
Location, Age, and Antibodies Predict Avian Influenza Virus Shedding in Ring-Billed and Franklin’s Gulls in Minnesota

 ,
,  and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Gull Sampling
2.2. Influenza A Virus RNA and Antibody Detection
2.3. Generalized Linear Models
3. Results
3.1. Sampling Distribution
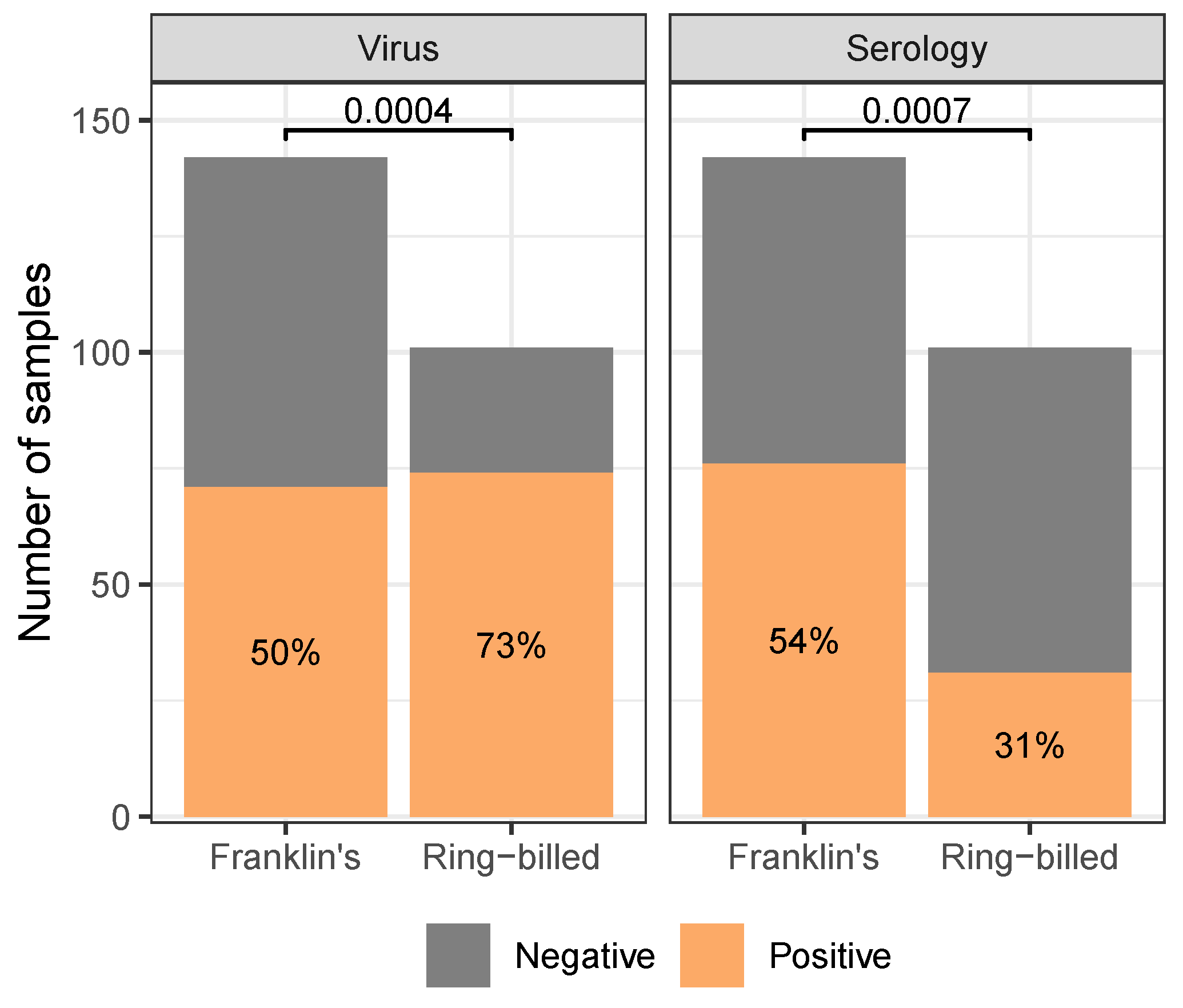
3.2. Observational Results
3.3. Model Results
4. Discussion
4.1. Serological Sampling Can Complement Current Surveillance
4.2. The Role of the Surrounding Environment
4.3. Insights into Influenza Reservoirs
4.4. Limitations of BCI in Cross-Species Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| IAV | Influenza A Virus |
| RNA | Ribonucleic acid |
| PCR CT | Polymerase Chain Reaction cycle threshold |
| ELISA S/N | Enzyme-linked Immunosorbent Assay sample result-to-negative control absorbance ratio |
| HPAI | High Pathogenic Avian Influenza |
| LPAI | Low Pathogenic Avian Influenza |
| BIC | Body Condition Index |
References
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Emerging Infectious Diseases of Wildlife—Threats to Biodiversity and Human Health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.J.; Albery, G.F.; Merow, C.; Trisos, C.H.; Zipfel, C.M.; Eskew, E.A.; Olival, K.J.; Ross, N.; Bansal, S. Climate change increases cross-species viral transmission risk. Nature 2022, 607, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Meadows, A.J.; Stephenson, N.; Madhav, N.K.; Oppenheim, B. Historical trends demonstrate a pattern of increasingly frequent and severe spillover events of high-consequence zoonotic viruses. BMJ Glob. Health 2023, 8, e012026. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.F.; Acevedo-Whitehouse, K.; Pedersen, A.B. The role of infectious diseases in biological conservation. Anim. Conserv. 2009, 12, 1–12. [Google Scholar] [CrossRef]
- Musoke, J.; Hlokwe, T.; Marcotty, T.; du Plessis, B.J.; Michel, A.L. Spillover of Mycobacterium bovis from Wildlife to Livestock, South Africa. Emerg. Infect. Dis. 2015, 21, 448–451. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Miller, R.S.; Farnsworth, M.L.; Malmberg, J.L. Diseases at the livestock-wildlife interface: Status, challenges, and opportunities in the United States. Prev. Vet. Med. 2013, 110, 119–132. [Google Scholar] [CrossRef]
- Ruiz-Aravena, M.; McKee, C.; Gamble, A.; Lunn, T.; Morris, A.; Snedden, C.E.; Yinda, C.K.; Port, J.R.; Buchholz, D.W.; Yeo, Y.Y.; et al. Ecology, evolution and spillover of coronaviruses from bats. Nat. Rev. Microbiol. 2022, 20, 299–314. [Google Scholar] [CrossRef]
