
Dietary Shift in a Barn Owl (Tyto alba) Population Following Partial Abandonment of Cultivated Fields (Central Apennine Hills, Italy)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
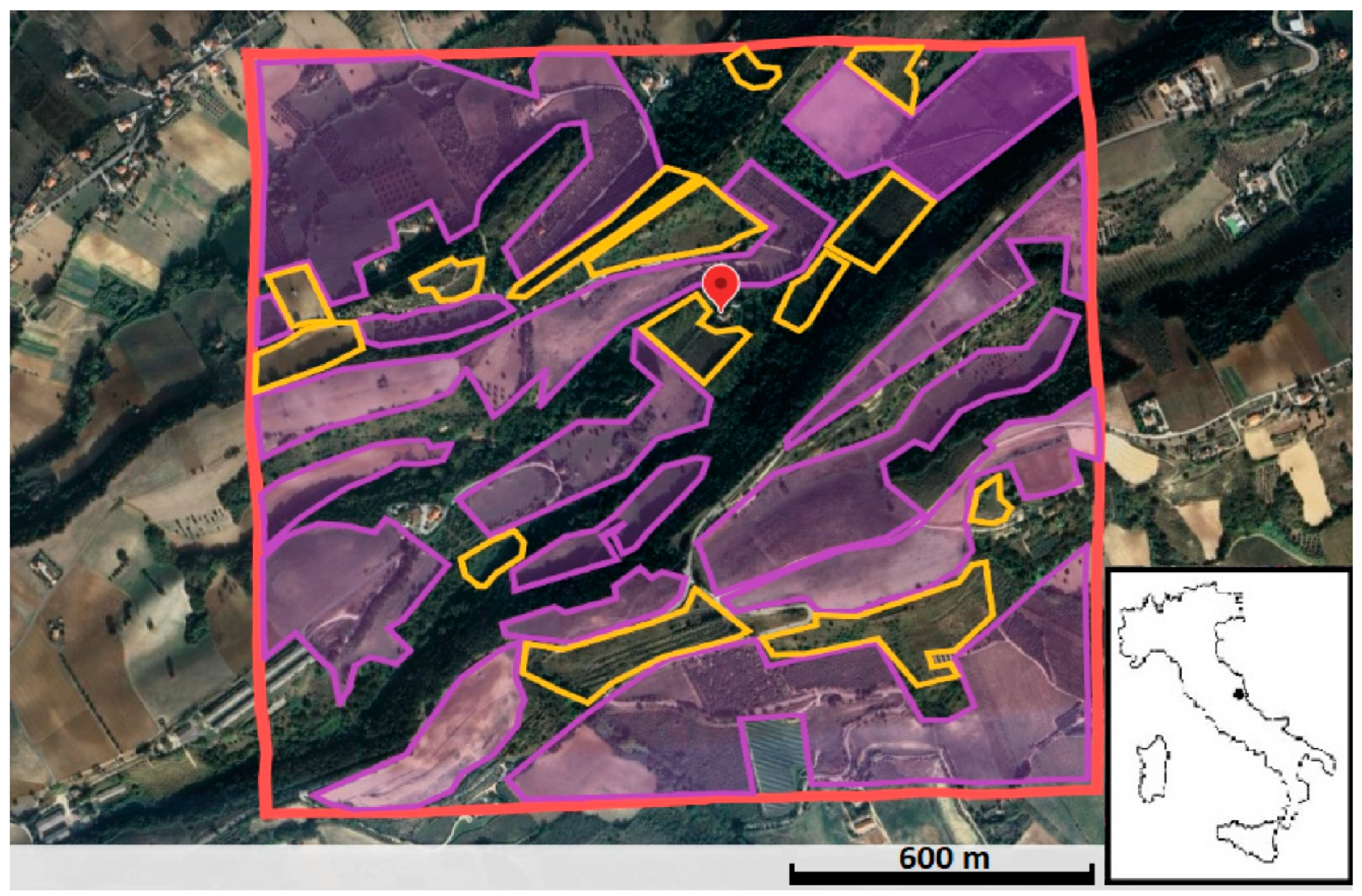
2.1. Study Area
2.2. Data Collection and Transformation
2.3. Numerical Data Analysis
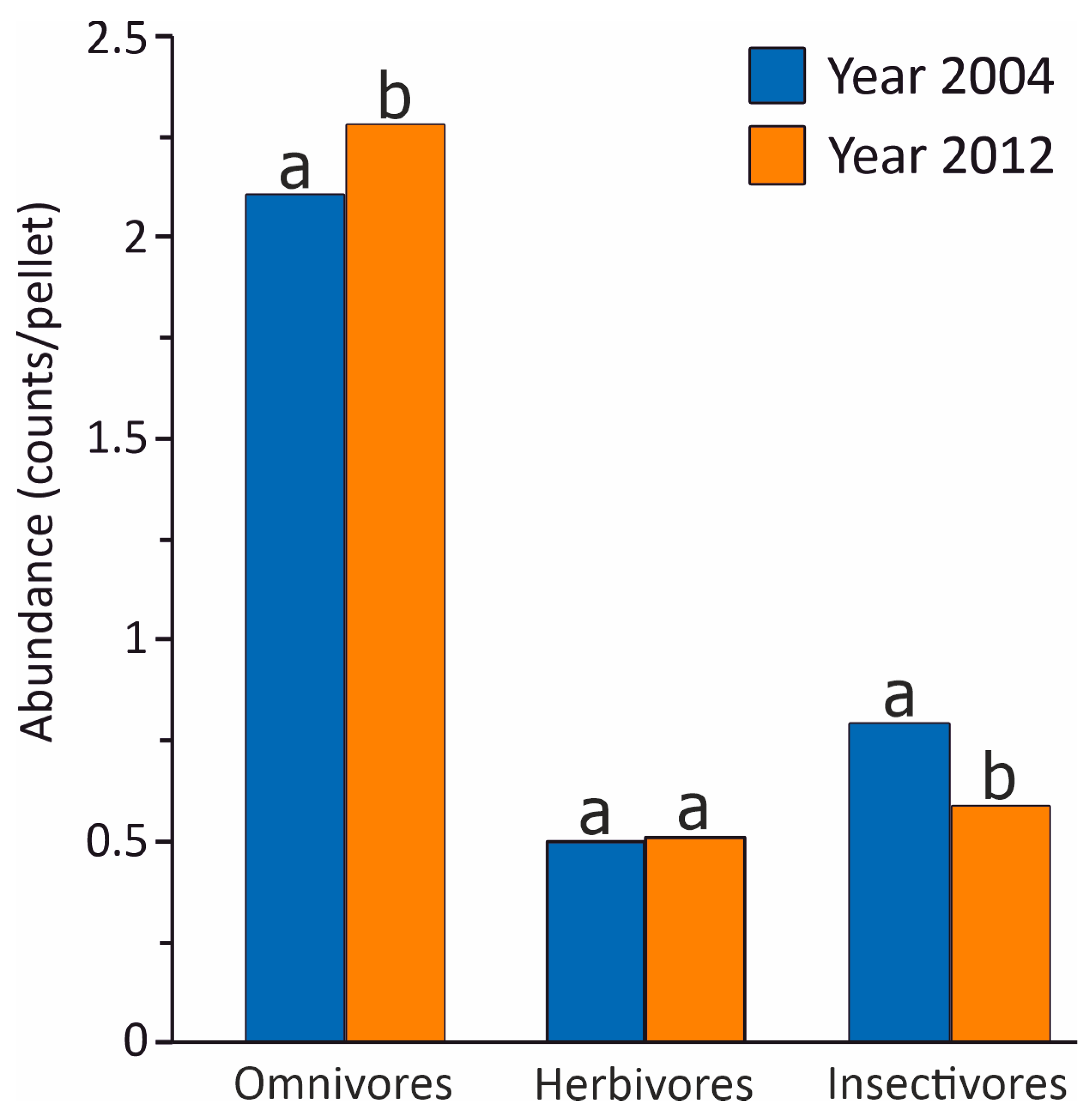
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Debussche, M.; Lepart, J.; Dervieux, A. Mediterranean landscape changes: Evidence from old postcards. Glob. Ecol. Biogeogr. 1999, 8, 3–15. [Google Scholar] [CrossRef]
- Benayas, R.L.M.; Martins, A.; Nicolau, L.M.; Schulz, J. Abandonment of agricultural land: An overview of drivers and consequences. CABI Rev. 2007, 14. [Google Scholar] [CrossRef]
- Benjamin, K.; Domon, G.; Bouchard, A. Vegetation composition and succession of abandoned farmland: Effects of ecological, historical and spatial factors. Landsc. Ecol. 2005, 20, 627–647. [Google Scholar] [CrossRef]
- Fischer, C.; Schröder, B. Predicting spatial and temporal habitat use of rodents in a highly intensive agricultural area. Agric. Ecosyst. Environ. 2014, 189, 145–153. [Google Scholar] [CrossRef]
- Janova, E.; Heroldová, M. Response of small mammals to variable agricultural landscapes in Central Europe. Mamm. Biol. 2016, 81, 488–493. [Google Scholar] [CrossRef]
- Giraudoux, P.; Delattre, P.; Quéré, J.P.; Damange, J.P. Structure and kinetics of rodent populations in a region under agricultural land abandonment. Acta Oecologica 1994, 15, 385–400. [Google Scholar]
- Hansson, L. Dispersal routes of small mammals at an abandoned field in central Sweden. Ecography 1987, 10, 154–159. [Google Scholar] [CrossRef]
- Huntly, N.; Inouye, R.S. Small mammal populations of an old-field chronosequence: Successional patterns and associations with vegetation. J. Mammal. 1987, 68, 739–745. [Google Scholar] [CrossRef]
- Torre, I.; Arrizabalaga, A.; Flaquer, C. Three methods for assessing richness and composition of small mammal communities. J. Mammal. 2004, 85, 524–530. [Google Scholar] [CrossRef]
