
Sucralose Influences the Productive Performance, Carcass Traits, Blood Components, and Gut Microflora Using 16S rRNA Sequencing of Growing APRI-Line Rabbits

 ,
,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Animals
2.2. Experimental Design
2.3. Data Collection
2.4. The Carcass Traits and Serum Blood Analysis
2.5. Digestibility and Cecum Bacteriology Experiment
2.6. Isolation of Gut Bacteria DNA and qPCR Analysis Based on 16s rRNA Gene
2.7. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Carcass Traits and Blood Components
3.3. The Digestibility Traits and Caecum Microbial Activity
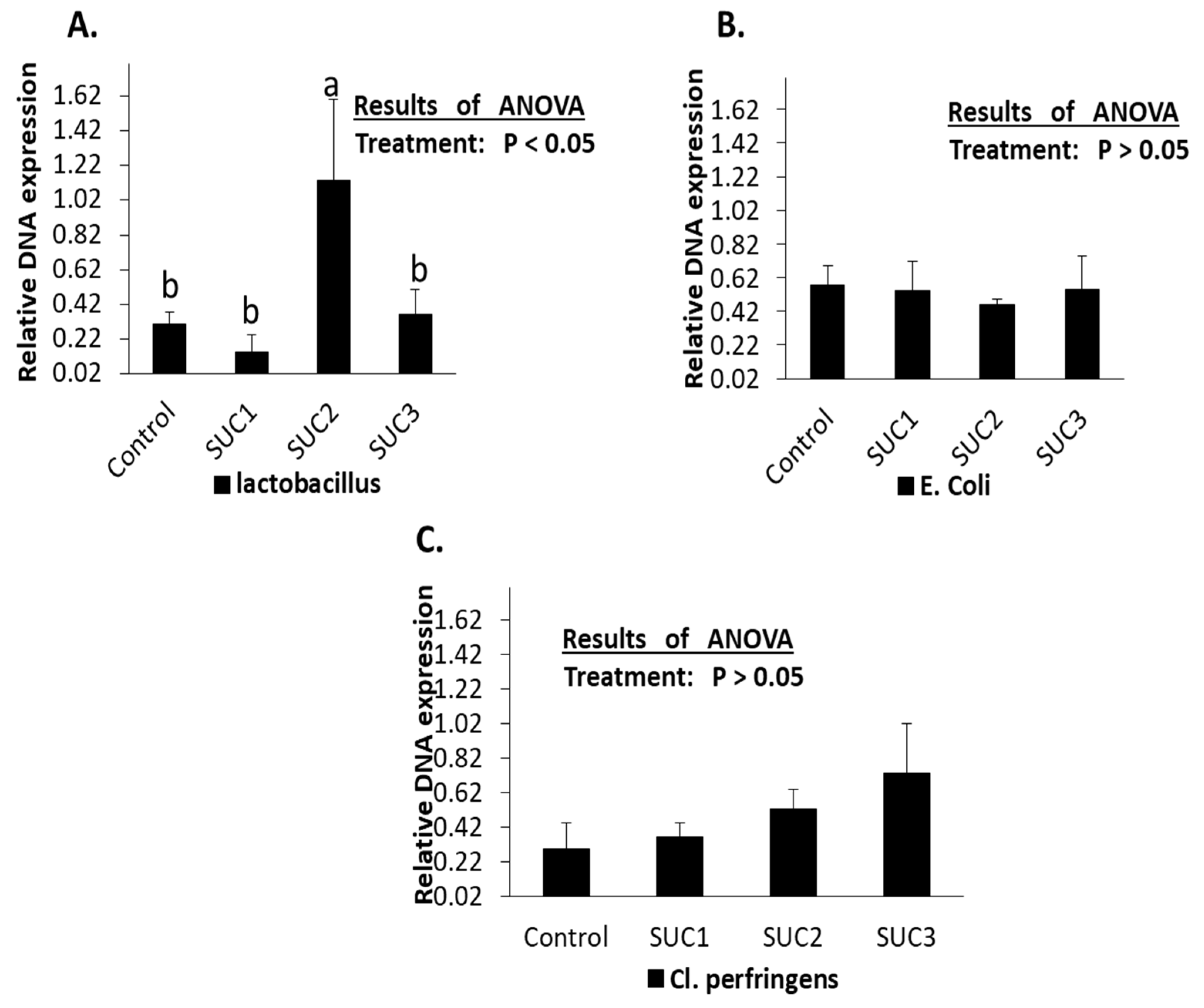
3.4. The qPCR Deferential of Lactobacillus, E. coli, and Cl. Perfringens in the Caecum of Rabbits after Sucralose Administration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amber, K.H.; Abd El-Nabi, F.; Morsy, W.A.; Morsy, S.H. Effect of dietary supplementation of probiotic and prebiotic on preventing post weaning digestive disorders and productive performance of growing rabbits. Egypt. Poult. Sci. 2014, 34, 19–38. [Google Scholar] [CrossRef]
- Berry, C.; Brusick, D.; Cohen, S.M.; Hardisty, J.F.; Grotz, V.L.; Williams, G.M. Sucralose Non-Carcinogenicity: A Review of the Scientific and Regulatory Rationale. Nutr. Cancer 2016, 68, 1247–1261. [Google Scholar] [CrossRef]
- Magnuson, B.A.; Roberts, A.; Nestmann, E.R. Critical review of the current literature on the safety of sucralose. Food Chem. Toxicol. 2017, 6, 324–355. [Google Scholar] [CrossRef]
- Mattes, R.D.; Popkin, B.M. Non-nutritive sweetener consumption in humans: Effects on appetite and food intake and their putative mechanisms. Am. J. Clin. Nutr. 2009, 89, 1–14. [Google Scholar] [CrossRef]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Sedghi, L.; DiMassa, V.; Harrington, A.; Lynch, S.V.; Kapila, Y.L. The oral microbiome: Role of key organisms and complex networks in oral health and disease. Periodontology 2000 2021, 87, 107–131. [Google Scholar] [CrossRef] [PubMed]
- Thomson, P.; Santibañez, R.; Aguirre, C.; Galgani, J.E.; Garrido, D. Short-term impact of sucralose consumption on the metabolic response and gut microbiome of healthy adults. Br. J. Nutr. 2019, 122, 856–862. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ojeda, F.J.; Plaza-Díaz, J.; Sáez-Lara, M.J.; Gil, A. Effects of Sweeteners on the Gut Microbiota: A Review of Experimental Studies and Clinical Trials. Adv. Nutr. 2019, 10, S31–S48. [Google Scholar] [CrossRef]
