
Nuancing ‘Emotional’ Social Play: Does Play Behaviour Always Underlie a Positive Emotional State?



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
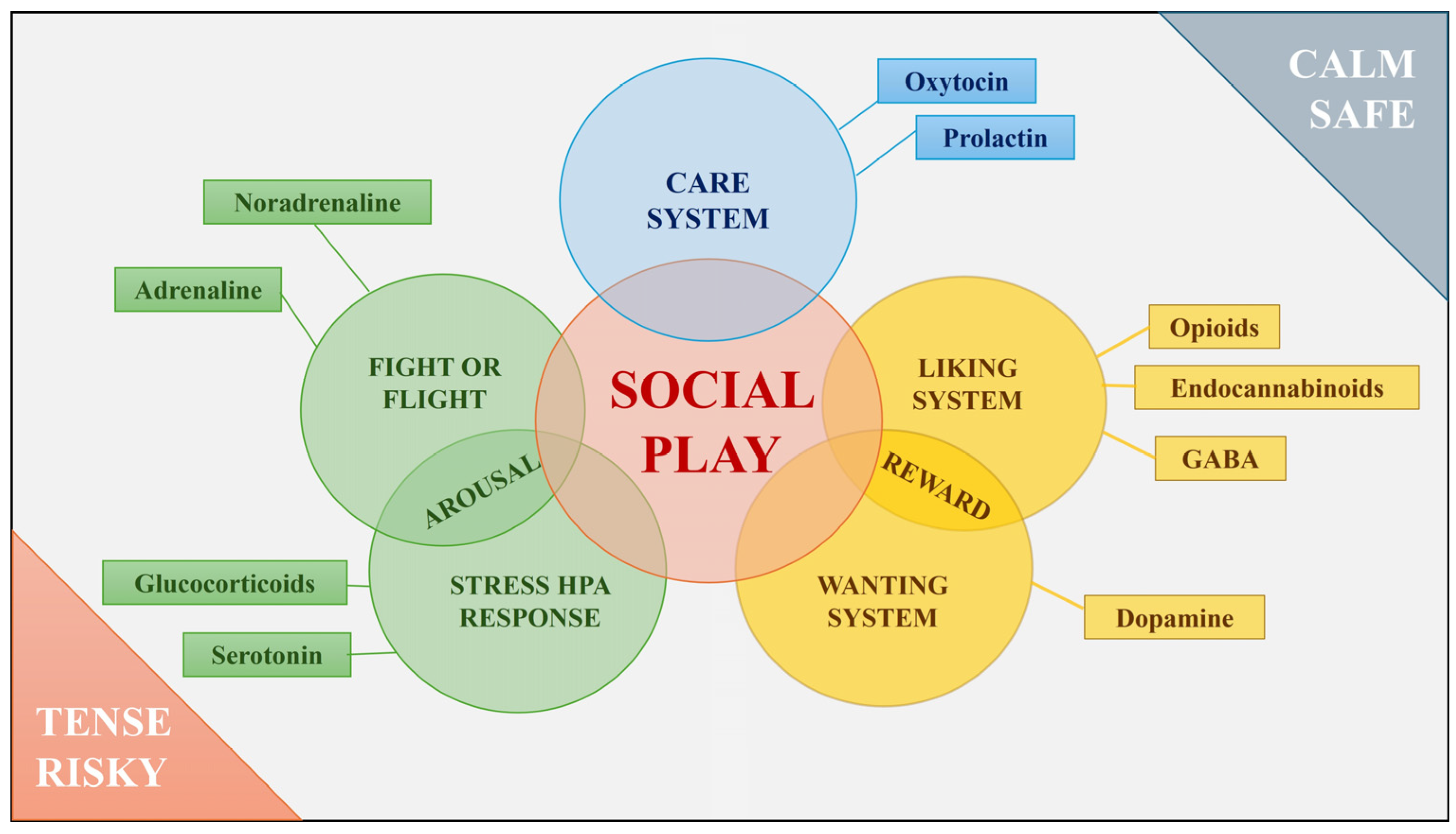
2. Social Play: What Are the Underlying Emotional Systems?
2.1. Play and Reward
2.2. Play and Social Attachment
2.3. Play and Arousal
3. The Dual Nature of Social Play: Doppelgänger of Aggression?
Play Fighting or Play for Fighting: Aggressive Play or Playful Aggression?
