

Chromium Affects Mitochondrial Function, Leading to Apoptosis and Autophagy in Turtle Primary Hepatocytes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Culture and Processing
2.3. Cell Growth Assay
2.4. Cytotoxicity Assay
2.5. Oxidative Stress Detection
2.6. Reactive Oxygen Species (ROS)
2.7. Apoptosis Detection
2.8. ATP Level Detection
2.9. qRT-PCR Analysis
2.10. Statistic Analysis
3. Results
3.1. Cell Growth Curve and Morphology of Primary Hepatocytes
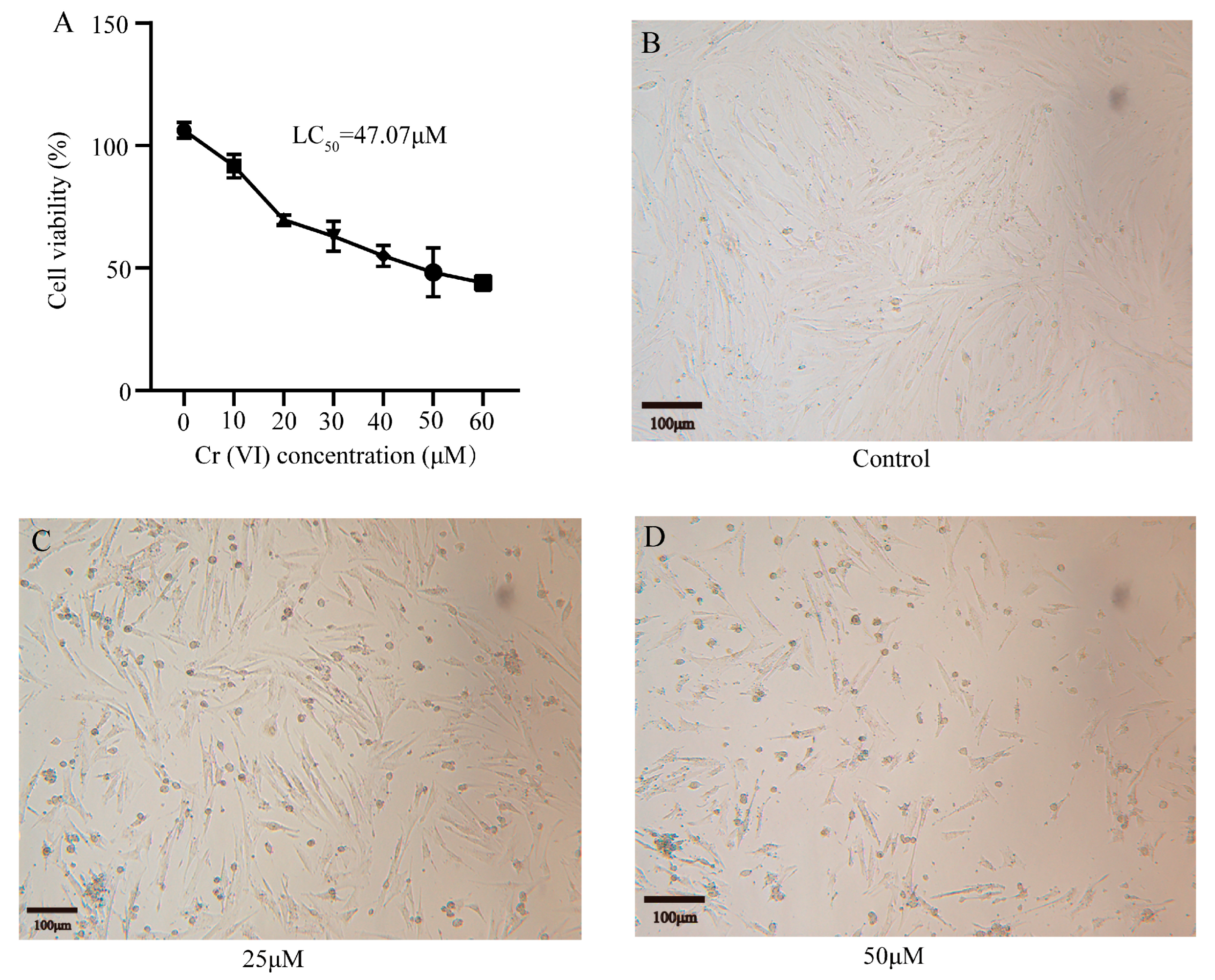
3.2. Cr(VI) Inhibited the Growth of Turtle Hepatocytes
3.3. Cr(VI) Induced Oxidative Stress in Turtle Hepatocytes
3.4. Cr(VI) Decreased Mitochondrial Dynamics in Hepatocytes
3.5. Cr(VI) Activated the p53 Signal Pathway in the Liver Cells
3.6. Cr(VI) Induced Apoptosis in Turtle Hepatocytes
3.7. Cr(VI) Caused Autophagy in Hepatocyte
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ray, P.D.; Huang, B.W.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell. Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef] [PubMed]
- Keane, M.; Siert, A.; Stone, S.; Chen, B.T. Profiling stainless steel welding processes to reduce fume emissions, hexavalent chromium emissions and operating costs in the workplace. J. Occup. Environ. Hyg. 2016, 13, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.F.; Zhang, N.; Liu, S.Y.; Ding, J.; Xu, W. Recent research progress of hexavalent chromium Cr(VI). Appl. Chem. Ind. 2020, 49, 1035–1038+1043. [Google Scholar] [CrossRef]
- Yi, M.; Yi, H.L.; Wu, L.H. Accumulation and toxicity of hexavalent chromium in mice. Asian J. Ecotoxicol. 2017, 12, 259–265. [Google Scholar] [CrossRef]
- Ray, R.R. Adverse hematological effects of hexavalent chromium: An overview. Interdiscip. Toxicol. 2016, 9, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, J.J.; Bansal, N.; Chirwa, E.M.N. Chromium in environment, its toxic effect from chromite-mining and ferrochrome industries, and its possible bioremediation. Expo. Health 2020, 12, 51–62. [Google Scholar] [CrossRef]
- Wu, X.; Ye, T.; Xie, C.; Li, K.; Liu, C.; Yang, Z.; Han, R.; Wu, H.; Wang, Z. Experimental and modeling study on Cr(VI) migration from slag into soil and groundwater. Processes 2022, 10, 2235. [Google Scholar] [CrossRef]
- Proctor, D.M.; Suh, M.; Campleman, S.L.; Thompson, C.M. Assessment of the mode of action for hexavalent chromium-induced lung cancer following inhalation exposures. Toxicology 2014, 325, 160–179. [Google Scholar] [CrossRef] [PubMed]
- Szecsody, J.E.; Truex, M.J.; Qafoku, N.P.; McKinley, J.P.; Ivarson, K.A.; Di Pietro, S. Persistence of chromate in vadose zone and aquifer sediments in Hanford, Washington. Sci. Total Environ. 2019, 676, 482–492. [Google Scholar] [CrossRef]
- Remy, L.L.; Byers, V.; Clay, T. Reproductive outcomes after non-occupational exposure to hexavalent chromium, Willits California, 1983–2014. Environ. Health A Glob. Access Sci. Source 2017, 16, 18. [Google Scholar] [CrossRef]