- Bunn, D.S.; Warburton, A.B.; Wilson, R.D. The Barn Owl; A&C Black: London, UK, 2010. [Google Scholar]
- Roulin, A. Barn Owls: Evolution and Ecology; Cambridge University Press: Cambridge, UK, 2020. [Google Scholar]
- Ševčík, R.; Riegert, J.; Štastný, K.; Zárybnický, J.; Zárybnická, M. The effect of environmental variables on owl distribution in Central Europe: A case study from the Czech Republic. Ecol. Inform. 2021, 64, 101375. [Google Scholar] [CrossRef]
- Séchaud, R.; Schalcher, K.; Machado, A.P.; Almasi, B.; Massa, C.; Safi, K.; Roulin, A. Behaviour-specific habitat selection patterns of breeding barn owls. Mov. Ecol. 2021, 9, 18. [Google Scholar] [CrossRef]
- Salvati, L.; Ranazzi, L.; Manganaro, A. Habitat preferences, breeding success and diet of Barn Owls in Rome: Urban versus rural territories. J. Raptor Res. 2002, 36, 224–228. [Google Scholar]
- Latorre, D.; Merino-Aguirre, R.; Fletcher, D.H.; Cruz, A.; Almeida, D. Effects of habitat structure and feeding habits on productivity and nestling quality of barn owl Tyto alba (Scopoli, 1769) (Strigiformes: Tytonidae) in the Iberian Peninsula. Acta Zool. Bulg. 2022, 74, 203–214. [Google Scholar]
- Herrera, C.M.; Hiraldo, F. Food-niche and trophic relationships among European owls. Ornis Scand. 1976, 7, 29–41. [Google Scholar] [CrossRef]
- Taylor, I. Barn Owls: Predator–Prey Relationships and Conservation; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Lyman, R.L. Rodent-prey content in long-term samples of Barn Owl (Tyto alba) pellets from the northwestern United States reflects local agricultural change. Am. Midl. Nat. 2012, 167, 150–163. [Google Scholar] [CrossRef]
- Torre, I.; Gracia-Quintas, L.; Arrizabalaga, A.; Baucells, J.; Díaz, M. Are recent changes in the terrestrial small mammal communities related to land use change? A test using pellet analyses. Ecol. Res. 2015, 30, 813–819. [Google Scholar] [CrossRef]
- Horváth, A.; Morvai, A.; Horváth, G.F. Food-niche pattern of the Barn Owl (Tyto alba) in intensively cultivated agricultural landscape. Ornis Hung. 2018, 26, 27–40. [Google Scholar] [CrossRef]
