


Age and Body Size of the Endemic and Critically Endangered Frog Species Rana tavasensis (Baran and Atatür, 1986) in Türkiye


Abstract
Simple Summary
Abstract
1. Introduction
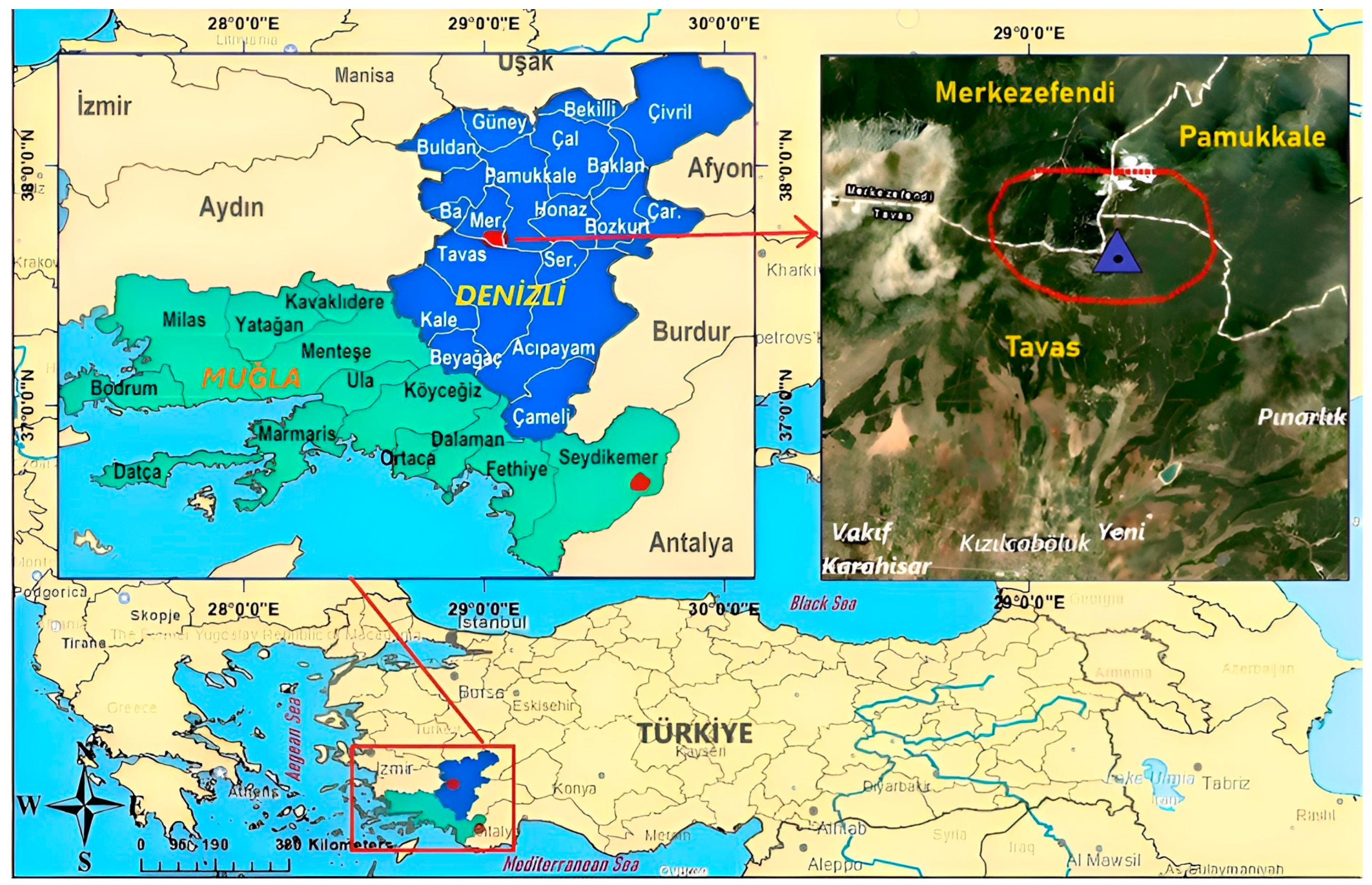
2. Materials and Methods
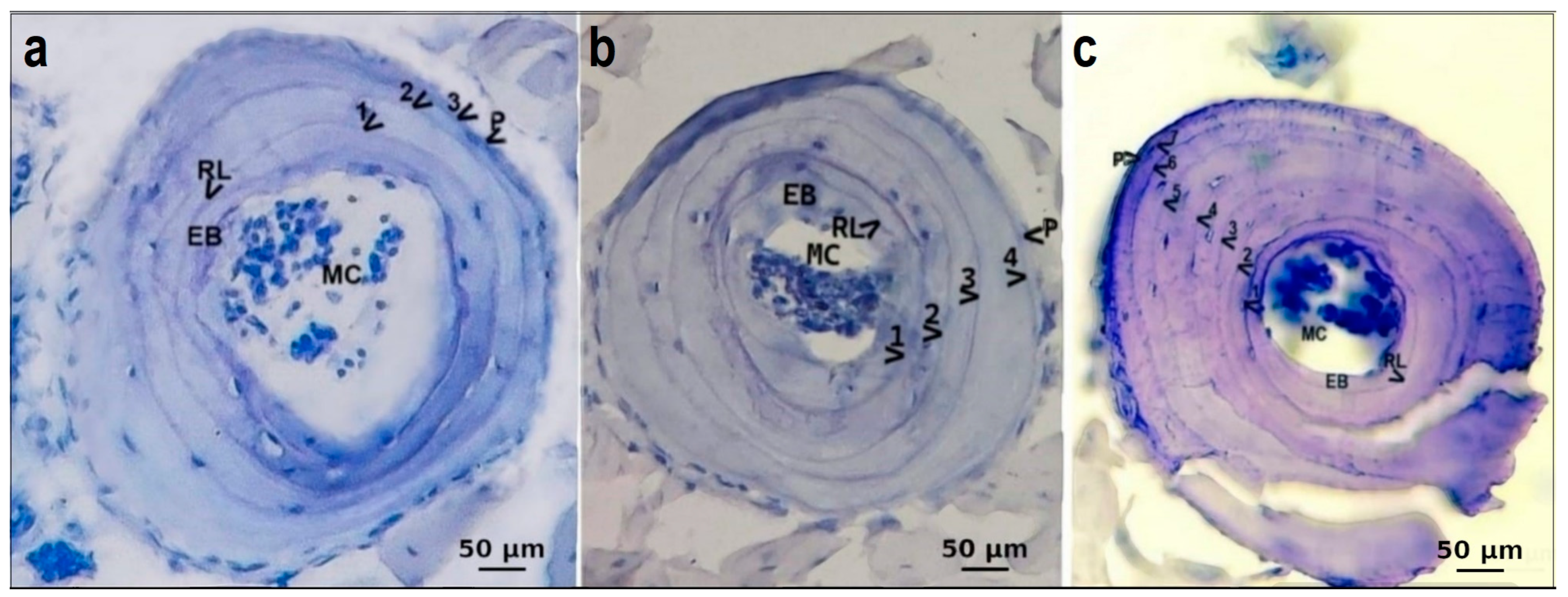
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baran, İ.; Atatür, M.K. A taxonomical survey of the mountain frogs of Anatolia. Amphib. Reptil. 1986, 7, 115–133. [Google Scholar] [CrossRef]
- Veith, M.; Schmidtler, F.; Kosuch, J.; Baran, İ.; Seitz, A. Paleoclimatic changes explain Anatolian mountain frog evolution: A test for alternating vicariance and dispersal event. Mol. Ecol. 2003, 12, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Franzen, M.; Bubmann, M.; Kordges, T.; Thiesmeier, B. Die Amphibien und Reptilien der Südwest-Türkei; Laurenti-Verlag: Berlin, Germany, 2008. (In German) [Google Scholar]
- Ergül-Kalaycı, T.; Özdemir, N. New locality records for Turkish endemic species Rana tavasensis (Baran and Atatür, 1986). J. Cryst. Growth 2018, 3, 77–79. [Google Scholar] [CrossRef]
- Çapar, D.; Başkale, E. Population size of endemic Rana tavasensis in its terra typica, Turkey. Turk. J. Zool. 2016, 40, 729–733. [Google Scholar] [CrossRef]
- IUCN SSC Amphibian Specialist Group. Rana tavasensis. The IUCN Red List of Threatened Species 2023: e.T61875A100225246. Available online: https://www.iucnredlist.org/species/61875/100225246 (accessed on 25 July 2024).