- Wang, X.; Li, L.; Yan, X.; Meng, X.; Chen, Y. Processes of chromium (VI) migration and transformation in chromate production site: A case study from the middle of China. Chemosphere 2020, 257, 127282. [Google Scholar] [CrossRef] [PubMed]
- Eleftheriou, E.P.; Adamakis, I.D.; Melissa, P. Effects of hexavalent chromium on microtubule organization, ER distribution and callose deposition in root tip cells of Allium cepa L. Protoplasma 2012, 249, 401–416. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.S.; Han, B.; Al-Mizan Pichtel, J. Assessment of soil and groundwater contamination at a former Tannery district in Dhaka, Bangladesh. Environ. Geochem. Health 2020, 42, 1905–1920. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.L.; Liu, Z.M.; Liu, Y. Effects of lead, cadmium, and chromium contents in Xiangjiang River on freshwater fish. Food Mach. 2013, 29, 68–70, 216. [Google Scholar]
- Zhang, Y.; Li, C.; Zhang, J.; Wang, Y. Interpretation on the Standards for drinking water quality (GB 5749—2022). Water Technol. 2022, 16, 38–43. [Google Scholar]
- Cao, Q.; Wang, X.R. Study on pollution characteristics of contaminated sites with chrome and discussion on common remediation technologies. Chin. J. Environ. Eng. 2009, 8, 1493–1497. [Google Scholar]
- Sun, H.; Brocato, J.; Costa, M. Oral Chromium exposure and toxicity. Curr. Environ. Health Rep. 2015, 2, 295–303. [Google Scholar] [CrossRef]
- Xiao, F.; Feng, X.; Zeng, M.; Guan, L.; Hu, Q.; Zhong, C. Hexavalent chromium induces energy metabolism disturbance and p53-dependent cell cycle arrest via reactive oxygen species in L-02 hepatocytes. Mol. Cell. Biochem. 2012, 371, 65–76. [Google Scholar] [CrossRef]
- Kirman, C.R.; Aylward, L.L.; Suh, M.; Harris, M.A.; Thompson, C.M.; Haws, L.C.; Proctor, D.M.; Lin, S.S.; Parker, W.; Hays, S.M. Physiologically based pharmacokinetic model for humans orally exposed to chromium. Chem.-Biol. Interact. 2013, 204, 13–27. [Google Scholar] [CrossRef]
- Zhang, X.H.; Zhang, X.; Wang, X.C.; Jin, L.F.; Yang, Z.P.; Jiang, C.X.; Chen, Q.; Ren, X.B.; Cao, J.Z.; Wang, Q.; et al. Chronic occupational exposure to hexavalent chromium causes DNA damage in electroplating workers. BMC Public Health 2011, 11, 224. [Google Scholar] [CrossRef]
- Wise, J.P., Sr.; Wise, S.S.; Little, J.E. The cytotoxicity and genotoxicity of particulate and soluble hexavalent chromium in human lung cells. Mutat. Res. 2002, 517, 221–229. [Google Scholar] [CrossRef]
- O’Brien, T.J.; Ceryak, S.; Patierno, S.R. Complexities of chromium carcinogenesis: Role of cellular response, repair and recovery mechanisms. Mutat. Res. 2003, 533, 3–36. [Google Scholar] [CrossRef]
- Bagchi, D.; Vuchetich, P.J.; Bagchi, M.; Hassoun, E.A.; Tran, M.X.; Tang, L.; Stohs, S.J. Induction of oxidative stress by chronic administration of sodium dichromate [chromium VI] and cadmium chloride [cadmium II] to rats. Free Radic. Biol. Med. 1997, 22, 471–478. [Google Scholar] [CrossRef]