- Horváth, A.; Morvai, A.; Horváth, G.F. Difference in small mammal assemblages in the diet of the common barn-owl (Tyto alba) between two landscapes. Acta Zool. Acad. Sci. Hung. 2022, 68, 189–216. [Google Scholar] [CrossRef]
- Paniccia, C.; Carranza, M.L.; Frate, L.; Di Febbraro, M.; Rocchini, D.; Loy, A. Distribution and functional traits of small mammals across the Mediterranean area: Landscape composition and structure definitively matter. Ecol. Indic. 2022, 135, 108550. [Google Scholar] [CrossRef]
- de la Peña, N.M.; Butet, A.; Delettre, Y.; Paillat, G.; Morant, P.; Le Du, L.; Burel, F. Response of the small mammal community to changes in western French agricultural landscapes. Landsc. Ecol. 2003, 18, 265–278. [Google Scholar] [CrossRef]
- Rodríguez, C.; Peris, S. Habitat associations of small mammals in farmed landscapes: Implications for agri-environmental schemes. Anim. Biol. 2007, 57, 301–314. [Google Scholar] [CrossRef]
- Balestrieri, A.; Gazzola, A.; Formenton, G.; Canova, L. Long-term impact of agricultural practices on the diversity of small mammal communities: A case study based on owl pellets. Environ. Monit. Assess. 2019, 191, 725. [Google Scholar] [CrossRef]
- Heisler, L.M.; Somers, C.M.; Poulin, R.G. Owl pellets: A more effective alternative to conventional trapping for broad-scale studies of small mammal communities. Methods Ecol. Evol. 2016, 7, 96–103. [Google Scholar] [CrossRef]
- Milchev, B. Diet of Barn Owl Tyto alba in Central South Bulgaria as influenced by landscape structure. Turk. J. Zool. 2015, 39, 933–940. [Google Scholar] [CrossRef]
- Kross, S.M.; Bourbour, R.P.; Martinico, B.L. Agricultural land use, barn owl diet, and vertebrate pest control implications. Agric. Ecosyst. Environ. 2016, 223, 167–174. [Google Scholar] [CrossRef]
- Massa, C.; Teta, P.; Cueto, G.R. Effects of regional context and landscape composition on diversity and composition of small rodent assemblages in Argentinian temperate grasslands and wetlands. Mammalia 2014, 78, 371–382. [Google Scholar] [CrossRef]
