
A Whole Range of Cattle—An Isotopic Perspective on Roman Animal Husbandry in Lower Austria and Burgenland (Austria)


Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Assessing Size in the Skeletal Remains of Roman Cattle
1.2. Stable Isotope Ratios to Investigate Cattle Diet and Origin
2. Materials and Methods
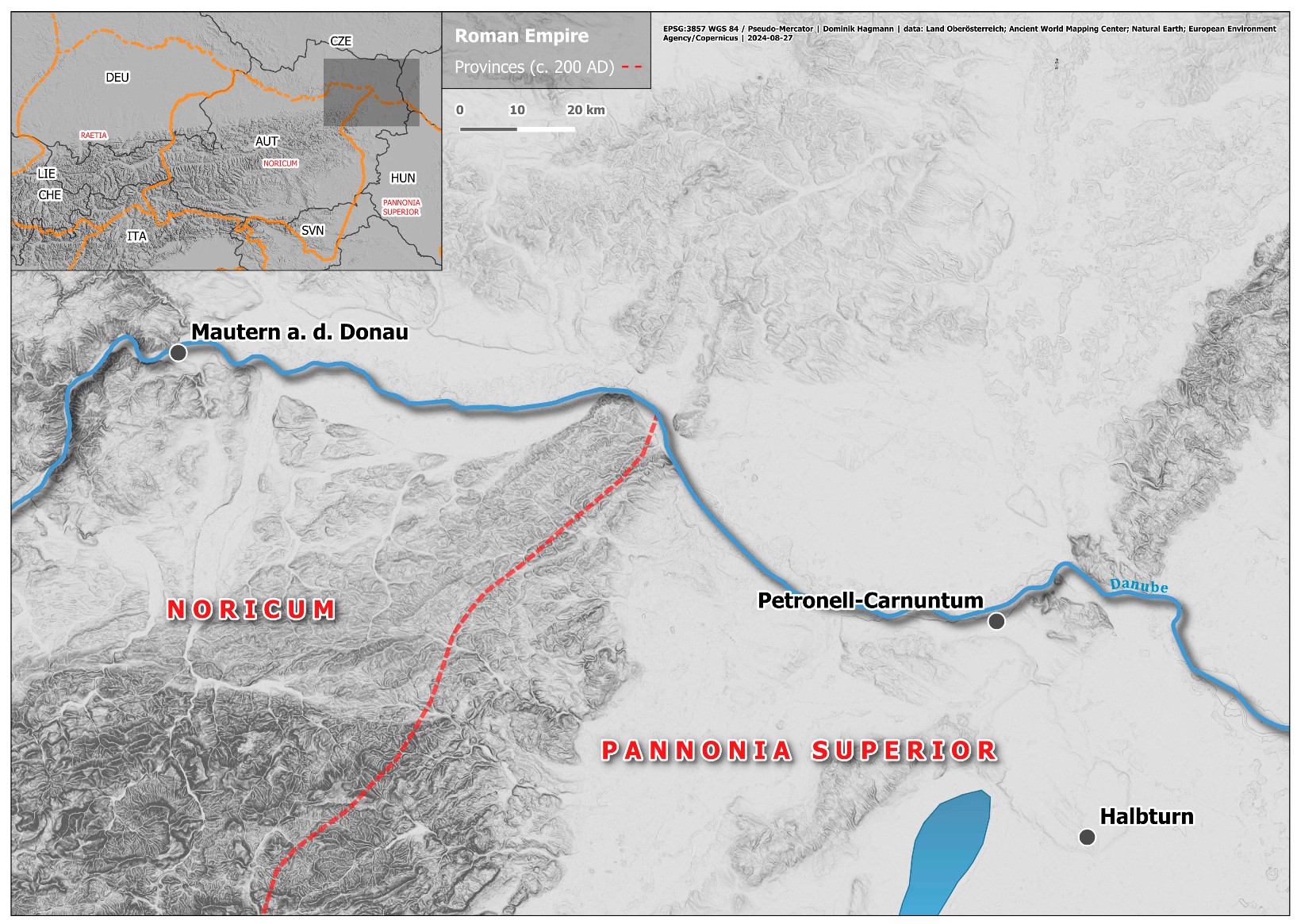
2.1. Material—Cattle Bones Used in This Study
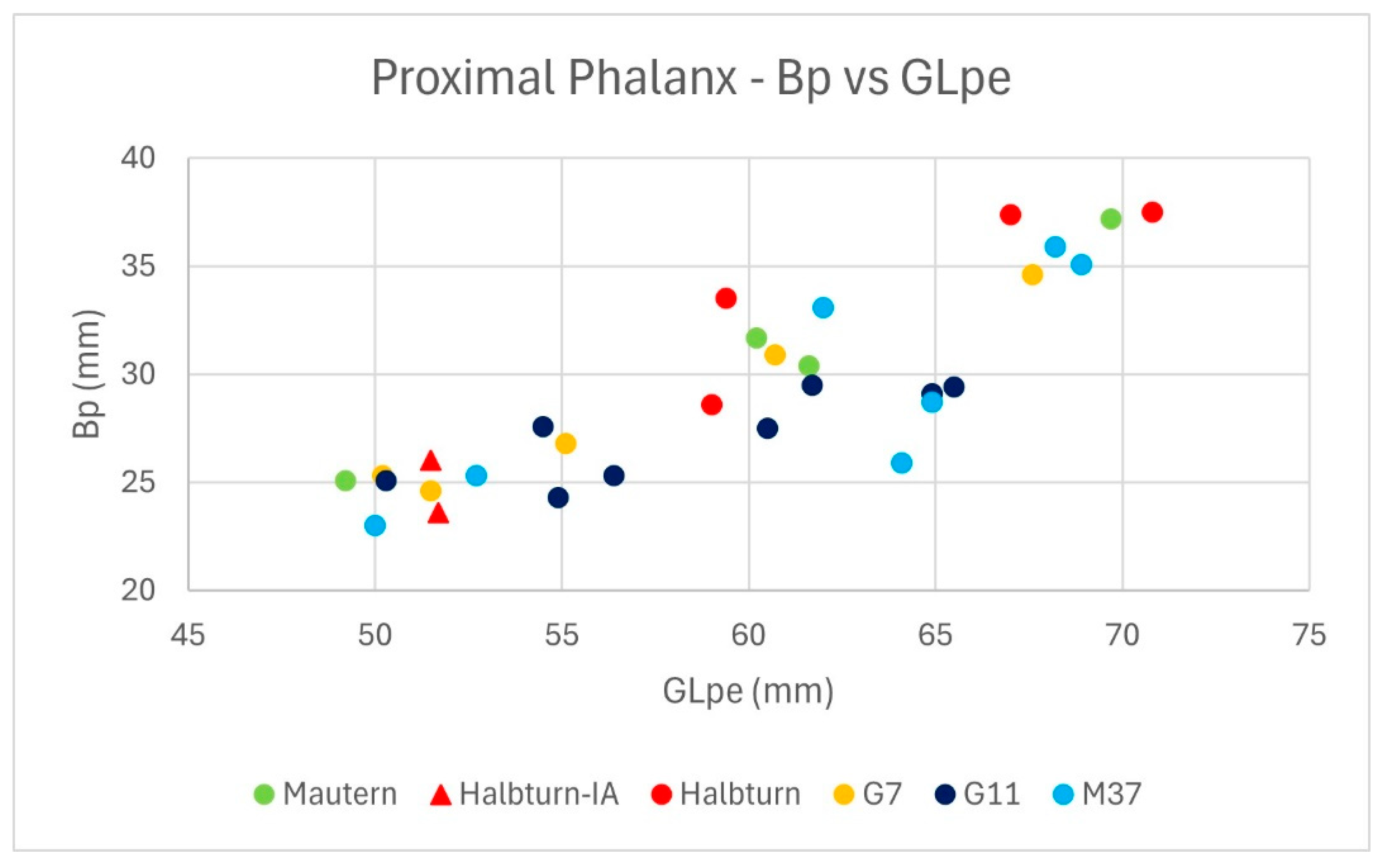
2.2. Osteometric Variability in the Cattle Sample
2.3. Stable Isotopes—Methods
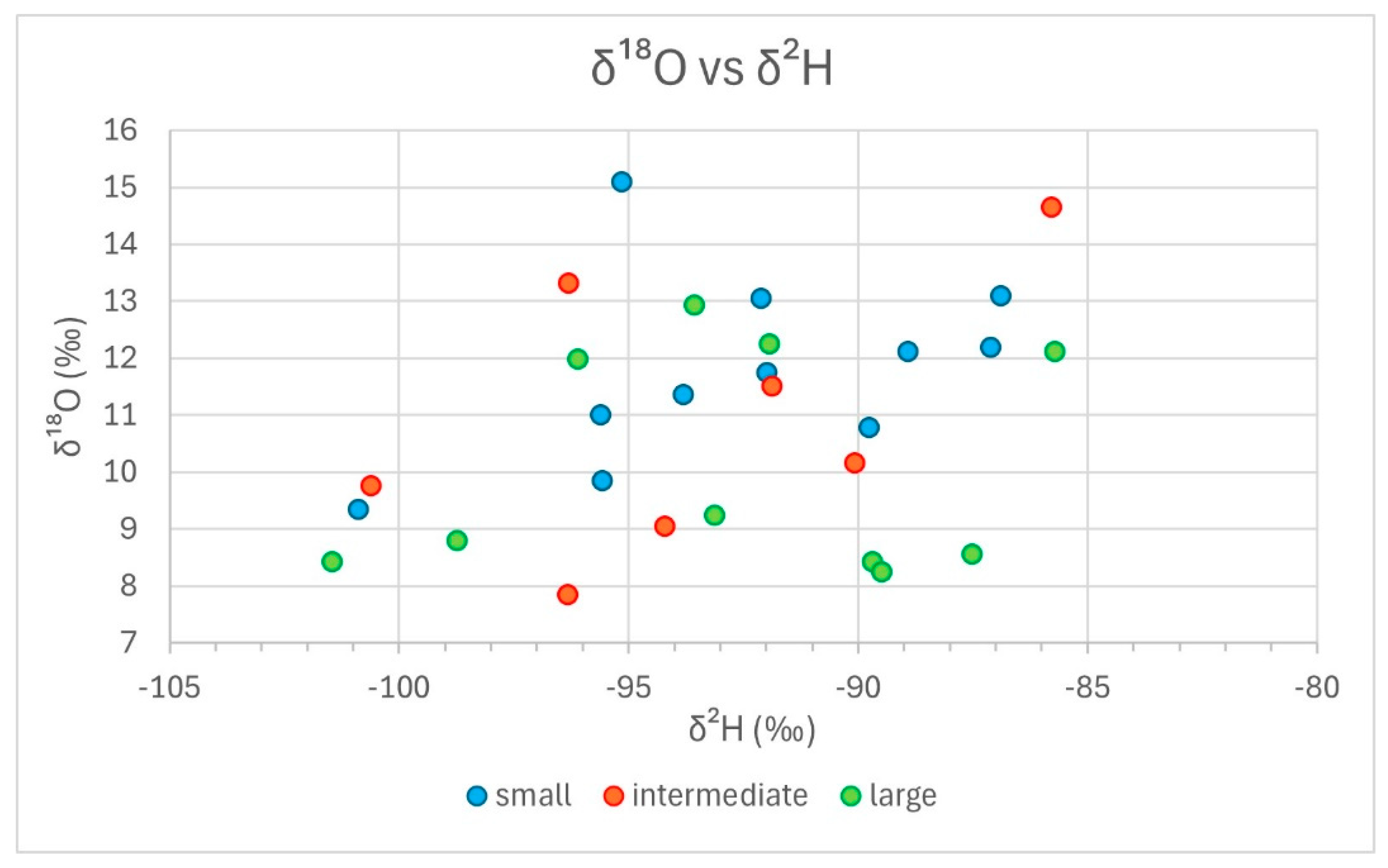
3. Results from Stable Isotope Analyses
4. Discussion
- -
- There is no consistent relationship between modal size and stable isotope composition measured in Roman cattle bones;
- -
- The observed variability is explained by inter-site, or inter-context, differences rather than by size dependence.