4. Play and Its Structure
5. Playful Signals: What Kind of Message Do They Convey?
6. Rapid Facial Mimicry: The Transfer of Mood, but What Mood?
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lancy, D.F. Play in species adaptation. Ann. Rev. Anthropol. 1980, 9, 471–495. [Google Scholar] [CrossRef]
- Pellis, S.M.; Burghardt, G.M. Play and exploration. In APA Handbook of Comparative Psychology. Concepts, History, and Methods; Call, J., Burghardt, G.M., Pepperberg, I., Snowdon, C., Zentall, T., Eds.; American Psychological Association: Washington, DC, USA, 2017; Volume 2, pp. 699–722. [Google Scholar]
- Pellis, S.M.; Pellis, V.C.; Pelletier, A.; Leca, J.B. Is play a behavior system, and, if so, what kind? Behav. Proc. 2019, 160, 1–9. [Google Scholar] [CrossRef]
- Burghardt, G.M. The Genesis of Animal Play: Testing the Limits; MIT Press: Cambridge, MA, USA, 2005. [Google Scholar]
- Smith, P.K. Play fighting and real fighting. Perspectives on their relationship. In New Aspects of Human Ethology; Schmitt, A., Atzwanger, K., Grammar, K., Schäfer, K., Eds.; Plenum Press: New York, NY, USA, 1997; pp. 47–64. [Google Scholar]
- Panksepp, J. The basic emotional circuits of mammalian brains: Do animals have affective lives? Neurosci. Biobehav. Rev. 2011, 35, 1791–1804. [Google Scholar] [CrossRef]
- Porges, S.W. Play as neural exercise: Insights from the Polyvagal theory. In The Power of Play for Mind Brain Health; Mindgains.org. GAINS: Philadelphia, PA, USA, 2015; pp. 3–7. [Google Scholar]
- Palagi, E.; Burghardt, G.M.; Smuts, B.; Cordoni, G.; Dall’Olio, S.; Fouts, H.N.; Řeháková-Petrů, M.; Siviy, S.M.; Pellis, S.M. Rough-and-tumble play as a window on animal communication. Biol. Rev. 2016, 91, 311–327. [Google Scholar] [CrossRef] [PubMed]
- van Hooff, J.A.R.A.M.; Preuschoft, S. Laughter and smiling: The intertwining of nature and culture. In Animal Social Complexity: Intelligence, Culture, and Individualized Societies; de Waal, F.B.M., Tyack, P.L., Eds.; Harvard University Press: Cambridge, MA, USA, 2003; pp. 261–287. [Google Scholar]
- Davila-Ross, M.; Dezecache, G. The complexity and phylogenetic continuity of laughter and smiles in hominids. Front. Psychol. 2021, 12, 2065. [Google Scholar] [CrossRef]
- Cordoni, G.; Brescini, M.; Pirarba, L.; Giaretto, F.; Norscia, I. Functional and morphological differences in the play face and full play face in lowland gorillas (Gorilla gorilla gorilla), a Hominid species: Implications for the evolutionary roots of smile and laughter. Am. J. Biol. Anthropol. 2024; under review. [Google Scholar]
- Porges, S.W. The Polyvagal theory: New insights into adaptive reactions of the autonomic nervous system. Clev. Clin. J Med. 2009, 76, 86. [Google Scholar] [CrossRef]
- Gordon, I.; Zagoory-Sharon, O.; Leckman, J.F.; Feldman, R. Prolactin, oxytocin, and the development of paternal behavior across the first six months of fatherhood. Horm. Behav. 2010, 58, 513–518. [Google Scholar] [CrossRef]
- Meaney, M.J.; Viau, V.; Bhatnagar, S.; Betito, K.; Iny, L.J.; O’Donnell, D.; Mitchell, J.B. Cellular mechanisms underlying the development and expression of individual differences in the hypothalamic-pituitary-adrenal stress response. J. Steroid Biochem. Mol. Biol. 1991, 39, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Vanderschuren, L.J.; Achterberg, E.J.; Trezza, V. The neurobiology of social play and its rewarding value in rats. Neurosci. Biobehav. Rev. 2016, 70, 86–105. [Google Scholar] [CrossRef]
- Papilloud, A.; Guillot de Suduiraut, I.; Zanoletti, O.; Grosse, J.; Sandi, C. Peripubertal stress increases play fighting at adolescence and modulates nucleus accumbens CB1 receptor expression and mitochondrial function in the amygdala. Transl. Psychiatry 2018, 8, 156. [Google Scholar] [CrossRef]
- Achterberg, E.M.; Vanderschuren, L.J. Treatment with low doses of nicotine but not alcohol affects social play reward in rats. Int. J. Play 2020, 9, 39–57. [Google Scholar] [CrossRef]