- Ahmad, S.Y.; Friel, J.; Mackay, D. The effects of non-nutritive artificial sweeteners, aspartame and sucralose, on the gut microbiome in healthy adults: Secondary outcomes of a randomized double-blinded crossover clinical trial. Nutrients 2020, 12, 3408. [Google Scholar] [CrossRef]
- Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef]
- Plaza-Diaz, J.; Pastor-Villaescusa, B.; Rueda, A.; Abadia-Molina, F.; Ruiz-Ojeda, F.J. Plausible Biological Interactions of Low- and Non-Calorie Sweeteners with the Intestinal Microbiota: An Update of Recent Studies. Nutrients 2020, 12, 1153. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Xiao, Y.; Ma, L.; Lyu, W.; Peng, H.; Wang, X.; Ren, Y.; Li, J. Low Dose of Sucralose Alter Gut Microbiome in Mice. Front. Nutr. 2022, 9, 848392. [Google Scholar] [CrossRef] [PubMed]
- Omran, A.; Baker, R.; Coughlin, C. Differential Bacteriostatic Effects of Sucralose on Various Species of Environmental Bacteria. Int. Sch. Res. Not. 2013, 2013, 415070. [Google Scholar] [CrossRef] [PubMed]
- del Pozo, S.; Gómez-Martínez, S.; Díaz, L.E.; Nova, E.; Urrialde, R.; Marcos, A. Potential Effects of Sucralose and Saccharin on Gut Microbiota: A Review. Nutrients 2022, 14, 1682. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.P.; Lin, Y.Q.; Zhang, L.; Wilson, Y.A.; Oyston, L.J.; Cotterell, J.; Qi, Y.; Khuong, T.M.; Bakhshi, N.; Planchenault, Y.; et al. Sucralose promotes food intake through NPY and a neuronal fasting response. Cell Metab. 2016, 24, 75–90. [Google Scholar] [CrossRef]
- Wang, Q.P.; Simpson, S.J.; Herzog, H.; Neely, G.G. Chronic sucralose or L-glucose ingestion does not suppress food intake. Cell Metab. 2017, 26, 279–280. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; He, H.; Gong, L.; Lai, W.; Dong, B.; Zhang, L. Effects of sweetener sucralose on diet preference, growth performance and hematological and biochemical parameters of weaned piglets. Asian-Australas. J. Anim. Sci. 2020, 33, 802–811. [Google Scholar] [CrossRef] [PubMed]
- El-Deghadi, A.; Elkassas, N.; Arafa, M.; Seif El-Naser, M. Evaluation of productive performance in synthetic maternal line (APRI rabbits) under egyptian conditions. Sinai J. Appl. Sci. 2022, 11, 727–738. [Google Scholar] [CrossRef]
- Eltahan, H.M.; Nguyen, L.T.N.; Han, G.; Yang, H.; Ali, M.N.; Amber, K.A.; Furuse, M.; Chowdhury, V.S. Hypothermia induced by central injection of sucralose potentially occurs via monoaminergic pathways in the hypothalamus of chicks. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 248, 110752. [Google Scholar] [CrossRef]
