall-housed dairy cows. J. Dairy Sci. 2014, 97, 2081–2089. [Google Scholar] [CrossRef]
- Westin, R.; Vaughan, A.; de Passillé, A.M.; Devries, T.J.; Pajor, E.A.; Pellerin, D.; Siegford, J.M.; Vasseur, E.; Rushen, J. Lying times of lactating cows on dairy farms with automatic milking systems and the relation to lameness, leg lesions, and body condition score. J. Dairy Sci. 2016, 99, 551–561. [Google Scholar] [CrossRef]
- Könyves, T.; Zlatković, N.; Memiši, N.; Lukač, D.; Puvača, N.; Stojšin, M.; Halász, A.; Miščević, B. Relationship of temperature-humidity index with milk production and feed intake of holstein-frisian cows in different year seasons. Thai J. Vet. Med. 2017, 47, 15–23. [Google Scholar] [CrossRef]
- Thammahakin, P.; Yawongsa, A.; Rukkwamsuk, T. Effect of heat stress on reproductive performance of dairy cows under tropical climate: A review. J. Kasetsart Vet. 2020, 30, 111–132. [Google Scholar]
- Perera, K.; Gwazdauskas, F.; Pearson, R.; Brumback, T., Jr. Effect of season and stage of lactation on performance of Holsteins. J. Dairy Sci. 1986, 69, 228–236. [Google Scholar] [CrossRef]
- Vázquez-Diosdado, J.A.; Gruhier, J.; Miguel-Pacheco, G.; Green, M.; Dottorini, T.; Kaler, J. Accurate prediction of calving in dairy cows by applying feature engineering and machine learning. Prev. Vet. Med. 2023, 219, 106007. [Google Scholar] [CrossRef] [PubMed]
- Rutten, C.; Kamphuis, C.; Hogeveen, H.; Huijps, K.; Nielen, M.; Steeneveld, W. Sensor data on cow activity, rumination, and ear temperature improve prediction of the start of calving in dairy cows. Comput. Electron. Agric. 2017, 132, 108–118. [Google Scholar] [CrossRef]
- Crociati, M.; Sylla, L.; De Vincenzi, A.; Stradaioli, G.; Monaci, M. How to predict parturition in cattle? A literature review of automatic devices and technologies for remote monitoring and calving prediction. Animals 2022, 12, 405. [Google Scholar] [CrossRef] [PubMed]
- Barkema, H.W.; von Keyserlingk, M.A.; Kastelic, J.P.; Lam, T.J.; Luby, C.; Roy, J.-P.; LeBlanc, S.J.; Keefe, G.P.; Kelton, D.F. Invited review: Changes in the dairy industry affecting dairy cattle health and welfare. J. Dairy Sci. 2015, 98, 7426–7445. [Google Scholar] [CrossRef] [PubMed]
- Martin-Collado, D.; Hely, F.; Byrne, T.; Evans, R.; Cromie, A.; Amer, P. Farmer views on calving difficulty consequences on dairy and beef farms. Animal 2017, 11, 318–326. [Google Scholar] [CrossRef]