- Hodara, K.; Poggio, S.L. Frogs taste nice when there are few mice: Do dietary shifts in barn owls result from rapid farming intensification? Agric. Ecosyst. Environ. 2016, 230, 42–46. [Google Scholar] [CrossRef]
- Marti, C.D. Dietary trends of Barn Owls in an agricultural ecosystem in northern Utah. Wilson J. Ornithol. 2010, 122, 60–67. [Google Scholar] [CrossRef]
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland biodiversity: Is habitat heterogeneity the key? Trends Ecol. Evol. 2003, 18, 182–188. [Google Scholar] [CrossRef]
- Fischer, J.; Lindenmayer, D.B. Landscape modification and habitat fragmentation: A synthesis. Glob. Ecol. Biogeogr. 2007, 16, 265–280. [Google Scholar] [CrossRef]
- Chaudhary, A.; Pfister, S.; Hellweg, S. Spatially explicit analysis of biodiversity loss due to global agriculture, pasture and forest land use from a producer and consumer perspective. Environ. Sci. Technol. 2016, 50, 3928–3936. [Google Scholar] [CrossRef]
- Labuschagne, L.; Swanepoel, L.H.; Taylor, P.J.; Belmain, S.R.; Keith, M. Are avian predators effective biological control agents for rodent pest management in agricultural systems? Biol. Control 2016, 101, 94–102. [Google Scholar] [CrossRef]
- Montoya, A.; Cabodevilla, X.; Fargallo, J.A.; Biescas, E.; Mentaberre, G.; Villanúa, D. Vertebrate diet of the common kestrel (Falco tinnunculus) and barn owl (Tyto alba) in rain-fed crops: Implications to the pest control programs. Eur. J. Wildl. Res. 2021, 67, 79. [Google Scholar] [CrossRef]
- Frey, C.; Sonnay, C.; Dreiss, A.; Roulin, A. Habitat breeding performance, diet and individual age in Swiss barn owls (Tyto alba). J. Ornithol. 2010, 152, 279–290. [Google Scholar] [CrossRef]
- Brusaferro, A.; Forconi, P.; Fusari, M.; Marini, G.; Renzini, F.; Polini, N.; Mancini, M. L’Atlante degli uccelli nidificanti delle Province di Ascoli Piceno e Fermo. Alula 2009, 16, 674–676. [Google Scholar]
- World Weather & Climate Information. Lapedona Climate by Month. A Year-Round Guide. 2010–2024. Available online: https://weather-and-climate.com/average-monthly-Rainfall-Temperature-Sunshine,lapedona-marche-it,Italy (accessed on 31 July 2024).