- Alia-Klein, N.; Gan, G.; Gilam, G.; Bezek, J.; Bruno, A.; Denson, T.F.; Hendler, T.; Lowe, L.; Mariotti, V.; Muscatello, M.R.; et al. The feeling of anger: From brain networks to linguistic expressions. Neurosci. Biobehav. Rev. 2020, 108, 480–497. [Google Scholar] [CrossRef] [PubMed]
- Hines, M.; Kaufman, F.R. Androgen and the development of human sex-typical behavior: Rough-and-tumble play and sex of preferred playmates in children with congenital adrenal hyperplasia (CAH). Child Dev. 1994, 65, 1042–1053. [Google Scholar] [CrossRef]
- Sánchez-Martın, J.R.; Fano, E.; Ahedo, L.; Cardas, J.; Brain, P.F.; Azpíroz, A. Relating testosterone levels and free play social behavior in male and female preschool children. Psychoneuroendocrinology 2000, 25, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Roberts, B.A.; Martel, M.M. Prenatal testosterone and preschool Disruptive Behavior Disorders. Personal. Individ. Differ. 2013, 55, 962–966. [Google Scholar] [CrossRef] [PubMed]
- Menon, R.; Süß, T.; de Moura Oliveira, V.E.; Neumann, I.D.; Bludau, A. Neurobiology of the lateral septum: Regulation of social behavior. Trends Neurosci. 2022, 45, 27–40. [Google Scholar] [CrossRef]
- Achterberg, E.M.; Vanderschuren, L.J. The neurobiology of social play behaviour: Past, present and future. Neurosci. Biobehav. Rev. 2023, 152, 105319. [Google Scholar] [CrossRef]
- Trezza, V.; Baarendse, P.J.J.; Vanderschuren, L.J.M.J. The pleasures of play: Pharmacological insights into social reward mechanisms. Trends Pharmacol. Sci. 2010, 31, 463–469. [Google Scholar] [CrossRef]
- Trezza, V.; Campolongo, P.; Vanderschuren, L.J.M.J. Evaluating the rewarding nature of social interactions in laboratory animals. Dev. Cogn. Neurosci. 2011, 1, 444–458. [Google Scholar] [CrossRef]
- Cordoni, G.; Gioia, M.; Demuru, E.; Norscia, I. The dark side of play: Play fighting as a substitute for real fighting in domestic pigs (Sus scrofa). Anim. Behav. 2021, 175, 21–31. [Google Scholar] [CrossRef]
- Cordoni, G.; Ciarcelluti, G.; Pasqualotto, A.; Perri, A.; Bissiato, V.; Norscia, I. Is it for real? Structural differences between play and real fighting in adult chimpanzees (Pan troglodytes). Am. J. Primatol. 2023, 85, e23537. [Google Scholar] [CrossRef] [PubMed]
- Cordoni, G.; Ciantia, A.; Guéry, J.P.; Mulot, B.; Norscia, I. Rapid facial mimicry in Platyrrhini: Play face replication in spider monkeys (Ateles fusciceps, Ateles hybridus, and Ateles paniscus). Am. J. Primatol. 2024, 86, e23607. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.S.; Pournajafi-Nazarloo, H.; Kramer, K.M.; Ziegler, T.E.; White-Traut, R.; Bello, D.; Schwertz, D. Oxytocin: Behavioral associations and potential as a salivary biomarker. Ann. N. Y. Acad. Sci. 2007, 1098, 312–322. [Google Scholar] [CrossRef] [PubMed]
- White-Traut, R.; Watanabe, K.; Pournajafi-Nazarloo, H.; Schwertz, D.; Bell, A.; Carter, C.S. Detection of salivary oxytocin levels in lactating women. Dev. Psychobiol. 2009, 51, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Bredewold, R.; Schiavo, J.K.; van der Hart, M.; Verreij, M.; Veenema, A.H. Dynamic changes in extracellular release of GABA and glutamate in the lateral septum during social play behavior in juvenile rats: Implications for sex-specific regulation of social play behavior. Neuroscience 2015, 307, 117–127. [Google Scholar] [CrossRef]
- Cutuli, D.; Berretta, E.; Caporali, P.; Sampedro-Piquero, P.; De Bartolo, P.; Laricchiuta, D.; Gelfo, F.; Pesoli, M.; Foti, F.; Vecchioli, S.F.; et al. Effects of pre-reproductive maternal enrichment on maternal care, offspring’s play behavior and oxytocinergic neurons. Neuropharmacology 2019, 145A, 99–113. [Google Scholar] [CrossRef]
- Shimada, M.; Sueur, C. Social play among juvenile wild Japanese macaques (Macaca fuscata) strengthens their social bonds. Am. J. Primatol. 2018, 80, e22728. [Google Scholar] [CrossRef]
- Drea, C.M.; Hawk, J.E.; Glickman, S.E. Aggression decreases as play emerges in infant spotted hyaenas: Preparation for joining the clan. Anim. Behav. 1996, 51, 1323–1336. [Google Scholar] [CrossRef]
- Lee, P.C. Allomothering among African elephants. Anim. Behav. 1987, 35, 278–291. [Google Scholar] [CrossRef]