- De Blas, J.C.; Mateos, G.G. Feed Formulation. In The Nutrition of the Rabbit; De Blas, C., Wiseman, J., Eds.; CABI Publishing, CAB International: Oxfordshire, UK, 2020; pp. 243–253. [Google Scholar] [CrossRef]
- Blasco, A.; Ouhayoun, J.; Masoero, G. Hormonization of criteria and terminology in rabbit meat research. World Rabbit Sci. 1993, 1, 93–99. [Google Scholar] [CrossRef]
- Krunt, O.; Zita, L.; Kraus, A.; Bureš, D.; Needham, T.; Volek, Z. The effect of housing system on rabbit growth performance, carcass traits, and meat quality characteristics of different muscles. Meat Sci. 2022, 193, 108953. [Google Scholar] [CrossRef]
- Perez, J.M.; Lebas, F.; Gidenne, T.; Martens, L.; Xiccato, G.; Perigi-Bini, R.; Dallo- Zotte, A.; Cossu, M.E.; Carazzolo, A.; Villamide, M.J.; et al. European reference method for in vivo determination of diet digestibility in rabbits. World Rabbit Sci. 1995, 3, 41–43. [Google Scholar]
- AOAC. Association of Official Analytical Chemists. Official Methods of Analysis, 18th ed.; AOAC: Washington, DC, USA, 2005. [Google Scholar]
- Mackie, T.J.; McCartney, J.E. Handbook of Practical Bacteriology. A Guide Bacteriology Laboratory Work, 9th ed.; Livingstone Ltd.: Edinburgh, UK; London, UK, 1953. [Google Scholar]
- APHA. American Health Association. Standard Methods for the Examination of Dairy Products, 11th ed.; American Published Health Association: New York, NY, USA, 1960. [Google Scholar]
- Difco Mannual. Dehydrated Culture Media and Reagents, 8th ed.; Difco Laboratories: Detroit, MI, USA, 1977. [Google Scholar]
- Matsuki, T.; Watanabe, K.; Fujimoto, J.; Takada, T.; Tanaka, R. Use of 16S rRNA gene-targeted group-specific primers for real-time PCR analysis of predominant bacteria in human feces. Appl. Environ. Microbiol. 2004, 70, 7220–7228. [Google Scholar] [CrossRef] [PubMed]
- Eltahan, H.M.; Bahry, M.A.; Yang, H.; Han, G.; Nguyen, L.T.N.; Ikeda, H.; Ali, M.N.; Amber, K.A.; Furuse, M.; Chowdhury, V.S. Central NPY-Y5 sub-receptor partially functions as a mediator of NPY-induced hypothermia and affords thermotolerance in heat-exposed fasted chicks. Physiol. Rep. 2017, 23, e13511. [Google Scholar] [CrossRef] [PubMed]