- Velma, V.; Tchounwou, P.B. Chromium-induced biochemical, genotoxic and histopathologic effects in liver and kidney of goldfish, carassius auratus. Mutat. Res. 2010, 698, 43–51. [Google Scholar] [CrossRef]
- Wise, S.S.; Xie, H.; Fukuda, T.; Douglas Thompson, W.; Wise, J.P. Hexavalent chromium is cytotoxic and genotoxic to hawksbill sea turtle cells. Toxicol. Appl. Pharmacol. 2014, 279, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Speer, R.M.; Wise, S.S.; Croom-Perez, T.J.; Aboueissa, A.M.; Martin-Bras, M.; Barandiaran, M.; Bermúdez, E.; Wise, J.P., Sr. A comparison of particulate hexavalent chromium cytotoxicity and genotoxicity in human and leatherback sea turtle lung cells from a one environmental health perspective. Toxicol. Appl. Pharmacol. 2019, 376, 70–81. [Google Scholar] [CrossRef]
- Webb, S.J.; Zychowski, G.V.; Bauman, S.W.; Higgins, B.M.; Raudsepp, T.; Gollahon, L.S.; Wooten, K.J.; Cole, J.M.; Godard-Codding, C. Establishment, characterization, and toxicological application of loggerhead sea turtle (Caretta caretta) primary skin fibroblast cell cultures. Environ. Sci. Technol. 2014, 48, 14728–14737. [Google Scholar] [CrossRef]
- Finlayson, K.A.; Leusch FD, L.; van de Merwe, J.P. Cytotoxicity of organic and inorganic compounds to primary cell cultures established from internal tissues of Chelonia mydas. Sci. Total Environ. 2019, 664, 958–967. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.N.; Duan, H.R.; Wei, Y.F.; Ge, Y.G.; Shi, P. In vitro vulture of red-eared slider embryonic fibroblasts. Chin. J. Zool. 2018, 53, 733–741. [Google Scholar] [CrossRef]
- Yu, X.; Yu, R.Q.; Gui, D.; Zhang, X.; Zhan, F.; Sun, X.; Wu, Y. Hexavalent chromium induces oxidative stress and mitochondria-mediated apoptosis in isolated skin fibroblasts of Indo-Pacific humpback dolphin. Aquat. Toxicol. 2018, 203, 179–186. [Google Scholar] [CrossRef]
- Wang, C.; Dai, X.; Xing, C.; Zhang, C.; Cao, H.; Guo, X.; Liu, P.; Yang, F.; Zhuang, Y.; Hu, G. Hexavalent-chromium-induced disruption of mitochondrial dynamics and apoptosis in the liver via the AMPK-PGC-1α pathway in ducks. Int. J. Mol. Sci. 2023, 24, 17241. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.H.; Venuprasad, M.P.; Jayashree, G.V.; Rachitha, P.; Krupashree, K.; Pal, A.; Khanum, F. Celastrus paniculatus Willd. mitigates t-BHP induced oxidative and apoptotic damage in C2C12 murine muscle cells. Cytotechnology 2015, 67, 955–967. [Google Scholar] [CrossRef] [PubMed]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef]
- Liang, Q.; Zhang, Y.; Huang, M.; Xiao, Y.; Xiao, F. Role of mitochondrial damage in Cr(VI)-induced endoplasmic reticulum stress in L-02 hepatocytes. Mol. Med. Rep. 2019, 19, 1256–1265. [Google Scholar] [CrossRef]