- Raviv, L.; Jacobson, S.L.; Plotnik, J.M.; Bowman, J.; Lynch, V.; Benítez-Burraco, A. Elephants as an animal model for self-domestication. Proc. Natl. Acad. Sci. USA 2023, 120, e2208607120. [Google Scholar] [CrossRef]
- Norscia, I.; Hecker, M.; Caselli, M.; Collarini, E.; Gallego Aldama, B.; Borragán Santos, S.; Cordoni, G. Social play in African savannah elephants may inform selection against aggression. Curr. Zool. 2024, zoae009. [Google Scholar] [CrossRef]
- Pellis, S.M.; Pellis, V.C. The Playful Brain: Venturing to the Limits of Neuroscience; Oneworld Publications: London, UK, 2009. [Google Scholar]
- Antonacci, D.; Norscia, I.; Palagi, E. Stranger to familiar: Wild strepsirhines manage xenophobia by playing. PLoS ONE 2010, 5, e13218. [Google Scholar] [CrossRef] [PubMed]
- Cordoni, G.; Palagi, E. Ontogenetic trajectories of chimpanzee social play: Similarities with humans. PLoS ONE 2011, 6, e27344. [Google Scholar] [CrossRef]
- Pellis, S.M.; Pellis, V.C. What is play fighting and what is it good for? Learn. Behav. 2017, 45, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Caselli, M.; Malaman, B.; Cordoni, G.; Guéry, J.P.; Kok, J.; Demuru, E.; Norscia, I. Not lost in translation: Changes in social dynamics in Bonobos after colony relocation and fusion with another group. App. Anim. Behav. Sci. 2023, 261, 105905. [Google Scholar] [CrossRef]
- Pellis, S.M.; Hastings, E.; Shimizu, T.; Kamitakahara, H.; Komorowska, J.; Forgie, M.L.; Kolb, B. The effects of orbital frontal cortex damage on the modulation of defensive responses by rats in playful and non-playful social contexts. Behav. Neurosci. 2006, 120, 72–84. [Google Scholar] [CrossRef]
- Bell, H.C.; McCaffrey, D.R.; Forgie, M.L.; Kolb, B.; Pellis, S.M. The role of the medial prefrontal cortex in the play fighting of rats. Behav. Neurosci. 2009, 123, 1158–1168. [Google Scholar] [CrossRef] [PubMed]
- Achterberg, E.J.M.; Van Kerkhof, L.W.M.; Damsteegt, R.; Trezza, V.; Vanderschuren, L.J.M.J. Methylphenidate and atomoxetine inhibit social play behavior through prefrontal and subcortical limbic mechanisms in rats. J. Neurosci. 2015, 35, 161–169. [Google Scholar] [CrossRef]
- Monaghan, E.P.; Glickman, S.E. Hormones and aggressive behavior. In Behavioral Endocrinology; Becker, J.B., Breedlove, S.M., Crews, D., Eds.; MIT Press: Cambridge, MA, USA, 1992; pp. 261–286. [Google Scholar]
- Archer, J. The Behavioural Biology of Aggression; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Goldstein, J.H. Sex differences in aggressive play and toy preference. In Of Mice and Women: Aspects of Female Aggression; Bjorkqvist, K., Niemela, P., Eds.; Academic Press Inc.: San Diego, CA, USA, 1992; pp. 65–76. [Google Scholar]
- Goy, R.W.; Deputte, B.L. The effects of diethylstilbestrol (DES) before birth on the development of masculine behavior in juvenile female rhesus monkeys. Horm. Behav. 1996, 30, 379–386. [Google Scholar] [CrossRef]
- Wallen, K. Hormonal influences on sexually differentiated behavior in nonhuman primates. Front. Neuroendocrinol. 2005, 26, 7–26. [Google Scholar] [CrossRef]
- Beatty, W.W.; Dodge, A.M.; Dodge, L.J.; White, K. Psychomotor stimulants, social deprivation and play in juvenile rats. Pharmacol. Biochem. Behav. 1982, 16, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Pompili, M.; Serafini, G.; Innamorati, M.; Möller-Leimkühler, A.M.; Giupponi, G.; Girardi, P.; Tatarelli, R.; Lester, D. The hypothalamic-pituitary-adrenal axis and serotonin abnormalities: A selective overview for the implications of suicide prevention. Eur. Arch. Psychiatry Clin. Neurosci. 2010, 260, 583–600. [Google Scholar] [CrossRef]
- Gemmel, M.; Hazlett, M.; Bögi, E.; De Lacalle, S.; Hill, L.A.; Kokras, N.; Hammond, G.L.; Dalla, C.; Charlier, T.D.; Pawluski, J.L. Perinatal fluoxetine effects on social play, the HPA system, and hippocampal plasticity in pre-adolescent male and female rats: Interactions with pre-gestational maternal stress. Psychoneuroendocrinology 2017, 84, 159–171. [Google Scholar] [CrossRef]