- Furet, J.P.; Firmesse, O.; Gourmelon, M.; Bridonneau, C.; Tap, J.; Mondot, S.; Doré, J.; Corthier, G. Comparative assessment of human and farm animal faecal microbiota using real-time quantitative PCR. FEMS Microbiol. Ecol. 2009, 68, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Kramski, M.; Gaeguta, A.J.; Lichtfuss, G.F.; Rajasuriar, R.; Crowe, S.M.; French, M.A.; Lewin, S.R.; Center, R.J.; Purcell, D.F. Novel sensitive real-time PCR for quantification of bacterial 16S rRNA genes in plasma of HIV-infected patients as a marker for microbial translocation. J. Clin. Microbiol. 2011, 49, 3691–3693. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. NAR 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Bahry, M.A.; Yang, H.; Tran, P.V.; Do, P.H.; Han, G.; Eltahan, H.M.; Chowdhury, V.S.; Furuse, M. Reduction in voluntary food intake, but not fasting, stimulates hypothalamic gonadotropin-inhibitory hormone precursor mRNA expression in chicks under heat stress. Neuropeptides 2018, 71, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Eltahan, H.M.; Kang, C.W.; Chowdhury, V.S.; Eltahan, H.M.; Abdel-Maksoud, M.A.; Mubarak, A.; Lim, C.I. Cold Drinking Water Boosts the Cellular and Humoral Immunity in Heat-Exposed Laying Hens. Animals 2023, 13, 580. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute, Inc. SAS User’s Guide: Statistics; SAS Inst. Inc.: Cary, NC, USA, 2000. [Google Scholar]
- Duncan, D.B. Multiple range and multiple F Tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Kobayashi, K.; Pillai, K.S. Transformation of data and outliers. In A Handbook of Applied Statistics in Pharmacology; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2013; pp. 37–46. [Google Scholar]
- Ahn, S.R.; An, J.H.; Song, H.S.; Park, J.W.; Lee, S.H.; Kim, J.H.; Jang, J.; Park, T.H. Duplex Bioelectronic Tongue for sensing umami and sweet tastes based on human taste receptor Nanovesicles. ACS Nano 2016, 10, 7287–7296. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, L.A. Acute and subchronic toxicity of sucralose. Food Chem. Toxicol. 2000, 38, 53–69. [Google Scholar] [CrossRef] [PubMed]
- Kille, J.W.; Ford, W.C.; McAnulty, P.; Tesh, J.M.; Ross, F.W.; Willoughby, C.R. Sucralose: Lack of effects on sperm glycolysis and reproduction in the rat. Food Chem. Toxicol. 2000, 38, S19–S29. [Google Scholar] [CrossRef] [PubMed]
- Soffritti, M.; Padovani, M.; Tibaldi, E.; Falconi, L.; Manservisi, F.; Lauriola, M.; Bua, L.; Manservigi, M.; Belpoggi, F. Sucralose administered in feed beginning prenatally through lifespan, induces hematopoietic neoplasias in male Swiss mice. Int. J. Occup. Environ. Health 2016, 22, 7–17. [Google Scholar] [CrossRef]
- Saada, H.N.; Mekky, N.H.; Eldawy, H.A.; Abdelaal, A.F. Biological effect of sucralose in diabetic rats. Food Nutr. Sci. 2013, 4, 82–89. [Google Scholar] [CrossRef]