- Huang, Q.; Chen, H.; Li, J.; Oliver, M.; Ma, X.; Byck, D.; Gao, Y.; Jiang, S.W. Epigenetic and non-epigenetic regulation of syncytin-1 expression in human placenta and cancer tissues. Cell. Signal. 2014, 26, 648–656. [Google Scholar] [CrossRef]
- Yang, Q.; Han, B.; Xue, J.; Lv, Y.; Li, S.; Liu, Y.; Wu, P.; Wang, X.; Zhang, Z. Hexavalent chromium induces mitochondrial dynamics disorder in rat liver by inhibiting AMPK/PGC-1α signaling pathway. Environ. Pollut. 2020, 265 Pt A, 114855. [Google Scholar] [CrossRef]
- Zhang, H.; Yan, A.; Liu, X.; Ma, Y.; Zhao, F.; Wang, M.; Loor, J.J.; Wang, H. Melatonin ameliorates ochratoxin a induced liver inflammation, oxidative stress and mitophagy in mice involving in intestinal microbiota and restoring the intestinal barrier function. J. Hazard. Mater. 2021, 407, 124489. [Google Scholar] [CrossRef]
- Chtourou, Y.; Aouey, B.; Kebieche, M.; Fetoui, H. Protective role of naringin against cisplatin induced oxidative stress, inflammatory response and apoptosis in rat striatum via suppressing ROS-mediated NF-κB and P53 signaling pathways. Chem.-Biol. Interact. 2015, 239, 76–86. [Google Scholar] [CrossRef]
- Xu, G.; Liu, S.; Huang, M.; Jiang, X.; Yang, M. Cadmium induces apoptosis of human granulosa cell line KGN via mitochondrial dysfunction-mediated pathways. Ecotoxicol. Environ. Saf. 2021, 220, 112341. [Google Scholar] [CrossRef]
- Huang, Q.; Su, H.; Qi, B.; Wang, Y.; Yan, K.; Wang, X.; Li, X.; Zhao, D. A SIRT1 activator, ginsenoside Rc, promotes energy metabolism in cardiomyocytes and neurons. J. Am. Chem. Soc. 2021, 143, 1416–1427. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xiang, H.; Liu, J.; Chen, Y.; He, R.R.; Liu, B. Mitochondrial sirtuin 3: New emerging biological function and therapeutic target. Theranostics 2020, 10, 8315–8342. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Zhao, Q.; Sun, X.; Qian, H.; Lyu, S.; Chen, R.; Xia, H.; Yuan, W. Sirtuin 3: Emerging therapeutic target for cardiovascular diseases. Free Radic. Biol. Med. 2022, 180, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Li, S.; Li, J.; Lv, Y.; Wang, X.; Wu, P.; Yang, Q.; Tang, Y.; Liu, Y.; Zhang, Z. Hexavalent chromium induces renal apoptosis and autophagy via disordering the balance of mitochondrial dynamics in rats. Ecotoxicol. Environ. Saf. 2020, 204, 111061. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Alvarez, M.I.; Sebastián, D.; Vives, S.; Ivanova, S.; Bartoccioni, P.; Kakimoto, P.; Plana, N.; Veiga, S.R.; Hernández, V.; Vasconcelos, N.; et al. Deficient endoplasmic reticulum-mitochondrial phosphatidylserine transfer causes liver disease. Cell 2019, 177, 881–895.e17. [Google Scholar] [CrossRef]
- Obeng, E. Apoptosis (programmed cell death) and its signals—A review. Braz. J. Biol. Rev. Brasleira Biol. 2021, 81, 1133–1143. [Google Scholar] [CrossRef]
- Cory, S.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nature reviews. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef]