- Pariante, C.M.; Lightman, S.L. The HPA axis in major depression: Classical theories and new developments. Trends Neurosci. 2008, 31, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Mustoe, A.C.; Taylor, J.H.; Birnie, A.K.; Huffman, M.C.; French, J.A. Gestational cortisol and social play shape development of marmosets’ HPA functioning and behavioral responses to stressors. Dev. Psychobiol. 2014, 56, 1229–1243. [Google Scholar] [CrossRef]
- Olivera-Figueroa, L.A.; Juster, R.P.; Morin-Major, J.K.; Marin, M.F.; Lupien, S.J. A time to be stressed? Time perspectives and cortisol dynamics among healthy adults. Biol. Psychol. 2015, 111, 90–99. [Google Scholar] [CrossRef]
- Chronister, B.N.; Gonzalez, E.; Lopez-Paredes, D.; Suarez-Torres, J.; Gahagan, S.; Martinez, D.; Barros, J.; Jacobs, D.R., Jr.; Checkoway, H.; Suarez-Lopez, J.R. Testosterone, estradiol, DHEA and cortisol in relation to anxiety and depression scores in adolescents. J. Affect. Dis. 2021, 294, 838–846. [Google Scholar] [CrossRef] [PubMed]
- Burleson, C.A.; Pedersen, R.W.; Seddighi, S.; DeBusk, L.E.; Burghardt, G.M.; Cooper, M.A. Social play in juvenile hamsters alters dendritic morphology in the medial prefrontal cortex and attenuates effects of social stress in adulthood. Behav. Neurosci. 2016, 130, 437–447. [Google Scholar] [CrossRef]
- Klein, Z.A.; Padow, V.A.; Romeo, R.D. The effects of stress on play and home cage behaviors in adolescent male rats. Dev. Psychobiol. 2010, 52, 62–70. [Google Scholar] [CrossRef]
- Hausberger, M.; Fureix, C.; Bourjade, M.; Wessel-Robert, S.; Richard-Yris, M.A. On the significance of adult play: What does social play tell us about adult horse welfare? Naturwissenschaften 2012, 99, 291–302. [Google Scholar] [CrossRef]
- Held, S.D.; Špinka, M. Animal play and animal welfare. Anim. Behav. 2011, 81, 891–899. [Google Scholar] [CrossRef]
- Palagi, E.; Cordoni, G.; Borgognini Tarli, S.M. Immediate and delayed benefits of play behaviour: New evidence from chimpanzees. Ethology 2004, 110, 949–962. [Google Scholar] [CrossRef]
- Yamanashi, Y.; Nogami, E.; Teramoto, M.; Morimura, N.; Hirata, S. Adult-adult social play in captive chimpanzees: Is it indicative of positive animal welfare? App. Anim. Behav. Sci. 2018, 199, 75–83. [Google Scholar] [CrossRef]
- Palagi, E.; Paoli, T.; Borgognini Tarli, S.M. Short-term benefits of play behavior and conflict prevention in Pan paniscus. Int. J. Primatol. 2006, 27, 1257–1270. [Google Scholar] [CrossRef]
- Palagi, E.; Antonacci, D.; Cordoni, G. Fine-tuning of social play in juvenile lowland gorillas (Gorilla gorilla gorilla). Dev. Psychobiol. 2007, 49, 433–445. [Google Scholar] [CrossRef] [PubMed]
- Špinka, M.; Newberry, R.C.; Bekoff, M. Mammalian play: Training for the unexpected. Q. Rev. Biol. 2001, 76, 141–167. [Google Scholar] [CrossRef]
- Oliveira, A.F.S.; Rossi, A.O.; Silva, L.F.R.; Lau, M.C.; Barreto, R.E. Play behaviour in nonhuman animals and the animal welfare issue. J. Ethol. 2010, 28, 1–5. [Google Scholar] [CrossRef]
- Potegal, M.; Nordman, J.C. Non-angry aggressive arousal and angriffsberietschaft: A narrative review of the phenomenology and physiology of proactive/offensive aggression motivation and escalation in people and other animals. Neurosci. Biobehav. Rev. 2023, 147, 105110. [Google Scholar] [CrossRef]
- De Waal, F.B. Primates—A natural heritage of conflict resolution. Science 2000, 289, 586–590. [Google Scholar] [CrossRef]
- Hart, J.L.; Tannock, M.T. Playful aggression in early childhood settings. Child. Aust. 2013, 38, 106–114. [Google Scholar] [CrossRef]
- Boulton, M.J. A comparison of adults’ and children’s abilities to distinguish between aggressive and playful fighting in middle school pupils: Implications for playground supervision and behaviour management. Educ. Stud. 1993, 19, 193–203. [Google Scholar] [CrossRef]
- Pellegrini, A.D. Rough-and-tumble play from childhood through adolescence: Development and possible functions. In Blackwell Handbook of Social Development, 1st ed.; Smith, P.K., Hart, C., Eds.; Blackwell: Hoboken, NJ, USA, 2002; pp. 438–453. [Google Scholar]
- Symons, D. Aggressive play and communication in rhesus monkeys (Macaca mulatto). Am. Zool. 1974, 14, 317–322. [Google Scholar] [CrossRef]