- Fenlon, C.; O’Grady, L.; Mee, J.F.; Butler, S.T.; Doherty, M.L.; Dunnion, J. A comparison of 4 predictive models of calving assistance and difficulty in dairy heifers and cows. J. Dairy Sci. 2017, 100, 9746–9758. [Google Scholar] [CrossRef]

| Ingredients | Inclusion (kg/day) |
|---|---|
| Soya bean meal | 1.1 |
| Napier grass | 30 |
| Rice straw | 4 |
| Premix | 0.14 |
| Molasses | 0.2 |
| Selenium | 0.08 |
| Total | 35.52 |
| Proximate analysis (dry matter basis) | |
| Dry matter% | 36.7 |
| Crude protein% | 16.7 |
| Crude fat% | 2.0 |
| Crude fiber% | 27.5 |
| Ash% | 26.1 |
| Detergent analysis (dry matter basis) | |
| NEFL | 27.7 |
| ADF% | 25.03 |
| NDF% | 39.26 |
| ADL% | 3.29 |
| Cellulose% | 21.74 |
| Hemicellulose | 14.23 |
| Behavioral Metrics | Description |
|---|---|
| Activity | The cumulative sum of the movement or physical activity displayed by the dairy cow per day. |
| Rest time | The cumulative duration the dairy cow spends lying down and resting per day. |
| Rest per bout | A measure of the average duration of continuous period during which the dairy cow remains lying down. |
| Restlessness ratio | A measure of how much the dairy cow is moving or shifting while it is lying down. |
| Independent Variables | Activity (Minutes/Day) | Rest Time (Minutes/Day) | Rest per Bout (Minutes/Day) | Restlessness Ratio | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| β ± SE | p-Value | 95% CI | β ± SE | p-Value | 95% CI | β ± SE | p-Value | 95% CI | β ± SE | p-Value | 95% CI | |||||
| Lower | Upper | Lower | Upper | Lower | Upper | Lower | Upper | |||||||||
| Intercept | 2017 ± 8.2 | <0.0 | 185.7 | 217.7 | 641.5 ± 16.0 | <0.0 | 610.2 | 672.8 | 77.7 ± 2.8 | <0.0 | 72.1 | 83.3 | 2.9 ± 0.3 | <0.0 | 2.3 | 3.5 |
| Lact. 1 | 12.1± 7.2 | <0.0 | −2.0 | 26.2 | −71.7 ± 13.7 | <0.0 | −98.5 | −45.0 | −9.1 ± 2.5 | <0.0 | −14.0 | −4.2 | 0.9 ± 0.2 | <0.0 | 0.5 | 1.3 |
| Lact. 2 | 2.7 ± 7.7 | 0.7 | −124 | 17.7 | 4.4 ± 14.7 | 0.8 | −24.5 | 33.2 | −5.4 ± 2.7 | <0.0 | −10.6 | −0.2 | 0.1 ± 0.2 | 0.8 | −0.4 | 0.5 |
| Lact. ≥ 3 | 0 b | - | - | - | 0 b | - | - | - | 0 b | - | - | - | 0 b | - | - | - |
| January | −6.5 ± 5.0 | 0.2 | −16.4 | 3.4 | −30.2 ± 10.2 | <0.0 | −50.1 | −10.2 | −5.2 ± 1.8 | <0.0 | −8.7 | −1.7 | 0.1 ± 0.2 | 0.6 | −0.3 | 0.5 |
| February | −29.6 ± 5.7 | <0.0 | −40.8 | −18.5 | −18.1 ± 11.4 | 0.1 | −40.4 | 4.2 | −10.6 ± 2.0 | <0.0 | −14.5 | −6.7 | −0.5 ± 0.2 | <0.0 | −1.0 | −0.1 |