- Lande, R.; Engen, S.; Saether, B.E. Stochastic Population Dynamics in Ecology and Conservation; Oxford University Press: Cary, NC, USA, 2003. [Google Scholar]
- Piferrer, F.; Anastasiadi, D. Age estimation in fishes using epigenetic clocks: Applications to fisheries management and conservation biology. Front. Mar. Sci. 2023, 10, 1062151. [Google Scholar] [CrossRef]
- Khonsue, W.; Matsui, M.; Misawa, Y. Age determination by skeletochronology of Rana nigrovittata, a frog from tropical forest of Thailand. Zool. Sci. 2000, 17, 253–257. [Google Scholar] [CrossRef]
- Smirina, E.M. Age determination and longevity in amphibians. Gerontology 1994, 40, 133–146. [Google Scholar] [CrossRef]
- Guarino, F.M.; Erismis, U.C. Age determination and growth by skeletochronology of Rana holtzi, an endemic frog from Turkey. Ital. J. Zool. 2008, 75, 237–242. [Google Scholar] [CrossRef]
- Guarino, F.M.; de Pous, P.; Crottini, A.; Mezzasalma, M.; Andreone, F. Age structure and growth in a population of Pelobates varaldii (Anura, Pelobatidae) from northwestern Morocco. Amphib. Reptil. 2011, 32, 550–556. [Google Scholar]
- Bionda, C.D.L.; Kost, S.; Salas, N.E.; Lajmanovich, R.C.; Sinsch, U.; Martino, A.L. Age structure, growth and longevity in the common toad, Rhinella arenarum, from Argentina. Acta Herpetol. 2015, 10, 55–62. [Google Scholar]
- Başkale, E.; Ulubeli, S.A.; Kaska, Y. Age structures and growth parameters of the Levantine frog, Pelophylax bedriagae, at different localities in Denizli, Turkey. Acta Herpetol. 2018, 13, 147–154. [Google Scholar]
- Genç, Z.; Tok, C.V. A preliminary study on age determination and examination of some growth parameters in Agile Frog (Rana dalmatina Bonaparte, 1839) (Anura: Ranidae) specimens. Comm. J. Biol. 2021, 5, 59–62. [Google Scholar] [CrossRef]
- Albayrak, M.; Bülbül, U.; Zaman, E.; Koç-Gür, H. Life history traits in a Turkish population of the Agile frog Rana dalmatina Fitzinger in Bonaparte, 1839 (Anura: Ranidae). Acta Zool. Bulg. 2023, 75, 497–503. [Google Scholar]
- Miaud, C.; Üzüm, N.; Avcı, A.; Olgun, K. Age, size and growth of the endemic Anatolian mountain frog Rana holtzi from Turkey. J. Herpetol. 2007, 17, 167–173. [Google Scholar]
- Yıldız, M.Z.; Göçmen, B. Population dynamics, reproduction, and life history traits of Taurus frog, Rana holtzi Werner, 1898 (Anura: Ranidae) in Karagöl (Ulukışla, Niğde), Turkey. Herpetol. Rom. 2012, 6, 1–40. [Google Scholar]