- Eckert, J.; Winkler, S.L.; Cartmill, E.A. Just kidding: The evolutionary roots of playful teasing. Biol. Lett. 2020, 16, 20200370. [Google Scholar] [CrossRef] [PubMed]
- Paquette, D. Fighting and playfighting in captive adolescent chimpanzees. Aggress. Behav. 1994, 20, 49–65. [Google Scholar] [CrossRef]
- Palagi, E.; Cordoni, G. The right time to happen: Play developmental divergence in the two Pan species. PLoS ONE 2012, 7, e52767. [Google Scholar] [CrossRef]
- Bauer, E.B.; Smuts, B.B. Cooperation and competition during dyadic play in domestic dogs, Canis familiaris. Anim Behav. 2007, 73, 489–499. [Google Scholar] [CrossRef]
- Fouts, H.N.; Hallam, R.A.; Purandare, S. Gender segregation in early-childhood social play among the Bofi Foragers and Bofi Farmers in Central Africa. Am. J. Play 2013, 5, 333–356. [Google Scholar]
- Briffa, M.; Lane, S.M. The role of skill in animal contests: A neglected component of fighting ability. Proc. R. Soc. Lond. B Biol. Sci. 2017, 284, 20171596. [Google Scholar] [CrossRef]
- Cordoni, G.; Nicotra, V.; Palagi, E. Unveiling the ‘secret’ of dog play success: Asymmetry and signals. J. Comp. Psychol. 2016, 130, 278–287. [Google Scholar] [CrossRef]
- Cordoni, G.; Norscia, I.; Bobbio, M.; Palagi, E. Differences in play can illuminate differences in affiliation: A comparative study on chimpanzees and gorillas. PLoS ONE 2018, 13, e0193096. [Google Scholar] [CrossRef]
- Ward, C.; Bauer, E.B.; Smuts, B.B. Partner preferences and asymmetries in social play among domestic dog, Canis lupus familiaris, littermates. Anim. Behav. 2008, 76, 1187–1199. [Google Scholar] [CrossRef]
- Bekoff, M. Social play and play-soliciting by infant canids. Am. Zool. 1974, 14, 323–340. [Google Scholar] [CrossRef]
- Collarini, E.; Gioia, M.; Cordoni, G.; Norscia, I. Does the domestication syndrome apply to the domestic pig? Not completely. Animals 2022, 12, 2458. [Google Scholar] [CrossRef] [PubMed]
- De Vries, H. An improved test of linearity in dominance hierarchies containing unknown or tied relationships. Anim. Behav. 1995, 50, 1375–1389. [Google Scholar] [CrossRef]
- Kendall, M.G.; Gibbons, J.D. Rank Correlation Methods, 5th ed.; Griffin: London, UK, 1955. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Keylock, C.J. Simpson diversity and the Shannon-Wiener index as special cases of a generalized entropy. Oikos 2005, 109, 203–207. [Google Scholar] [CrossRef]
- Maglieri, V.; Palagi, E. Wolves and dogs unleashed: Exploring differences and similarities in adult play. Int. J. Play 2024, 1–17. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Baltimore, MD, USA, 1949. [Google Scholar]
- Bekoff, M. Play signals as punctuation: The structure of social play in canids. Behaviour 1995, 132, 419–429. [Google Scholar] [CrossRef]
- Waller, B.M.; Dunbar, R.I. Differential behavioural effects of silent bared teeth display and relaxed open mouth display in chimpanzees (Pan troglodytes). Ethology 2005, 111, 129–142. [Google Scholar] [CrossRef]
- Cordoni, G.; Palagi, E. Fair play and honest signals in immature chimpanzees. Atti della Società Toscana di Scienze Naturali Serie B 2012, 119, 97–101. [Google Scholar] [CrossRef]
- Bekoff, M.; Allen, C. Intentional communication and social play: How and why animals negotiate and agree to play. In Animal Play: Evolutionary, Comparative, and Ecological Perspectives; Byers, J.A., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 97–114. [Google Scholar]
- Byosiere, S.E.; Espinosa, J.; Smuts, B. Investigating the function of play bows in adult pet dogs (Canis lupus familiaris). Behav. Proc. 2016, 125, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Poole, J.H.; Granli, P. The visual, tactile and acoustic signals of play in African savannah elephants. In Endangered Elephants; Past, Present and Future; Jayewardene, J., Ed.; Biodiversity & Elephant Conservation Trust: Colombo, Sri Lanka, 2004; pp. 44–50. [Google Scholar]
- Burke, C.J.; Euston, D.R.; Pellis, S.M. What do you hear, what do you say? Ultrasonic calls as signals during play fighting in rats. Int. J. Play 2020, 9, 92–107. [Google Scholar] [CrossRef]