- Regione Marche. Carta Geologica Regionale 1:10000; Direzione Paesaggio, Territorio, Urbanistica, Genio Civile: Ancona, Italy, 2020. [Google Scholar]
- Contoli, L. Ruolo dei Micromammiferi nella nicchia trofica del Barbagianni Tyto alba nell’Italia centro-meridionale. Avocetta 1981, 5, 49–64. [Google Scholar]
- Glue, D.E. Food of the barn owl in Britain and Ireland. Bird Study 1974, 21, 200–210. [Google Scholar] [CrossRef]
- Yalden, D.W.; Morris, P.A. The Analysis of Owl Pellets; The Mammal Society: London, UK, 1993. [Google Scholar]
- Amori, G.; Contoli, L.; Nappi, A. Fauna d’Italia. Mammalia II. Erinaceomorpha—Soricomorpha—Lagomorpha—Rodentia; Edizioni Calderini; Comitato Scientifico per la Fauna d’Italia: Bologna, Italy, 2009; Volume 44, 736p. [Google Scholar]
- Brown, R.W.; Lawrence, M.J.; Pope, J. Animals. Tracks, Trails and Signs. Hamlyn Guide; Bounty Books: London, UK, 2004. [Google Scholar]
- Corti, C.; Capula, M.; Luiselli, L.; Razzetti, E.; Sindaco, R. Fauna d’Italia. Reptilia; Edizioni Calderini; Comitato Scientifico per la Fauna d’Italia: Bologna, Italy, 2011; Volume 45, 800p. [Google Scholar]
- Svensson, L.; Mullarney, K.; Zetterström, D. Bird Guide. The Most Complete Guide to the Birds of Britain and Europe; Collins Bird Guide: London, UK, 2022. [Google Scholar]
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Petraitis, P.S. Likelihood measures of niche breadth and overlap. Ecology 1979, 60, 703–710. [Google Scholar] [CrossRef]
- Winemiller, K.O.; Pianka, E.R. Organization in natural assemblages of desert lizards and tropical fishes. Ecol. Monogr. 1990, 60, 27–55. [Google Scholar] [CrossRef]
- Lhotsky, B.; Kovács, B.; Ónodi, G.; Csecserits, A.; Rédei, T.; Lengyel, A.; Kertész, M.; Botta-Dukát, Z. Changes in assembly rules along a stress gradient from open dry grasslands to wetlands. J. Ecol. 2016, 104, 507–517. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; Weedon, J. ‘vegan’ Community Ecology Package (v2.6-6.1). 2024. Available online: https://cran.r-project.org/package=vegan (accessed on 14 June 2024).
- Hothorn, T.; Hornik, K.; van de Wiel, M.A.; Zeileis, A. Implementing a Class of Permutation Tests: The coin Package. J. Stat. Softw. 2008, 28, 1–23. [Google Scholar] [CrossRef]
- Zhang, J. ‘spaa’ Species Association Analysis (v0.2.2). 2022. Available online: https://cran.r-project.org/package=spaa (accessed on 14 June 2024).