- Kutrup, B.; Özdemir, N.; Bülbül, U.; Çakır, E. A skeletochronological study of age, growth and longevity of Rana macrocnemis populations from four locations at different altitudes in Turkey. Amphibia-Reptilia 2011, 32, 113–118. [Google Scholar] [CrossRef]
- Leskovar, C.; Oromi, N.; Sanuy, D.; Sinsch, U. Demographic life history traits of reproductive natterjack toads (Bufo calamita) vary between northern and southern latitudes. Amphibia-Reptilia 2006, 27, 365–375. [Google Scholar]
- Castanet, J.; Smirina, E.M. Introduction to the skeletochronological method in amphibians and reptiles. Ann. Sci. Nat. Zool. 1990, 11, 191–196. [Google Scholar]
- Castanet, J.; Francillon-Vieillot, H.; Meunier, F.J.; DeRicqlès, A. Bone and individual aging. In Bone, 2nd ed.; Hall, B.K., Ed.; CRC Press: Boca Raton, FL, USA, 1993; Volume 7, pp. 245–283. [Google Scholar]
- Miaud, C.; Joly, P.; Castanet, J. Variation in age structures in a subdivided population of Triturus cristatus. Can. J. Zool. 1993, 71, 1874–1879. [Google Scholar] [CrossRef]
- Castanet, J.; Francillon-Vieillot, H.; Bruce, R.C. Age estimation in desmognathine salamanders assessed by skeletochronology. Herpetologica 1996, 52, 160–171. [Google Scholar]
- Odabaş, Y.; Bülbül, U.; Eroğlu, A.İ.; Koc, H.; Kurnaz, M.; Kutrup, B. Age structure and growth in a Turkish population of the Balkan Green Lizard, Lacerta trilineata Bedriaga, 1886. Herpetozoa 2019, 31, 3–4. [Google Scholar]
- Smirina, E.M. Prospects of age determination by bone layers in Reptilia. Zool. Zhurnal 1974, 53, 111–117. [Google Scholar]
- Aragón, P.; Fitze, P.S. Geographical and temporal body size variation in a reptile: Roles of sex, ecology, phylogeny and ecology structured in phylogeny. PLoS ONE 2014, 9, e104026. [Google Scholar] [CrossRef] [PubMed]
- Bülbül, U.; Kutrup, B.; Eroğlu, A.İ.; Koç, H.; Kurnaz, M.; Odabaş, Y. Life history traits of a Turkish population of the Yellow-bellied Toad, Bombina variegata (Linnaeus, 1758) (Anura: Bombinatoridae). Herpetozoa 2018, 31, 11–19. [Google Scholar]
- Ento, K.; Matsui, M. Estimation of age structure by skeletochronology of a population of Hynobius nebulosus in a breeding season (Amphibia, Urodela). Zool. Sci. 2002, 19, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Revelle, W. Psych: Procedures for Psychological, Psychometric, and Personality Research (Version 2.4.6); Northwestern University: Evanston, IL, USA, 2019; Available online: https://CRAN.R-project.org/package=psych (accessed on 28 July 2024).