- Sahin, K.; Orhan, C.; Tuzcu, M.; Ali, S.; Sahin, N.; Hayirli, A. Epigallocatechin-3-gallate prevents lipid peroxidation and enhances antioxidant defense system via modulating hepatic nuclear transcription factors in heat-stressed quails. Poult. Sci. 2010, 89, 2251–2258. [Google Scholar] [CrossRef] [PubMed]
- Erbaş, O.; Erdoğan, M.A.; Khalilnezhad, A.; Solmaz, V.; Gürkan, F.T.; Yiğittürk, G.; Eroglu, H.A.; Taskiran, D. Evaluation of long-term effects of artificial sweeteners on rat brain: A biochemical, behavioral, and histological study. J. Biochem. Mol. Toxicol. 2018, 32, e22053. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Liu, S.; Jamal, T.; Ding, T.; Qi, L.; Lv, Z.; Yu, D.; Shi, F. Effects of dietary sweeteners supplementation on growth performance, serum biochemicals, and jejunal physiological functions of broiler chickens. Poult. Sci. 2020, 99, 3948–3958. [Google Scholar] [CrossRef]
- Schiffman, S.S.; Scholl, E.H.; Furey, T.S.; Troy, N.H. Toxicological and pharmacokinetic properties of sucralose-6-acetate and its parent sucralose: In vitro screening assays. J. Toxicol. Environ. Health B 2023, 26, 307–341. [Google Scholar] [CrossRef]
- Bian, X.; Chi, L.; Gao, B.; Tu, P.; Ru, H.; Lu, K. Gut Microbiome Response to Sucralose and Its Potential Role in Inducing Liver Inflammation in Mice. Front. Physiol. 2017, 8, 487. [Google Scholar] [CrossRef]
- Abou-Donia, M.B.; El-Masry, E.M.; Abdel-Rahman, A.A.; McLendon, R.E.; Schiffman, S.S. Splenda alters gut microflora and increases intestinal P-glycoprotein and cytochrome P-450 in male rats. J. Toxicol. Environ. Health 2008, 71, 1415–1429. [Google Scholar] [CrossRef] [PubMed]
- Uebanson, T.; Ohnishi, A.; Kitayama, R.; Yoshimoto, A.; Nakahashi, M.; Shimohata, T.; Mawatari, K.; Takahashi, A. Effects of Low-Dose Non-Caloric Sweetener Consumption on Gut Microbiota in Mice. Nutrients 2017, 9, 560. [Google Scholar] [CrossRef] [PubMed]
- Corder, B.; Knobbe, A. The effects of the artificial sweetener sucralose on the gut bacteria Escherichia coli and Enterobacter aerogenes. J. Exp. Microbiol. Immunol. 2018, 4, 1–9. [Google Scholar]
- Shil, A.; Chichger, H. Artificial Sweeteners Negatively Regulate Pathogenic Characteristics of Two Model Gut Bacteria, E. coli and E. faecalis. Int. J. Mol. Sci. 2021, 22, 5228. [Google Scholar] [CrossRef]