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amschler, J.W. Ur-und frühgeschichtliche Haustierfunde aus Österreich. Archaeol. Austriaca 1949, 3, 100. [Google Scholar]
- Bökönyi, S. History of Domestic Mammals in Central and Eastern Europe; Akad. Kiadó, Reprint: Budapest, Hungary, 1988. [Google Scholar]
- Benecke, N. Archäozoologische Studien zur Entwicklung der Haustierhaltung in Mitteleuropa und Südskandinavien von den Anfängen bis zum Ausgehenden Mittelalter. Schriften zur Ur-und Frühgeschichte; Akademie Verlag: Berlin, Germany, 1994; Schriften zur Ur-und Frühgeschichte 46. [Google Scholar]
- Peters, J. Römische Tierhaltung und Tierzucht: Eine Synthese aus Archäozoologischer Untersuchung und Schriftlich-Bildlicher Überlieferung. Passauer Universitätsschriften zur Archäologie 5; Leidorf: Rahden, Germany, 1998. [Google Scholar]
- Valenzuela-Lamas, S.; Albarella, U. Animal Husbandry across the Western Roman Empire: Changes and Continuities. Eur. J. Archaeol. 2017, 20, 402–415. [Google Scholar] [CrossRef]
- Colominas, L. Arqueozoología y Romanización: Producción, Distribución y Consumo de Animales en el Nordeste de la Península Ibérica Entre Los Siglos V ane-V dne; BAR International Series 2480; Archaeopress: Oxford, UK, 2013. [Google Scholar]
- Colominas, L.; Schlumbaum, A.; Saña, M. The impact of the Roman Empire on animal husbandry prac-tices: Study of the changes in cattle morphology in the north-east of the Iberian Peninsula through osteometric and ancient DNA analyses. Archaeol. Anthropol. Sci. 2014, 6, 1–16. [Google Scholar] [CrossRef]
- Trentacoste, A.; Nieto-Espinet, A.; Guimarães, S.; Wilkens, B.; Petrucci, G.; Valenzuela-Lamas, S. New trajectories or accelerating change? Zooarchaeological evidence for Roman transformation of animal husbandry in Northern Italy. Archaeol Anthr. Sci. 2021, 13, 25. [Google Scholar] [CrossRef] [PubMed]
- Seetah, K. Humans, Animals, and the Craft of Slaughter in Archaeo-Historic Societies; Cambridge University Press: Cambridge, UK, 2019. [Google Scholar]
- Lepetz, S. L’animal dans la société gallo-romaine de la France du Nord, Revue archéologique de Picardie. Numéro Spécial 1996, 12, 1–174. [Google Scholar]
- Groot, M.; Albarella, U. Cattle husbandry in the Iron Age and Roman Netherlands: Chronological developments and regional differences in cattle frequencies, management, size and shape. Praehist. Z. 2022, 99, 1–54. [Google Scholar] [CrossRef]
- Albarella, U.; Johnstone, C.; Vickers, K. The development of animal husbandry from the Late Iron Age to the end of the Roman period: A case study from South-East Britain. J. Archaeol. Sci. 2008, 35, 1828–1848. [Google Scholar] [CrossRef]
- Pigière, F. The Evolution of Cattle Husbandry Practices in the Roman Period in Gallia Belgica and Western Germania Inferior. Eur. J. Archaeol. 2017, 20, 472–493. [Google Scholar] [CrossRef]
- Boessneck, J. Die Tierknochenfunde aus dem Oppidum von Manching. Die Ausgrabungen in Manching; Steiner: Wiesbaden, Germany, 1971; Volume 6. [Google Scholar]