| March | −15.5 ± 6.3 | <0.0 | −27.9 | −3.1 | −45.8 ± 12.6 | <0.0 | −70.6 | −21.1 | −14.1 ± 2.2 | <0.0 | −18.4 | −9.7 | −0.0 ± 0.2 | 1.0 | −0.5 | 0.5 |
| April | −16.3 ± 6.6 | <0.0 | −29.3 | −3.2 | −33.6 ± 13.1 | <0.0 | −59.4 | −7.9 | −11.3 ± 2.3 | <0.0 | −15.8 | −6.7 | −0.1 ± 0.2 | 0.6 | −0.6 | 0.4 |
| May | −6.4 ± 6.8 | 0.4 | −19.8 | 7.0 | −58.3 ± 13.4 | <0.0 | −84.7 | −32.0 | −15.2 ± 2.4 | <0.0 | −19.9 | −10.5 | 0.2 ± 0.3 | 0.4 | −0.3 | 0.7 |
| June | −9.6 ± 7.8 | 0.2 | −24.8 | 5.6 | −52.3 ± 15.3 | <0.0 | −82.3 | −22.3 | −23.4 ± 2.7 | <0.0 | −28.7 | −18.1 | 0.6 ± 0.3 | 0.1 | −0.1 | 1.0 |
| July | −51.7 ± 8.8 | <0.0 | −69.0 | −34.5 | −57.2 ± 17.5 | <0.0 | −91.5 | −23.0 | −18.7 ± 3.1 | <0.0 | −24.7 | −12.6 | −0.6 ± 0.3 | 0.1 | −1.2 | 0.1 |
| August | −39.6 ± 7.5 | <0.0 | −54.3 | −24.9 | −46.2 ± 14.9 | <0.0 | −75.4 | −17.1 | −13.8 ± 2.6 | <0.0 | −19.0 | −8.7 | −0.3 ± 0.3 | 0.3 | −0.8 | 0.3 |
| September | −28.4 ± 6.8 | <0.0 | −41.9 | −15.2 | −48.1 ± 13.5 | <0.0 | −74.5 | −21.8 | −1.9 ± 2.4 | 0.4 | −6.6 | 2.7 | 0.0 ± 0.3 | 1.0 | −0.5 | 0.5 |
| October | −29.3 ± 5.7 | <0.0 | −40.5 | −18.1 | −61.1 ± 11.5 | <0.0 | −83.7 | −38.6 | 4.0 ± 2.1 | <0.0 | 0.1 | 8.0 | −0.1 ± 0.23 | 0.7 | −0.6 | 0.4 |
| November | −23.1 ± 4.5 | <0.0 | −32.0 | −14.2 | −19.8 ± 9.2 | <0.0 | −37.9 | −1.7 | −5.8 ± 1.6 | <0.0 | −8.9 | −2.6 | −0.3 ± 0.2 | 0.2 | −0.7 | 0.1 |
| December | 0 b | - | - | - | 0 b | - | - | - | 0 b | - | - | - | 0 b | - | - | - |
| Day −14 | 24.3 ± 5.5 | <0.0 | 13.8 | 34.7 | 164.6 ± 11.0 | <0.0 | 142.9 | 186.3 | 13.2 ± 1.9 | <0.0 | 9.5 | 16.9 | −1.0 ± 0.3 | <0.0 | −1.5 | −0.5 |
| Day −13 | −4.8 ± 5.3 | 0.4 | −15.2 | 5.7 | 128.7 ± 11.0 | <0.0 | 107.1 | 150.2 | 11.3 ± 1.9 | <0.0 | 7.6 | 15.0 | −1.1 ± 0.3 | <0.0 | −1.6 | −0.6 |
| Day −12 | −5.7 ± 5.3 | 0.3 | −16.1 | 4.7 | 121.3 ± 11.0 | <0.0 | 99.8 | 142.8 | 12.0 ± 1.9 | <0.0 | 8.4 | 15.7 | −1.0 ± 0.3 | <0.0 | −1.5 | −0.6 |
| Day −11 | −4.5 ± 5.3 | 0.4 | −14.9 | 5.9 | 116.4 ± 11.0 | <0.0 | 94.9 | 137.9 | 9.8 ± 1.9 | <0.0 | 6.1 | 13.5 | −1.0 ± 0.3 | <0.0 | −1.5 | −0.6 |
| Day −10 | −3.6 ± 5.3 | 0.5 | −14.0 | 6.8 | 121.9 ± 11.0. | <0.0 | 100.4 | 143.4 | 9.7 ± 1.9 | <0.0 | 6.0 | 13.4 | −1.1 ± 0.3 | <0.0 | −1.6 | −0.6 |
| Day −9 | −8.4 ± 5.3 | 0.1 | −18.8 | 2.0 | 124.1 ± 11.0 | <0.0 | 102.6 | 145.6 | 8.7 ± 1.9 | <0.0 | 5.0 | 12.4 | −1.1 ± 0.3 | <0.0 | −1.6 | −0.6 |