- Davila-Ross, M.; Palagi, E. Laughter, play faces and mimicry in animals: Evolution and social functions. Phil. Trans. R. Soc. B 2022, 377, 20210177. [Google Scholar] [CrossRef]
- Waller, B.M.; Micheletta, J. Facial expression in nonhuman animals. Emot. Rev. 2013, 5, 54–59. [Google Scholar] [CrossRef]
- Pellis, S.M.; Field, E.F.; Smith, L.K.; Pellis, V.C. Multiple differences in the play fighting of male and female rats. Implications for the causes and functions of play. Neurosci. Biobehav. Rev. 1997, 21, 105–120. [Google Scholar] [CrossRef]
- Owren, M.J.; Bachorowski, J.A. Reconsidering the evolution of nonlinguistic communication: The case of laughter. J. Nonverb. Behav. 2003, 27, 183–200. [Google Scholar] [CrossRef]
- Mehu, M.; Grammer, K.; Dunbar, R.I. Smiles when sharing. Evol. Hum. Behav. 2007, 28, 415–422. [Google Scholar] [CrossRef]
- Gervais, M.; Wilson, D.S. The evolution and functions of laughter and humor: A synthetic approach. Q. Rev. Biol. 2005, 80, 395–430. [Google Scholar] [CrossRef] [PubMed]
- Davila-Ross, M.; Allcock, B.; Thomas, C.; Bard, K.A. Aping expressions? Chimpanzees produce distinct laugh types when responding to laughter of others. Emotion 2011, 11, 1013. [Google Scholar] [CrossRef]
- Ekman, P. Darwin, deception, and facial expression. Ann. N. Y. Acad. Sci. 2003, 1000, 205–221. [Google Scholar] [CrossRef]
- Ambadar, Z.; Cohn, J.F.; Reed, L.I. All smiles are not created equal: Morphology and timing of smiles perceived as amused, polite, and embarrassed/nervous. J. Nonverbal Behav. 2009, 33, 17–34. [Google Scholar] [CrossRef] [PubMed]
- Scott, S.K.; Lavan, N.; Chen, S.; McGettigan, C. The social life of laughter. Trends Cogn. Sci. 2014, 18, 618–620. [Google Scholar] [CrossRef]
- Wood, A.; Niedenthal, P. Developing a social functional account of laughter. Soc. Personal. Psychol. Comp. 2018, 12, e12383. [Google Scholar] [CrossRef]
- Liu, X.; Chen, Y.; Ge, J.; Mao, L. Funny or Angry? Neural Correlates of Individual Differences in Aggressive Humor Processing. Front. Psychol. 2019, 10, 1849. [Google Scholar] [CrossRef]
- Emory, G.R.; Payne, R.G.; Chance, M.R.A. Observations on a newly described usage of the primate play face. Behav. Proc. 1979, 4, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Palagi, E.; Celeghin, A.; Tamietto, M.; Winkielman, P.; Norscia, I. The neuroethology of spontaneous mimicry and emotional contagion in human and non-human animals. Neurosci. Biobehav. Rev. 2020, 111, 149–165. [Google Scholar] [CrossRef]
- Waller, B.M.; Cherry, L. Facilitating play through communication: Significance of teeth exposure in the gorilla play face. Am. J. Primatol. 2012, 74, 157–164. [Google Scholar] [CrossRef]
- Thierry, B.; Demaria, C.; Preuschoft, S.; Desportes, C. Structural convergence between silent bared-teeth display and relaxed open-mouth display in the Tonkean macaque (Macaca tonkeana). Folia Primatol. 1989, 52, 178–184. [Google Scholar] [CrossRef]
- Fagen, R. Animal Play Behavior; Oxford University Press: Oxford, UK, 1981. [Google Scholar]
- Thompson, K.V. Self-assessment in juvenile play. In Animal Play: Evolutionary, Comparative and Ecological Perspective; Bekoff, M., Byers, J.A., Eds.; Cambridge University Press: Cambridge, UK, 1998; pp. 183–204. [Google Scholar]
- Sarra, S.; Otta, E. Different types of smiles and laughter in preschool children. Psychol. Rep. 2001, 89, 547–558. [Google Scholar] [CrossRef]
- Ekman, P.; Friesen, W.V.; Hager, J.C. Facial Action Coding System; Research Nexus: Salt Lake City, UT, USA, 2002. [Google Scholar]
- Baltrušaitis, T.; Zadeh, A.; Lim, Y.C.; Morency, L.P. OpenFace 2.0: Facial behavior analysis toolkit. In Proceedings of the IEEE International Conference on Automatic Face and Gesture Recognition, Xi’an, China, 15–19 May 2018. [Google Scholar]
- Waller, B.M.; Julle-Daniere, E.; Micheletta, J. Measuring the evolution of facial ‘expression’ using multi-species FACS. Neurosci. Biobehav. Rev. 2020, 113, 1–11. [Google Scholar] [CrossRef]