- Saliari, K.; Amory, C.; Draganits, E.; Ramsl, P.C.; Tobias, B.; Pucher, E.; Parson, W. Morphometric and genetic evidence for cattle imports from the Mediterranean into present-day Austria during the Iron Age. J. Archaeol. Sci. Rep. 2023, 48, 103842. [Google Scholar] [CrossRef]
- Groot, M.; Deschler-Erb, S. Think global, act local: Regionalism and the supply of meat to the Roman army. In The Role of Zooarchaeology in the Study of the Western Roman Empire; Journal of Roman Archaeology Supplementary Series 107; Allen, M.G., Ed.; The Society of the Promotion of Roman Studies: Portsmouth, RI, USA, 2019; pp. 53–72. [Google Scholar]
- Breuer, G.; Rehazek, A.A.; Stopp, B. Grössenveränderungen des Hausrindes: Osteometrische Untersuchungen grosser Fundserien aus der Nordschweiz von der Spätlatènezeit bis ins Frühmittelalter am Beispiel von Basel, Augst (Augusta Raurica) und Schleitheim-Brüel. Jahresber Augst Kaiseraugst 1999, 20, 207–228. [Google Scholar]
- Pucher, E. Der Tierknochenfundkomplex eines germanischen Dorfs im römischen Machtbereich: Bruckneudorf, Fundberichte aus Österreich. Fundberichte Osterr. 2016, 55, D235–D422. [Google Scholar]
- Trixl, S.; Steidl, B.; Peters, J. Archaeology and Zooarchaeology of the Late Iron Age-Roman Transition in the Prov-ince of Raetia (100 BC–100 AD). Eur. J. Archaeol. 2017, 20, 431–450. [Google Scholar] [CrossRef]
- Groot, M. Developments in Animal Husbandry and Food Supply in Roman Germania Inferior. Eur. J. Archaeol. 2017, 20, 451–471. [Google Scholar] [CrossRef]
- Duval, C.; Lepetz, S.; Horard-Herbin, M.-P. Diversité des cheptels et diversification des morphotypes bovins dans le tiers nord-ouest des Gaules entre la fin de l’âge du Fer et la période romaine. Gallia 2012, 69, 79–114. [Google Scholar]
- Madgwick, R.; Lewis, J.; Grimes, V.; Guest, P. On the hoof: Exploring the supply of animals to the Roman legionary fortress at Caerleon using strontium (87Sr/86Sr) isotope analysis. Archaeol. Anthropol. Sci. 2019, 11, 223–235. [Google Scholar] [CrossRef]
- Groot, M.; Evans, J.; Albarella, U. Mobility of cattle in the Iron Age and Roman Netherlands. J. Archaeol. Sci. Rep. 2020, 32, 102416. [Google Scholar] [CrossRef]
- von den Driesch, A. A Guide to the Measurement of Animal Bone from Archaeological Sites; Peabody Museum Press: Cambridge, MA, USA, 1976. [Google Scholar]
- Kunst, G.K.; Gál, E. The Harvest is great, but the Workers are few: Cattle Measurements from Carnuntum and beyond. In Magnis Itineribus: Festschrift für Verena Gassner zum 65; Ployer, R., Svoboda-Baas, D., Eds.; Phoibos Verlag: Geburtstag, Austria, 2021; pp. 163–177. [Google Scholar]
- Marom, N.; Bar-Oz, G. Zooarchaeology and Social Identity in Bronze and Iron Ages Israel: A Research Framework. In Hamilton-Dyer Archaeozoology of the Near East X Proceedings of the Tenth Inter-National Symposium on the Archaeozoology of South-Western Asia and Adjacent Areas; De Cupere, B., Linseele, V.S., Eds.; Belgium Peeters Publishers: Brussels, Belgium, 2013; pp. 222–247. [Google Scholar]
- Baas, H. Die Tierknochenfunde aus den Spätrömischen Siedlungsschichten von Lauriacum. 1. Die Rinderknochen. Diss. med. vet. 1966, LMU-München. Ph.D. Thesis, LMU Munich University, Munich, Germany, 1966. [Google Scholar]