- Sánchez-Tapia, M.; Miller, A.W.; Granados-Portillo, O.; Tovar, A.R.; Torres, N. The development of metabolic endotoxemia is dependent on the type of sweetener and the presence of saturated fat in the diet. Gut Microbes 2020, 12, 1840766. [Google Scholar] [CrossRef]
- Wang, Q.-P.; Browman, D.; Herzog, H.; Neely, G.G. Non-nutritive sweeteners possess a bacteriostatic effect and alter gut microbiota in mice. PLoS ONE 2018, 13, e0199080. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The microbiota-gut-brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Ingredients | % | Chemical Analysis (% as DM): | % |
|---|---|---|---|
| Berseem hay | 30.05 | Dry matter (DM) | 85.81 |
| Barley grain | 24.60 | Crude protein (CP) | 17.36 |
| Wheat brain | 21.50 | Organic matter | 91.42 |
| Soybean meal (44% CP) | 17.50 | Crude fibre | 12.37 |
| Molasses | 3.00 | Ether extract | 2.230 |
| Limestone | 0.95 | Digestible energy (MJ/kg DM) | 10.11 |
| Di-calcium phosphate | 1.60 | Calcium | 1.243 |
| Sodium chloride | 0.30 | Phosphorus | 0.808 |
| Mineral–vitamin premix (1) | 0.30 | Methionine | 0.454 |
| DL-Methionine | 0.20 | Lysine | 0.862 |
| Nitrogen-free extract (NFE) | 56.7 | ||
| Ash 8.6 ADF 16.25 | |||
| NDF 28.75 ADL 3.1 | |||
| Total | 100 | ||
| Gene | Accession No. | Sequences 5′−3′ (Forward/Reverse) | Annealing Temperature (°C) | Product Size (bp) | Reference |
|---|---|---|---|---|---|
| All bacteria # | _ | 5′−CGGTGAATACGTTCCCGG−3′/ 5′−TACGGCTACCTTGTTACGACTT−3′ | 60 | 145 | Furet et al. [30] |
| Cl. perfringens | NR_121697.2 | 5′−AAGATGGCATCATCATTCAACCA−3′/ 5′−GTGCAATATTCCCCACTGCTGCCT-3′ | 60 | 188 | This study |
| E. coli | J01859.1 | 5′−AAGACCAAAGAGGGGGACCT−3′/ 5′−TGTCTCAGTTCCAGTGTGGC−3′ | 60 | 141 | Kramski et al., [31] |
| Lactobacillus * | NC_004957.1 | 5′−GGCAGAGCCTCCATAAGCAA−3′/ 5′−GACGATTTTTCGTCTCGGCG−3′ | 60 | 278 | This study |
| Parameters | Control | SUC1 | SUC2 | SUC3 | SEM | p Value | Linear p Value | Quadratic p Value |
|---|---|---|---|---|---|---|---|---|
| No. of rabbits at 5 weeks of age | 20 | 20 | 20 | 20 | - | - | - | - |
| Final number at 13 weeks of age | 19 | 19 | 19 | 18 | - | - | - | - |
| Initial body weight (g) | 581.2 | 581.2 | 582.0 | 580.0 | 6.078 | 0.89 | 0.855 | 0.8401 |
| Final body weight (g) | 2158.2 A | 2132.7 A | 2084.7 B | 2061.5 B | 12.28 | 0.007 | 0.00455 | 0.2301 |
| Daily weight gain (g): | ||||||||
| 5–9 weeks | 28.80 A | 27.82 AB | 27.04 BC | 26.38 C | 0.488 | 0.009 | 0.0680 | 0.802 |
| 9–13 weeks | 27.42 | 27.64 | 26.63 | 26.54 | 0.624 | 0.41 | 0.8.80 | 0.4301 |
| 5–13 weeks | 28.11 A | 27.73 A | 26.84 B | 26.46 B | 0.218 | 0.005 | 4.80 × 105 | 0.2012 |