- Heroldová, M.; Jánová, E.; Bryja, J.; Tkadlec, E. Set-aside plots-source of small mammal pests? Folia Zool. 2005, 54, 337. [Google Scholar]
- Torre, I.; Requejo, A.; Arrizabalaga, A.; Baucells, J. Disentangling the effects of climate and land uses on small mammals in agroecosystems of NE Spain. Diversity 2024, 16, 343. [Google Scholar] [CrossRef]
- Benedek, A.M.; Sîrbu, I. Responses of small mammal communities to environment and agriculture in a rural mosaic landscape. Mamm. Biol. 2018, 90, 55–65. [Google Scholar] [CrossRef]
- Milana, G.; Luiselli, L.; Amori, G. Forty years of dietary studies on barn owl (Tyto alba) reveal long term trends in diversity metrics of small mammal prey. Anim. Biol. 2018, 68, 129–146. [Google Scholar] [CrossRef]
- Torre, I.; Díaz, M. Assessing the effects of landscape change on the occupancy dynamics of the greater white-toothed shrew Crocidura russula. Life 2022, 12, 1230. [Google Scholar] [CrossRef]
- Horváth, G.F.; Mánfai, K.; Horváth, A. Relationship between landscape structure and the diet of Common Barn-owl at different distances from the Drava River ecological corridor. Ornis Hung. 2023, 31, 88–110. [Google Scholar] [CrossRef]
- Bosé, M.; Guidali, F. Seasonal and geographic differences in the diet of the Barn Owl in an agroecosystem in northern Italy. J. Raptor Res. 2001, 35, 240–246. [Google Scholar]
- Heroldová, M.; Bryja, J.; Zejda, J.; Tkadlec, E. Structure and diversity of small mammal communities in agriculture landscape. Agric. Ecosyst. Environ. 2007, 120, 206–210. [Google Scholar] [CrossRef]
- Veselovský, T.; Bacsa, K.; Tulis, F. Barn Owl (Tyto alba) diet composition on intensively used agricultural land in the Danube Lowland. Acta Univ. Agric. Et Silvic. Mendel. Brun. 2017, 65, 225–233. [Google Scholar] [CrossRef]
- Arlettaz, R.; Krähenbühl, M.; Almasi, B.; Roulin, A.; Schaub, M. Wildflower areas within revitalized agricultural matrices boost small mammal populations but not breeding Barn Owls. J. Ornithol. 2010, 151, 553–564. [Google Scholar] [CrossRef]
- Bühler, R.; Schalcher, K.; Séchaud, R.; Michler, S.; Apolloni, N.; Roulin, A.; Almasi, B. Influence of prey availability on habitat selection during the non-breeding period in a resident bird of prey. Mov. Ecol. 2023, 11, 14. [Google Scholar] [CrossRef]
- Embar, K.; Mukherjee, S.; Kotler, B.P. What do predators really want? The role of gerbil energetic state in determining prey choice by Barn Owls. Ecology 2014, 95, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Bernard, N.; Michelat, D.; Raoul, F.; Quere, J.-P.; Delattre, P.; Giraudoux, P. Dietary response of Barn Owls (Tyto alba) to large variations in populations of common voles (Microtus arvalis) and European water voles (Arvicola terrestris). Can. J. Zool. 2010, 88, 416–426. [Google Scholar] [CrossRef]
- Andrade, A.; de Menezes, J.F.S.; Monjeau, A. Are owl pellets good estimators of prey abundance? J. King Saud Univ.-Sci. 2016, 28, 239–244. [Google Scholar] [CrossRef]
- Pelosi, I.; Dodaro, G.; Battisti, C.; Scalici, M. Crop-land simplification impacts differently on small mammal communities: Evidence from diversity/dominance plots using Barn Owl (Tyto alba) pellets. Community Ecol. 2024, 25, 1–9. [Google Scholar] [CrossRef]
- Roulin, A. Shrews and moles are less often captured by European Barn Owls Tyto alba nowadays than 150 years ago. Bird Study 2016, 63, 559–563. [Google Scholar] [CrossRef]
- Gentili, S.; Sigura, M.; Bonesi, L. Decreased small mammals species diversity and increased population abundance along a gradient of agricultural intensification. Hystrix 2004, 25, 39–44. [Google Scholar] [CrossRef]