| Day −8 | −7.0 ± 5.3 | 0.2 | −17.4 | 3.4 | 123.2 ± 11.0 | <0.0 | 101.7 | 144.7 | 10.2 ± 1.9 | <0.0 | 6.5 | 13.9 | −1.1 ± 0.3 | <0.0 | −1.6 | −0.6 |
| Day −7 | −9.6 ± 5.3 | 0.1 | −19.9 | 0.8 | 120.1 ± 11.0 | <0.0 | 98.6 | 141.6 | 8.9 ± 1.9 | <0.0 | 5.2 | 12.6 | −1.1 ± 0.3 | <0.0 | −1.6 | −0.6 |
| Day −6 | −1.7 ± 5.3 | 0.8 | −12.0 | 8.7 | 107.6 ± 11.0 | <0.0 | 86.2 | 129.1 | 7.9 ± 1.9 | <0.0 | 4.2 | 11.6 | −0.9 ± 0.3 | <0.0 | −1.4 | −0.4 |
| Day −5 | −4.2 ± 5.3 | 0.4 | −14.5 | 6.2 | 119.2 ± 11.0 | <0.0 | 97.7 | 140.6 | 7.7 ± 1.9 | <0.0 | 4.0 | 11.4 | −0.9 ± 0.3 | <0.0 | −1.4 | −0.5 |
| Day −4 | 1.3 ± 5.3 | 0.8 | −9.0 | 11.7 | 96.8 ± 10.9 | <0.0 | 75.4 | 118.3 | 4.6 ± 1.9 | <0.0 | 0.9 | 8.3 | −0.8 ± 0.3 | <0.0 | −1.3 | −0.3 |
| Day −3 | 2.0 ± 5.3 | 0.7 | −8.4 | 12.3 | 88.3 ± 11.0 | <0.0 | 66.8 | 109.8 | 4.8 ± 1.9 | <0.0 | 1.2 | 8.5 | −0.6 ± 0.3 | <0.0 | −1.1 | −0.1 |
| Day −2 | 7.6 ± 5.3 | 0.2 | −2.8 | 17.9 | 86.7 ± 11.0 | <0.0 | 65.2 | 108.2 | 5.7 ± 1.9 | <0.0 | 2.0 | 9.4 | −0.5 ± 0.3 | <0.0 | −1.0 | −0.0 |
| Day −1 | 25.1 ± 5.3 | <0.0 | 14.7 | 35.5 | 104.1 ± 11.0 | <0.0 | 82.5 | 125.6 | 2.3 ± 1.9 | 0.2 | −1.4 | 6.0 | −0.2 ± 0.3 | 0.4 | −0.7 | 0.3 |
| Day 0 | 111.0 ± 5.0 | <0.0 | 101.2 | 120.8 | 94.5 ± 10.4 | <0.0 | 74.2 | 114.8 | 0.2 ± 1.8 | 0.9 | −3.3 | 3.7 | 1.8 ± 0.2 | <0.0 | 1.4 | 2.3 |
| Day 1 | 146.3 ± 5.1 | <0.0 | 136.4 | 156.3 | −54.1 ± 10.5 | <0.0 | −74.6 | −33.5 | 7.2 ± 1.8 | <0.0 | 3.7 | 10.7 | 4.1 ± 0.2 | <0.0 | 3.7 | 4.6 |
| Day 2 | 130.1 ± 5.1 | <0.0 | 120.2 | 140.0 | −51.5 ± 10.5 | <0.0 | −72.0 | −31.0 | 7.8 ± 1.8 | <0.0 | 4.3 | 11.3 | 3.5 ± 0.2 | <0.0 | 3.1 | 4.0 |
| Day 3 | 96.7 ± 5.1 | <0.0 | 86.8 | 106.6 | −21.2 ± 10.4 | <0.0 | −41.7 | −0.8 | 12.4 ± 1.8 | <0.0 | 8.9 | 15.9 | 2.0 ± 0.2 | <0.0 | 1.6 | 2.5 |
| Day 4 | 66.0 ± 5.0 | <0.0 | 56.1 | 75.9 | −0.0 ± 10.4 | 1.0 | −20.5 | 20.5 | 12.4 ± 1.8 | <0.0 | 8.8 | 15.9 | 1.1 ± 0.2 | <0.0 | 0.6 | 1.6 |
| Day 5 | 39.6 ± 5.0 | <0.0 | 29.7 | 49.5 | 3.7 ± 10.4 | 0.7 | −16.8 | 24.1 | 13.3 ± 1.8 | <0.0 | 9.7 | 16.8 | 0.6 ± 0.2 | <0.0 | 0.1 | 1.0 |
| Day 6 | 24.3 ± 5.0 | <0.0 | 14.4 | 34.2 | −11.3 ± 10.4 | 0.3 | −31.7 | 9.2 | 11.1 ± 1.8 | <0.0 | 7.6 | 14.6 | 0.5 ± 0.2 | <0.0 | 0.0 | 1.0 |
| Day 7 | 24.1 ± 5.0 | <0.0 | 14.2 | 34.0 | −15.1 ± 10.4 | 0.1 | −35.6 | 5.3 | 8.4 ± 1.8 | <0.0 | 4.9 | 12.0 | 0.5 ± 0.2 | <0.0 | 0.1 | 1.0 |