- Riedel, A. Die Tierknochenfunde des römerzeitlichen Lagervicus von Traismauer/Augustiana in Niederösterreich. Ann. Naturhistor. Mus. Wien 1993, 95, 179–294. [Google Scholar]
- Krueger, H.W.; Sullivan, C.H. Models for carbon isotope fractionation between diet and bone. Stable Isot. Nutr. ACS Symp. Ser. 1984, 258, 205–220. [Google Scholar]
- Lee-Thorp, J.A.; Sealy, J.C.; van der Merwe, N.J. Stable carbon isotope ratio differences between bone collagen and apatite, and their relationship to diet. J. Archaeol. Sci. 1989, 16, 585–599. [Google Scholar] [CrossRef]
- Schoeninger, M.J.; De Niro, M.J.; DeNiro, M.J. Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochim. Cosmochim. Acta. 1984, 48, 625–639. [Google Scholar] [CrossRef]
- Bogaard, A.; Heaton, T.H.E.; Poulton, P.; Merbach, I. The impact of manuring on nitrogen isotope ratios in cereals: Archaeological implications for reconstruction of diet and crop management practices. J. Archaeol. Sci. 2007, 34, 335–343. [Google Scholar] [CrossRef]
- Joseph, M.C.; Brookshire, E.N.J.; Michael, D.C.; Niles, J.H.; Keisuke, K.; Erika, M.-S.; Lixin, W. Ecological interpretations of nitrogen isotope ratios of terrestrial plants and soils. Plant Soil 2015, 396, 1–26. [Google Scholar]
- Drucker, D.G.; Bridault, A.; Cupillard, C.; Hujic, A.; Bocherens, H. Evolution of habitat and environment of red deer (Cervus elaphus) during the Late-glacial and early Holocene in eastern France (French Jura and the western Alps) using multi-isotope analysis (δ 13C, δ 15N, δ 18O, δ 34S) of archaeological remains. Quat. Int. 2011, 245, 268–278. [Google Scholar] [CrossRef]
- Gillis, R.; Zanon, M. Forest pasturing in Central and Northern Europe: Stable carbon isotope evidence and perspectives. In Biodiversités, Environnements et Sociétés Depuis la Préhistoire: Nouveaux Marqueurs et Approches Intégrées; Nicoud, E., Balasse, M., Desclaux, E., Théry-Parisot, I., Eds.; APDCA-CEPAM: Nice Côte d’Azur, French, 2021; pp. 117–126. [Google Scholar]
- Horacek, M.; Frischauf, C.; Pacher, M.; Rabeder, G. Stable isotopic analyses of cave bear bones from the Conturines cave (2,800 m, South Tyrol, Italy). Braunschweiger Naturkundliche Schriften 2012, 11, 47–52. [Google Scholar]
- Horacek, M.; Min, J.-S. Discrimination of Korean beef from beef of other origin by stable isotope measure-ments. Food Chem. 2010, 121, 517–520. [Google Scholar] [CrossRef]
- Bontempo, L.; Perini, M.; Pianezze, S.; Horacek, M.; Roßmann, A.; Kelly, S.; Thomas, F.; Heinrich, K.; Schlicht, C.; Schellenberg, A.; et al. Characterisation of Beef Coming from Dif-ferent European Countries through Stable Isotope (H, C, N and S) Ratio Analysis. Molecules 2023, 28, 2856. [Google Scholar] [CrossRef]
- Horacek, M.; Jakusch, M.; Krehan, H. Control of origin of larch wood: Discrimination between European (Austrian) and Siberian origin. Rapid Commun. Mass Spectrom. 2009, 23, 3688–3692. [Google Scholar] [CrossRef]
- Horacek, M.; Hansel-Hohl, K.; Burg, K.; Soja, G.; Okello-Anyanga, W.; Fluch, S. Control of origin of sesame oil from various countries by stable isotope analysis and DNA based markers—A pilot study. PLoS ONE 2015, 10, e0123020. [Google Scholar] [CrossRef]