- Moysi, M.; Christou, M.; Goutner, V.; Kassinis, N.; Iezekiel, S. Spatial and temporal patterns in the diet of barn owl (Tyto alba) in Cyprus. J. Biol. Res.-Thessalon. 2018, 25, 9. [Google Scholar] [CrossRef]
- Battisti, C.; Dodaro, G.; di Bagno, E.; Amori, G. Small mammal assemblages in land-reclaimed areas: Do historical soil use changes and recent anthropisation affect their dominance structure? Ethol. Ecol. Evol. 2020, 32, 282–288. [Google Scholar] [CrossRef]
- Serafini, V.N.; Priotto, J.W.; Gomez, M.D. Effects of agroecosystem landscape complexity on small mammals: A multi-species approach at different spatial scales. Landsc. Ecol. 2019, 34, 1117–1129. [Google Scholar] [CrossRef]
- Martin, J.M.; Branch, L.C.; Raid, R.N.; Beyeler, S.C. Temporal instability of agricultural habitat reduces reproductive success of Barn Owls (Tyto alba). Ornithology 2010, 127, 909–916. [Google Scholar] [CrossRef]
- Askew, N.P.; Searle, J.B.; Moore, N.P. Agri-environment schemes and foraging of barn owls Tyto alba. Agric. Ecosyst. Environ. 2007, 118, 109–114. [Google Scholar] [CrossRef]
- Martínez, J.A.; Zuberogoitia, I. Habitat preferences and causes of population decline for Barn Owls Tyto alba: A multi-scale approach. Ardeola 2004, 51, 303–317. [Google Scholar]
- Toms, M.P.; Crick, H.Q.P.; Shawyer, C.R. The status of breeding barn owls (Tyto alba) in the UK 1995–1997. Bird Study 2001, 48, 23–37. [Google Scholar] [CrossRef]
- Charter, M.; Izhaki, I.; Meyrom, K.; Motro, Y.; Leshem, Y. Diets of barn owls differ in the same agricultural region. Wilson J. Ornithol. 2009, 121, 378–383. [Google Scholar] [CrossRef]
- Contoli, L. Sistemi trofici e corologia: Dati su Soricidae, Talpidae ed Arvicolidae d’Italia predati da Tyto alba (Scopoli 1769). Hystrix 1986, 1, 95–118. [Google Scholar]
- Purger, J.J.; Szép, D. An attempt to determine the size of the Common Barn-owl’s (Tyto alba) hunting area based on its prey composition. Avian Biol. Res. 2022, 15, 41–46. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Area (ha) | ||
|---|---|---|
| Survey Years | 2004 | 2012 |
| Cultivated fields | 128.78 | 107.92 |
| Abandoned fields | 0 | 20.86 |
| Scrub | 41.97 | 41.97 |
| Woods | 52.01 | 52.01 |
| Human built-up areas | 10.30 | 10.30 |
| Dietary Feature | Observed Difference | Simulated Mean Difference | Standardised Effect Size | p |
|---|---|---|---|---|
| Prey taxon evenness | −0.06935 | −0.00163 | −3.891 | 0.0001 |
| Food niche overlap | 0.98656 | 0.99523 | −1.996 | 0.0459 |
| Omnivorous prey | 0.17206 | −0.01143 | 2.228 | 0.0259 |
| Herbivorous prey | 0.01224 | −0.01177 | 0.295 | 0.7683 |
| Insectivorous prey | −0.20762 | −0.01130 | −2.354 | 0.0186 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Achille, G.; Gafta, D.; Szabó, C.; Canzian, F.; Polini, N. Dietary Shift in a Barn Owl (Tyto alba) Population Following Partial Abandonment of Cultivated Fields (Central Apennine Hills, Italy). Animals 2024, 14, 2562. https://doi.org/10.3390/ani14172562
Achille G, Gafta D, Szabó C, Canzian F, Polini N. Dietary Shift in a Barn Owl (Tyto alba) Population Following Partial Abandonment of Cultivated Fields (Central Apennine Hills, Italy). Animals. 2024; 14(17):2562. https://doi.org/10.3390/ani14172562
Chicago/Turabian StyleAchille, Gabriele, Dan Gafta, Csaba Szabó, Fadia Canzian, and Nazzareno Polini. 2024. "Dietary Shift in a Barn Owl (Tyto alba) Population Following Partial Abandonment of Cultivated Fields (Central Apennine Hills, Italy)" Animals 14, no. 17: 2562. https://doi.org/10.3390/ani14172562