| Feed intake (g/d): | ||||||||
| 5–9 weeks | 80.08 A | 74.06 B | 73.22 BC | 71.16 C | 0.703 | 0.009 | 2.80 × 107 | 1 × 103 |
| 9–13 weeks | 113.5 A | 106.1 B | 104.7 B | 102.6 B | 1.173 | 0.007 | 6.80 × 105 | 0.0802 |
| 5–13 weeks | 96.78 A | 90.07 B | 88.96 BC | 86.89 C | 0.874 | 0.005 | 5.80 × 106 | 4 × 103 |
| Feed conversion ratio: | ||||||||
| 5–9 weeks | 2.806 | 2.675 | 2.725 | 2.703 | 0.060 | 0.07 | 0.693 | 0.4329 |
| 9–13 weeks | 4.184 A | 3.876 B | 3.958 AB | 3.894 B | 0.080 | 0.031 | 0.0393 | 0.1054 |
| 5–13 weeks | 3.442 A | 3.250 B | 3.317 B | 3.288 B | 0.027 | 0.006 | 2.93 × 106 | 0.2019 |
| Water consumption (mL/d): | ||||||||
| 5–9 weeks | 132.6 | 131.2 | 130.7 | 129.8 | 2.108 | 0.42 | 0.4980 | 0.9571 |
| 9–13 weeks | 211.2 | 210.4 | 208.4 | 207.2 | 2.383 | 0.76 | 0.1745 | 0.9427 |
| 5–13 weeks | 171.9 | 170.8 | 169.6 | 168.5 | 1.654 | 0.41 | 0.2110 | 0.9851 |
| Performance index (%) | 62.77 B | 65.75 A | 62.97 B | 62.82 B | 0.642 | 0.045 | 0.0593 | 0.2149 |
| Total edible (%) BW | 59.02 A | 56.84 B | 56.41 B | 55.97 B | 0.438 | 0.008 | 0.00455 | 0.148061 |
| GIT (1) (%) BW | 12.64 C | 13.93 B | 14.03 AB | 14.09 AB | 0.324 | 0.004 | 0.0293 | 0.1229 |
| Mortality rate (%) (2) | 5 | 5 | 5 | 10 | - | - | - | - |
| Parameter | Control | SUC1 | SUC2 | SUC3 | SEM | p-Value | Linear p Value | Quadratic p Value |
|---|---|---|---|---|---|---|---|---|
| Total protein (g/dL) | 6.08 | 5.92 | 5.86 | 5.80 | 0.145 | 0.15 | 0.184 | 0.72605 |
| Glucose (mg/dL) | 89.33 A | 80.00 B | 77.73 B | 68.33 C | 1.919 | 0.004 | 9.27 × 107 | 0.98128 |
| Cholesterol (mg/dL) | 69.95 B | 71.88 B | 73.95 A | 76.38 A | 0.831 | 0.007 | 2.75 × 105 | 0.72519 |
| H.D.L. (mg/dL) | 33.15 B | 34.22 A | 34.84 A | 35.09 A | 0.184 | 0.004 | 4.08 × 104 | 0.15381 |
| L.D.L. (mg/dL) | 30.15 C | 30.69 BC | 31.12 AB | 31.75 A | 0.136 | 0.008 | 7.81 × 104 | 0.63631 |
| Total lipids (g/L) | 294.0 C | 301.7 BC | 309.3 B | 323.0 A | 3.804 | 0.006 | 4.41 × 105 | 0.35780 |
| Triglyceride (mg/dL) | 90.05 A | 88.09 B | 86.44 C | 85.12 D | 0.614 | 0.006 | 4.60 × 105 | 0.58383 |
| Creatinine (mg/dL) | 0.95 | 0.91 | 0.91 | 0.92 | 0.016 | 0.08 | 0.274 | 0.07555 |
| Urea (mg/dL) | 15.25 | 15.10 | 14.95 | 14.91 | 0.207 | 0.09 | 0.00347 | 0.12595 |
| AST (U/L) | 22.91 | 22.39 | 22.70 | 22.91 | 0.721 | 0.74 | 0.921 | 0.62929 |
| ALT (U/L) | 15.59 | 14.69 | 14.71 | 15.17 | 0.693 | 0.85 | 0.724 | 0.40206 |
| Malonaldehyde (mg/mL) | 1.108 C | 1.155 C | 1.284 B | 1.415 A | 0.028 | 0.009 | 1.03 × 104 | 0.29308 |