- Pederzani, S.; Britton, K. Oxygen isotopes in bioarchaeology; principles and applications, challenges and opportuni-ties. Earth-Sci. Rev. 2019, 188, 77–107. [Google Scholar] [CrossRef]
- Makarewicz, C.A.; Sealy, J. Dietary reconstruction, mobility, and the analysis of ancient skeletal tissues: Expanding the prospects of stable isotope research in archaeology. J. Archaeol. Sci. 2015, 56, 146–158. [Google Scholar] [CrossRef]
- Bartosiewicz, L. The anatomical position and metric traits of phalanges in cattle. Rev. De Paléobiologie 1993, 12, 21–43. [Google Scholar]
- Kunst, G.K.; Gál, E.; Gassner, V. 2021, Choice beef for the worshippers—The cattle record from the sanctuary of Jupiter Heliopolitanus at Carnuntum (Austria). In Roman Animals in Ritual and Funerary Contexts, Proceedings of the 2nd Meeting of the Zooarchaeology of the Roman Period Working Group; Desch-ler-Erb, S., Albarella, U., Valenzuela Lamas, S., Rasbach, G., Eds.; Harassowitz: Wiesbaden, Germany, 2018; Volume 26, pp. 123–140. [Google Scholar]
- Kunst, G.K.; Bodeneingriffe, B.M.U. Tierreste aus dem römischen Gräberfeld Halbturn. In Das Kaiserzeitliche Gräberfeld von Halbturn, Burgenland: Teil 2: Intention, Abfall oder Zufall: Naturwissenschaftliche Untersuchungen; Doneus, N., Ed.; Monographien des Römisch-Germanischen Zentralmuseums: Mainz, Germany, 2014; Volume 122, pp. 485–640. [Google Scholar]
- Pieler, F.; von Schmitsberger, O. Rettungsgrabungen im Bereich des westlichen Vicus von Favianis in Mautern, Melkerstraße. Fundberichte Aus Osterr. 2004, 43, 794–806. [Google Scholar]
- Kunst, G.K. Die feine Linie—Hinweise auf Rinderschinken im römischen Niederösterreich. In Beiträge zum Tag der Niederösterreichischen Landesarchäologie 2017: Festschrift für Ernst Lauermann; Pieler, F., Trebsche, P., Eds.; Katalog des Niederösterreichischen Landesmuseums: Asparn/Zaya, Austria, 2017; Volume 541, pp. 316–326. [Google Scholar]
- Bocherens, H.; Sandrock, O.; Kullmer, O.; Schrenk, F. Hominin palaeoecology in Late Pliocene Malawi: First insights from isotopes (13C, 18O) in mammal teeth. S. Afr. J. Sci. 2011, 107, 1–6. [Google Scholar] [CrossRef]
- Thanheiser, U.; Wiesinger, S. Abfall, Zufall oder Intention? Die Pflanzenreste aus dem Gräberfeld Halbturn. In Das kaiserzeitliche Gräberfeld von Halbturn, Burgenland: Teil 2: Intention, Abfall oder Zufall: Naturwissenschaftliche Untersuchungen; Doneus, N., Ed.; Monographien des Römisch-Germanischen Zentralmuseums: Mainz, Germany, 2014; Volume 122, pp. 641–664. [Google Scholar]
- Blanz, M.; Mainland, I.; Richards, M.; Balasse, M.; Ascough, P.; Wolfhagen, J.; Taggart, M.A.; Feldmann, J. Identifying seaweed consumption by sheep using isotope analysis of their bones and teeth: Modern reference δ13C and δ15N values and their archaeological implications. J. Archaeol. Sci. 2020, 118, 105140. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lab.nr. | Site Context | Glpe (mm) | Bp (mm) | SD (mm) | Bd (mm) | Size Group | 13C (‰) | 15N (‰) | 2H (‰) | 18O (‰) |
|---|---|---|---|---|---|---|---|---|---|---|