- Pituch, K.A.; Stevens, J.P. Applied Multivariate Statistics for the Social Sciences, 6th ed.; Routledge: New York, NY, USA, 2016. [Google Scholar]
- Lovich, J.E.; Gibbons, J.W. A review of techniques for quantifying sexual size dimorphism. Growth Dev. Aging 1992, 56, 269–281. [Google Scholar]
- von Bertalanffy, L. A quantitative theory of organic growth. Hum. Biol. 1938, 10, 181–213. [Google Scholar]
- Miaud, C.; Andreone, F.; Ribéron, A.; De Michelis, S.; Clima, V.; Castanet, J.; Francillon-Vieillot, H.; Guyétant, R. Variations in age, size at maturity and gestation duration among two neighbouring popula-tions of the alpine salamander (Salamandra lanzai). J. Zool. 2001, 254, 251–260. [Google Scholar] [CrossRef]
- Gül, S.; Özdemir, N.; Üzüm, N.; Olgun, K.; Kutrup, B. Body size and age structure of Pelophylax ridibundus populations from two different altitudes in Turkey. Amphibia-Reptilia 2011, 32, 287–292. [Google Scholar]
- Üzüm, N.; Avcı, A.; Özdemir, N.; Ilgaz, Ç.; Olgun, K. Body size and age structure of a breeding population portion of the Urmia salamander, Neurergus crocatus Cope, 1862 (Caudata: Salamandridae). Ital. J. Zool. 2011, 78, 209–214. [Google Scholar] [CrossRef]
- Arısoy, A.G.; Başkale, E. Body size, age structure and survival rates in two populations of the Beyşehir frog Pelophylax caralitanus. Herpetozoa 2019, 32, 195–201. [Google Scholar] [CrossRef]
- Robson, D.S.; Chapman, D.G. Catch curves and mortality rates. Trans. Am. Fish. Soc. 1961, 90, 181–189. [Google Scholar] [CrossRef]
- Seber, G.A.F. The Estimation of Animal Abundance; Hafner Press: New York, NY, USA, 1973. [Google Scholar]
- Patil, I.; Powell, C. ggstatsplot: ‘ggplot2’ Based Plots with Statistical Details. CRAN. 2018. Available online: https://CRAN.R-project.org/package=ggstatsplot (accessed on 28 July 2024).
- R Core Team. R: A Language and Environment for Statistical Computing, Version 4.4.1 “Race for your Life”; R Foundation for Statistical Computing: Vienna, Austria, 2024; Available online: https://cran.r-project.org (accessed on 28 July 2024).
- Guarino, F.M.; Angelini, F.; Cammarota, M. A skeletochronological analysis of three syntopic amphibian species from southern Italy. Amphib. Reptil. 1995, 16, 297–302. [Google Scholar] [CrossRef]
- Esteban, M.; Sanchiz, B. Differential growth and longevity in low and high altitude Rana iberica (Anura, Ranidae). Herpetol. J. 2000, 10, 19–26. [Google Scholar]
- Miaud, C.; Guyétant, R.; Elmberg, J. Variations in life-history traits in the common frog Rana temporaria (Amphibia: Anura): A literature review and new data from the French Alps. J. Zool. Lond. 1999, 249, 61–73. [Google Scholar] [CrossRef]
- Sinsch, U. Review: Skeletochronological assessment of demographic life-history traits in amphibians. Herpetol. J. 2015, 25, 5–13. [Google Scholar]
- Ishchenko, V.G. Problems of demography and declining populations of some Euroasiatic brown frogs. Russ. J. Herpetol. 1996, 3, 143–151. [Google Scholar]
- Düşen, O.; Düşen, S.; Başkale, E. Denizli ili Tavas Kurbağası (Rana tavasensis) tür eylem planı; T.C. Tarım ve Orman Bakanlığı, Doğa Koruma ve Milli Parklar Genel Müdürlüğü: Ankara, Turkey, 2019. [Google Scholar]
- Castanet, J. Age estimation and longevity in reptiles. Gerontology 1994, 40, 174–192. [Google Scholar] [CrossRef]