| Day 8 | 23.7 ± 5.0 | <0.0 | 13.8 | 33.5 | −11.3 ± 10.4 | 0.3 | −31.8 | 9.1 | 7.6 ± 1.8 | <0.0 | 4.1 | 11.1 | 0.5 ± 0.2 | <0.0 | 0.0 | 1.0 |
| Day 9 | 28.0 ± 5.0 | <0.0 | 18.1 | 37.9 | −14.5 ± 10.4 | 0.2 | −34.9 | 6.0 | 7.2 ± 1.8 | <0.0 | 3.7 | 10.8 | 0.6 ± 0.2 | <0.0 | 0.1 | 1.0 |
| Day 10 | 21.6 ± 5.0 | <0.0 | 11.8 | 31.5 | −5.3 ± 10.4 | 0.6 | −25.8 | 15.1 | 5.7 ± 1.8 | <0.0 | 2.2 | 9.2 | 0.3 ± 0.2 | 0.3 | −0.2 | 0.7 |
| Day 11 | 20.7 ± 5.0 | <0.0 | 10.8 | 30.6 | −4.7 ± 10.4 | 0.6 | −25.2 | 15.7 | 1.8 ± 1.8 | 0.3 | −1.7 | 5.3 | 0.5 ± 0.2 | <0.0 | 0.0 | 1.0 |
| Day 12 | 9.4 ± 5.0 | 0.1 | −0.5 | 19.3 | 0.3 ± 10.4 | 1.0 | −20.2 | 20.7 | 1.6 ± 1.8 | 0.4 | −1.9 | 5.2 | 0.6 ± 0.2 | <0.0 | 0.2 | 1.1 |
| Day 13 | 10.1 ± 5.0 | <0.0 | 0.2 | 20.0 | 3.0 ± 10.4 | 0.8 | −17.4 | 23.5 | 2.5 ± 1.8 | 0.2 | −1.0 | 6.0 | 0.1 ± 0.2 | 0.6 | −0.4 | 0.6 |
| Day 14 | 0 b | - | - | - | 0 b | - | - | - | 0 b | - | - | - | 0 b | - | - | - |
| Machine Learning Models | Sensitivity (%) | Specificity (%) | Positive Predictive Values (%) | Negative Predictive Value (%) | Accuracy Score (%) | F2 Score (%) |
|---|---|---|---|---|---|---|
| Random forest | 40.0 | 98.8 | 68.8 | 96.2 | 95.2 | 43.7 |
| Decision tree | 49.1 | 94.1 | 35.5 | 96.6 | 91.4 | 45.6 |
| Gradient boosting | 34.6 | 98.8 | 55.9 | 95.8 | 94.3 | 37.4 |
| Naïve Bayes | 49.1 | 95.6 | 42.1 | 96.6 | 92.7 | 47.5 |
| Neural network (multilayer perceptron) | 40.0 | 98.9 | 71.0 | 96.2 | 95.3 | 43.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raza, A.; Abbas, K.; Swangchan-Uthai, T.; Hogeveen, H.; Inchaisri, C. Behavioral Adaptations in Tropical Dairy Cows: Insights into Calving Day Predictions. Animals 2024, 14, 1834. https://doi.org/10.3390/ani14121834
Raza A, Abbas K, Swangchan-Uthai T, Hogeveen H, Inchaisri C. Behavioral Adaptations in Tropical Dairy Cows: Insights into Calving Day Predictions. Animals. 2024; 14(12):1834. https://doi.org/10.3390/ani14121834
Chicago/Turabian StyleRaza, Aqeel, Kumail Abbas, Theerawat Swangchan-Uthai, Henk Hogeveen, and Chaidate Inchaisri. 2024. "Behavioral Adaptations in Tropical Dairy Cows: Insights into Calving Day Predictions" Animals 14, no. 12: 1834. https://doi.org/10.3390/ani14121834
APA StyleRaza, A., Abbas, K., Swangchan-Uthai, T., Hogeveen, H., & Inchaisri, C. (2024). Behavioral Adaptations in Tropical Dairy Cows: Insights into Calving Day Predictions. Animals, 14(12), 1834. https://doi.org/10.3390/ani14121834