| 17-0071 | Mautern | 69.7 | 37.2 | 29.5 | 33.8 | large | −21.0 | 5.8 | −93.1 | 9.2 |
| 17-0072 | Mautern | 49.2 | 25.1 | 21.2 | 23.1 | small | −21.6 | 5.8 | −95.6 | 9.8 |
| 17-0073 | Mautern | 60.2 | 31.7 | 28.6 | 30.5 | intermediate | −20.7 | 5.9 | −94.2 | 9.1 |
| 17-0074 | Mautern | 61.6 | 30.4 | 24.7 | 28.2 | intermediate | −21.1 | 5.9 | −100.6 | 9.8 |
| 17-0075 | Halbturn-IA | 51.7 | 23.6 | 19.6 | 23.1 | small | −20.3 | 11.1 | −100.9 | 9.3 |
| 17-0076 | Halbturn-IA | 51.5 | 26 | small | −21.2 | 7.1 | ||||
| 17-0077 | Halbturn | 59.4 | 33.5 | 26.8 | 31.7 | intermediate | −19.6 | 8.5 | −90.1 | 10.2 |
| 17-0078 | Halbturn | 59 | 28.6 | 21.9 | 28.3 | intermediate | −20.3 | 7.4 | −85.8 | 14.7 |
| 17-0079 | Halbturn | 70.8 | 37.5 | 30.9 | 35.6 | large | −20.5 | 7.9 | −91.9 | 12.2 |
| 17-0080 | Halbturn | 67 | 37.4 | 32.5 | 35.3 | large | −20.3 | 10.1 | −93.6 | 12.9 |
| CA1 | G11 | 54.5 | 27.6 | 23.5 | 27.7 | small | −22.2 | 5.5 | −93.8 | 11.4 |
| CA2 | G11 | 64.9 | 29.1 | 24.5 | 29.1 | large | −23.0 | 7.4 | −101.5 | 8.4 |
| CA3 | G7 | 67.6 | 34.6 | 29 | 34.8 | large | −20.4 | 8.5 | −85.7 | 12.1 |
| CA4 | G11 | 50.3 | 25.1 | 21.8 | 23.6 | small | −21.8 | 6.0 | −88.9 | 12.1 |
| CA5 | G11 | 61.7 | 29.5 | 27.5 | 30.5 | intermediate | −22.1 | |||
| CA6 | G11 | 65.5 | 29.4 | 23.7 | 28.2 | large | −22.3 | 7.9 | −96.1 | 12.0 |
| CA7 | G11 | 56.4 | 25.3 | 22.2 | 24.7 | small | −23.3 | 5.8 | −95.6 | 11.0 |
| CA8 | G11 | 60.5 | 27.5 | 22.1 | 26.6 | intermediate | −21.0 | 6.6 | −96.3 | 13.3 |
| CA9 | G11 | 54.9 | 24.3 | 21.2 | 22.7 | small | −23.7 | 7.1 | −95.1 | 15.1 |
| CA10 | G7 | 50.2 | 25.3 | 20.9 | 25.2 | small | −21.8 | 8.0 | −86.9 | 13.1 |
| G7 | 55.1 | 26.8 | 22 | 25.6 | small | −21.8 | 4.8 | −89.8 | 10.8 | |
| G7 | 51.5 | 24.6 | 20.6 | 23.3 | small | −20.8 | 6.0 | −92.0 | 11.8 | |
| G7 | 60.7 | 30.9 | 27.3 | 27.1 | intermediate | −21.4 | 6.5 | −91.9 | 11.5 | |
| M37 | 50 | 23 | 19.2 | 22.4 | small | −21.6 | 3.9 | −92.1 | 13.1 | |
| M37 | 52.7 | 25.3 | 22.9 | 24.6 | small | −19.5 | 6.5 | −87.1 | 12.2 | |
| M37 | 62 | 33.1 | 31.2 | 33.2 | intermediate | −20.7 | 6.1 | −96.3 | 7.9 | |
| M37 | 64.1 | 25.9 | 21.5 | 26 | large | −19.9 | 6.2 | −89.7 | 8.4 | |
| M37 | 68.2 | 35.9 | 30.7 | 38.2 | large | −22.0 | 5.8 | −87.5 | 8.6 | |
| M37 | 64.9 | 28.7 | 23.4 | 26.7 | large | −22.2 | 7.8 | −98.7 | 8.8 | |
| M37 | 68.9 | 35.1 | 27.7 | 31.8 | large | −22.1 | 5.0 | −89.5 | 8.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kunst, G.K.; Horacek, M. A Whole Range of Cattle—An Isotopic Perspective on Roman Animal Husbandry in Lower Austria and Burgenland (Austria). Animals 2024, 14, 2624. https://doi.org/10.3390/ani14172624
Kunst GK, Horacek M. A Whole Range of Cattle—An Isotopic Perspective on Roman Animal Husbandry in Lower Austria and Burgenland (Austria). Animals. 2024; 14(17):2624. https://doi.org/10.3390/ani14172624
Chicago/Turabian StyleKunst, Günther Karl, and Micha Horacek. 2024. "A Whole Range of Cattle—An Isotopic Perspective on Roman Animal Husbandry in Lower Austria and Burgenland (Austria)" Animals 14, no. 17: 2624. https://doi.org/10.3390/ani14172624