- Yakın, Y.; Tok, C.V. Age estimation of Anatololacerta anatolica (Werner, 1902) in the vicinity of Çanakkale by skeletochronology vicinity of Çanakkale by skeletochronology, Turk. J. Zool. 2015, 39, 66–73. [Google Scholar]
- Jakob, C.; Seitz, A.; Crivelli, A.J.; Miaud, C. Growth cycle of the marbled newt (Triturus marmoratus) in the Mediterranean region assessed by skeletochronology. Amphib. Reptil. 2002, 23, 407–418. [Google Scholar] [CrossRef]
- Smirina, E.M. Annual layers in bones of Rana temporaria. Zool. Zh. 1972, 51, 1529–1534. [Google Scholar]
- Sinsch, U.; Dehling, J.M. Tropical anurans mature early and die young: Evidence from eight Afromontane Hyperolius species and a meta-analysis. PLoS ONE 2017, 12, e0171666. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age Group | Males | Age Group | Females | ||||
|---|---|---|---|---|---|---|---|
| n | SVL | n | SVL | ||||
| M ± SD | min–max | M ± SD | min–max | ||||
| 3 | 1 | 51.13 | 51.13 | 3 | 3 | 46.12 ± 3.43 | 42.33–49.02 |
| 4 | 1 | 51.97 | 51.97 | 4 | 1 | 49.87 | 49.87 |
| 5 | 2 | 63.24 ± 11.94 | 54.80–71.69 | 5 | 2 | 56.92± 1.29 | 55.48–59.51 |
| 6 | 2 | 61.50 ± 3.48 | 58.02–64.98 | 6 | 5 | 58.12 ± 0.69 | 56.81–59.79 |
| 7 | 1 | 66.16 | 66.16 | 7 | 3 | 61.71 ± 3.57 | 54.71–66.46 |
| 8 | 1 | 68.21 | 68.21 | 8 | 1 | 65.23 | 65.23 |
| 11 | 1 | 69.14 | 69.14 | 11 | 1 | 73.97 | 73.97 |
| 12 | 1 | 71.47 | 71.47 | 12 | 1 | 70.84 | 70.84 |
| Total | 10 | 62.75± 8.02 | 51.13–71.69 | Total | 17 | 58.04± 8.37 | 42.33–73.97 |
| Subadult Males | Juveniles | ||||||
| SVL | SVL | ||||||
| M ± SD | min–max | M ± SD | min–max | ||||
| 2 | 2 | 46.56 ± 5.42 | 42.73–50.40 | 1 | 2 | 23.07 ± 9.64 | 16.95–29.89 |
| Species | Location (Meter above Sea Level) | n (M/F) | Longevity (Years) (M/F) | References |
|---|---|---|---|---|
| R. dalmatina | Çanakkale and Kırklareli (about 200 m) | 11/5 | 4/5 | [15] |
| R. dalmatina | Devrek, Zonguldak (626 m) | 17/13 | 6/8 | [16] |
| R. holtzi | Karagöl, Niğde (2560 and 2580 m) | 21/18 | 8/10.5 | [17] |
| R. holtzi | Karagöl, Niğde (2650 m) | 26/15 | 6/7 | [11] |
| R. holtzi | Karagöl, Niğde (2558 and 2673 m) | 47/42 | 8/9 | [18] |
| R. macrocnemis | Maçka and Hıdırnebi, Trabzon Ovit, Rize Sarıkamış, Kars (350–2850 m) | 100/61 | 8–10 | [19] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bülbül, U.; Başkale, E.; Özkan, H. Age and Body Size of the Endemic and Critically Endangered Frog Species Rana tavasensis (Baran and Atatür, 1986) in Türkiye. Animals 2024, 14, 2703. https://doi.org/10.3390/ani14182703
Bülbül U, Başkale E, Özkan H. Age and Body Size of the Endemic and Critically Endangered Frog Species Rana tavasensis (Baran and Atatür, 1986) in Türkiye. Animals. 2024; 14(18):2703. https://doi.org/10.3390/ani14182703
Chicago/Turabian StyleBülbül, Ufuk, Eyup Başkale, and Hatice Özkan. 2024. "Age and Body Size of the Endemic and Critically Endangered Frog Species Rana tavasensis (Baran and Atatür, 1986) in Türkiye" Animals 14, no. 18: 2703. https://doi.org/10.3390/ani14182703
APA StyleBülbül, U., Başkale, E., & Özkan, H. (2024). Age and Body Size of the Endemic and Critically Endangered Frog Species Rana tavasensis (Baran and Atatür, 1986) in Türkiye. Animals, 14(18), 2703. https://doi.org/10.3390/ani14182703