| Nutrient | Control | SUC1 | SUC2 | SUC3 | SEM | p-Value | Linear p Value | Quadratic p Value |
|---|---|---|---|---|---|---|---|---|
| Dry matter (DM) | 71.15 | 71.16 | 71.38 | 71.50 | 0.443 | 0.75 | 0.409 | 0.878 |
| Organic matter (OM) | 66.09 | 66.56 | 66.72 | 66.75 | 0.704 | 0.45 | 0.519 | 0.774 |
| Crude protein (CP) | 75.68 A | 74.36 BC | 75.22 AB | 73.78 C | 0.316 | 0.054 | 0.0280 | 0.902 |
| Crude fibre (CF) | 30.38 | 30.69 | 30.94 | 31.08 | 0.491 | 0.42 | 0.662 | 0.946 |
| Ether extract (EE) | 78.55 | 78.11 | 77.81 | 77.63 | 0.652 | 0.86 | 0.578 | 0.916 |
| Nitrogen-free extract (NFE) | 51.47 | 52.29 | 52.87 | 53.22 | 0.714 | 0.12 | 0.255 | 0.837 |
| Parameters | Control | SUC1 | SUC2 | SUC3 | SEM | p-Value | Linear p Value | Quadratic p Value |
|---|---|---|---|---|---|---|---|---|
| No. of rabbits | 4 | 4 | 4 | 4 | - | - | ||
| pH | 6.72 | 6.52 | 6.32 | 6.35 | 0.104 | 0.95 | 0.0599 | 0.458 |
| NH3 (mmol/L) | 8.20 A | 5.35 B | 3.22 C | 3.32 C | 0.234 | 0.007 | 9.79 × 106 | 3.19 × 103 |
| Total bacterial count (×106) (1) | 12.2 C | 16.12 B | 24.85 A | 23.75 A | 1.862 | 0.04 | 9.06 × 107 | 0.0259 |
| Lactobacilli (×105) (1) | 2.25 D | 14.45 C | 20.25 A | 16.45 B | 0.371 | 0.004 | 2.32 × 104 | 2.71 × 109 |
| Escherichia coli (×104) (1) | 12.92 A | 7.42 B | 3.28 C | 3.42 C | 0.649 | 0.007 | 3.77 × 107 | 1.04 × 106 |
| Clostridium spp. (1) | 2.12 C | 4.42 B | 2.52 C | 7.35 A | 0.406 | 0.008 | 1.86 × 103 | 0.120 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Tahan, H.M.; Elmasry, M.E.; Madian, H.A.; Alhimaidi, A.R.; Kim, I.H.; Park, J.H.; El-Tahan, H.M. Sucralose Influences the Productive Performance, Carcass Traits, Blood Components, and Gut Microflora Using 16S rRNA Sequencing of Growing APRI-Line Rabbits. Animals 2024, 14, 1925. https://doi.org/10.3390/ani14131925
El-Tahan HM, Elmasry ME, Madian HA, Alhimaidi AR, Kim IH, Park JH, El-Tahan HM. Sucralose Influences the Productive Performance, Carcass Traits, Blood Components, and Gut Microflora Using 16S rRNA Sequencing of Growing APRI-Line Rabbits. Animals. 2024; 14(13):1925. https://doi.org/10.3390/ani14131925
Chicago/Turabian StyleEl-Tahan, Hatem M., Mohamad Elsayed Elmasry, H. A. Madian, Ahmad R. Alhimaidi, In Ho Kim, Jae Hong Park, and Hossam M. El-Tahan. 2024. "Sucralose Influences the Productive Performance, Carcass Traits, Blood Components, and Gut Microflora Using 16S rRNA Sequencing of Growing APRI-Line Rabbits" Animals 14, no. 13: 1925. https://doi.org/10.3390/ani14131925