- Iacoboni, M. Imitation, empathy, and mirror neurons. Ann. Rev. Psychol. 2009, 60, 653–670. [Google Scholar] [CrossRef]
- Gallese, V.; Fadiga, L.; Fogassi, L.; Rizzolatti, G. Action recognition in the premotor cortex. Brain 1996, 119, 593–609. [Google Scholar] [CrossRef]
- Preston, S.D.; de Waal, F.B.M. Empathy: Its ultimate and proximate bases. Behav. Brain Sci. 2002, 25, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, P.F.; Gallese, V.; Rizzolatti, G.; Fogassi, L. Mirror neurons responding to the observation of ingestive and communicative mouth actions in the monkey ventral premotor cortex. Eur. J. Neurosci. 2003, 17, 1703–1714. [Google Scholar] [CrossRef] [PubMed]
- De Waal, F.B.M.; Preston, S.D. Mammalian empathy: Behavioural manifestations and neural basis. Nat. Rev. Neurosci. 2017, 18, 498–509. [Google Scholar] [CrossRef] [PubMed]
- Rizzolatti, G.; Caruana, F. Some considerations on de Waal and Preston review. Nat. Rev. Neurosci. 2017, 18, 769. [Google Scholar] [CrossRef]
- Hecht, E.E.; Gutman, D.A.; Preuss, T.M.; Sanchez, M.M.; Parr, L.A.; Rilling, J.K. Process versus product in social learning: Comparative diffusion tensor imaging of neural systems for action execution–observation matching in macaques, chimpanzees, and humans. Cereb. Cortex 2013, 23, 1014–1024. [Google Scholar] [CrossRef]
- Hess, U.; Fischer, A. Emotional mimicry as social regulation. Personal. Soc. Psychol. Rev. 2013, 17, 142–157. [Google Scholar] [CrossRef]
- Kraaijenvanger, E.J.; Hofman, D.; Bos, P.A. A neuroendocrine account of facial mimicry and its dynamic modulation. Neurosci. Biobehav. Rev. 2017, 77, 98–106. [Google Scholar] [CrossRef]
- Weisfeld, G.E. The adaptive value of humor and laughter. Ethol. Sociobiol. 1993, 14, 141–169. [Google Scholar] [CrossRef]
- Davila-Ross, M.; Menzler, S.; Zimmermann, E. Rapid facial mimicry in orangutan play. Biol. Lett. 2008, 4, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Palagi, E.; Norscia, I.; Pressi, S.; Cordoni, G. Facial mimicry and play: A comparative study in chimpanzees and gorillas. Emotion 2019, 19, 665. [Google Scholar] [CrossRef] [PubMed]
- Bresciani, C.; Cordoni, G.; Palagi, E. Playing together, laughing together: Rapid facial mimicry and social sensitivity in lowland gorillas. Curr. Zool. 2022, 68, 560–569. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Zanoli, A.; Caselli, M.; Norscia, I.; Palagi, E. The face never lies: Facial expressions and mimicry modulate playful interactions in wild geladas. Behav. Ecol. Sociobiol. 2022, 76, 19. [Google Scholar] [CrossRef]
- Mancini, G.; Ferrari, P.F.; Palagi, E. Rapid facial mimicry in geladas. Sci. Rep. 2013, 3, 1527. [Google Scholar] [CrossRef]
- Palagi, E.; Marchi, E.; Cavicchio, P.; Bandoli, F. Sharing playful mood: Rapid facial mimicry in Suricata suricatta. Anim. Cogn. 2019, 22, 719–732. [Google Scholar] [CrossRef]
- Kret, M.E.; Akyüz, R. Mimicry eases prediction and thereby smoothens social interactions. Cogn. Emot. 2022, 36, 794–798. [Google Scholar] [CrossRef]
- Norscia, I.; Caselli, M.; Scianna, C.; Morone, S.; Brescini, M.; Cordoni, G. Is it a match? Yawn contagion and smile mimicry in toddlers. Hum. Nat. 2024; under review. [Google Scholar]
- Cordoni, G. Do you copy? Trunk mimicry during play in African Savanna elephants. Sci. Rep. 2024; preprint. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cordoni, G.; Norscia, I. Nuancing ‘Emotional’ Social Play: Does Play Behaviour Always Underlie a Positive Emotional State? Animals 2024, 14, 2769. https://doi.org/10.3390/ani14192769
Cordoni G, Norscia I. Nuancing ‘Emotional’ Social Play: Does Play Behaviour Always Underlie a Positive Emotional State? Animals. 2024; 14(19):2769. https://doi.org/10.3390/ani14192769
Chicago/Turabian StyleCordoni, Giada, and Ivan Norscia. 2024. "Nuancing ‘Emotional’ Social Play: Does Play Behaviour Always Underlie a Positive Emotional State?" Animals 14, no. 19: 2769. https://doi.org/10.3390/ani14192769
APA StyleCordoni, G., & Norscia, I. (2024). Nuancing ‘Emotional’ Social Play: Does Play Behaviour Always Underlie a Positive Emotional State? Animals, 14(19), 2769. https://doi.org/10.3390/ani14192769