- Mizushima, N.; Levine, B.; Cuervo, A.M.; Klionsky, D.J. Autophagy fights disease through cellular self-digestion. Nature 2008, 451, 1069–1075. [Google Scholar] [CrossRef]
- Allaire, M.; Rautou, P.E.; Codogno, P.; Lotersztajn, S. Autophagy in liver diseases: Time for translation? J. Hepatol. 2019, 70, 985–998. [Google Scholar] [CrossRef]
- Shi, M.; Chen, L.; Shang, C.S.; Du, J.Q.; Feng, X.H. Advances in the correlation between autophagy and aging. Chin. J. Aesthetic Med. 2018, 27, 136–140. [Google Scholar] [CrossRef]
- Zhang, H.; Zheng, Y.; Liu, X.; Zha, X.; Elsabagh, M.; Ma, Y.; Jiang, H.; Wang, H.; Wang, M. Autophagy attenuates placental apoptosis, oxidative stress and fetal growth restriction in pregnant ewes. Environ. Int. 2023, 173, 107806. [Google Scholar] [CrossRef]
- Huang, Y.; Liu, H.T.; Yuan, Y.; Guo, Y.P.; Wan, D.F.; Pan, S.S. Exercise preconditioning increases beclin1 and induces autophagy to promote early myocardial protection via intermittent myocardial ischemia-hypoxia. Int. Heart J. 2021, 62, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Rubio, R.M.; Mohr, I. Inhibition of ULK1 and Beclin1 by an α-herpesvirus Akt-like Ser/Thr kinase limits autophagy to stimulate virus replication. Proc. Natl. Acad. Sci. USA 2019, 116, 26941–26950. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Li, Y.; Sheng, G.; Meng, Q.; Hu, Q.; Gao, X.; Shang, Z.; Lv, Q. Sirtuin 1 promotes autophagy and proliferation of endometrial cancer cells by reducing acetylation level of LC3. Cell Biol. Int. 2021, 45, 1050–1059. [Google Scholar] [CrossRef] [PubMed]
- Akcora Yildiz, D.; Irtegun Kandemir, S.; Agacayak, E.; Deveci, E. Evaluation of protein levels of autophagy markers (Beclin 1 and SQSTM1/p62) and phosphorylation of cyclin E in the placenta of women with preeclampsia. Cell. Mol. Biol. 2017, 63, 51–55. [Google Scholar] [CrossRef]
- Li, H.; Shi, J.; Gao, H.; Yang, X.; Fu, Y.; Peng, Y.; Xia, Y.; Zhou, D. Hexavalent chromium causes apoptosis and autophagy by inducing mitochondrial dysfunction and oxidative stress in broiler cardiomyocytes. Biol. Trace Elem. Res. 2022, 200, 2866–2875. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, S.; Xiao, Y.; Lin, J.; Yuan, Y.; Shi, H.; Hong, M.; Ding, L. Chromium Affects Mitochondrial Function, Leading to Apoptosis and Autophagy in Turtle Primary Hepatocytes. Animals 2024, 14, 2403. https://doi.org/10.3390/ani14162403
Lin S, Xiao Y, Lin J, Yuan Y, Shi H, Hong M, Ding L. Chromium Affects Mitochondrial Function, Leading to Apoptosis and Autophagy in Turtle Primary Hepatocytes. Animals. 2024; 14(16):2403. https://doi.org/10.3390/ani14162403
Chicago/Turabian StyleLin, Shuqin, Yunjuan Xiao, Jing Lin, Yue Yuan, Haitao Shi, Meiling Hong, and Li Ding. 2024. "Chromium Affects Mitochondrial Function, Leading to Apoptosis and Autophagy in Turtle Primary Hepatocytes" Animals 14, no. 16: 2403. https://doi.org/10.3390/ani14162403