- Plowright, R.K.; Parrish, C.R.; McCallum, H.; Hudson, P.J.; Ko, A.I.; Graham, A.L.; Lloyd-Smith, J.O. Pathways to zoonotic spillover. Nat. Rev. Microbiol. 2017, 15, 502–510. [Google Scholar] [CrossRef]
- Viana, M.; Mancy, R.; Biek, R.; Cleaveland, S.; Cross, P.C.; Lloyd-Smith, J.O.; Haydon, D.T. Assembling evidence for identifying reservoirs of infection. Trends Ecol. Evol. 2014, 29, 270–279. [Google Scholar] [CrossRef]
- Haydon, D.T.; Cleaveland, S.; Taylor, L.; Laurenson, M.K. Identifying Reservoirs of Infection: A Conceptual and Practical Challenge. Emerg. Infect. Dis. 2002, 8, 1468–1473. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wilber, M.Q.; Yang, A.; Boughton, R.; Manlove, K.R.; Miller, R.S.; Pepin, K.M.; Wittemyer, G. A model for leveraging animal movement to understand spatio-temporal disease dynamics. Ecol. Lett. 2022, 25, 1290–1304. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.K.; Craft, M.E.; Zuk, M.; Binning, S.A. Host migration strategy is shaped by forms of parasite transmission and infection cost. J. Anim. Ecol. 2019, 88, 1601–1612. [Google Scholar] [CrossRef]
- Taubenberger, J.K.; Morens, D.M. The 1918 Influenza Pandemic and Its Legacy. Cold Spring Harb. Perspect. Med. 2020, 10, a038695. [Google Scholar] [CrossRef] [PubMed]
- Bourret, V. Avian influenza viruses in pigs: An overview. Vet. J. 2018, 239, 7–14. [Google Scholar] [CrossRef]
- Loeb, J. Scottish seabirds hit by avian influenza. Vet. Rec. 2022, 190, 488. [Google Scholar] [CrossRef]
- Garber, L.; Bjork, K.; Patyk, K.; Rawdon, T.; Antognoli, M.; Delgado, A.; Ahola, S.; McCluskey, B. Factors Associated with Highly Pathogenic Avian Influenza H5N2 Infection on Table-Egg Layer Farms in the Midwestern United States, 2015. Avian Dis. 2016, 60, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Ssematimba, A.; Malladi, S.; Hagenaars, T.J.; Bonney, P.J.; Weaver, J.T.; Patyk, K.A.; Spackman, E.; Halvorson, D.A.; Cardona, C.J. Estimating within-flock transmission rate parameter for H5N2 highly pathogenic avian influenza virus in Minnesota turkey flocks during the 2015 epizootic. Epidemiol. Infect. 2019, 147, e179. [Google Scholar] [CrossRef] [PubMed]
- Spackman, E. A Brief Introduction to Avian Influenza Virus. In Animal Influenza Virus: Methods and Protocols; Spackman, E., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2020; pp. 83–92. [Google Scholar] [CrossRef]
- van Dijk, J.G.B.; Hoye, B.J.; Verhagen, J.H.; Nolet, B.A.; Fouchier, R.A.M.; Klaassen, M. Juveniles and migrants as drivers for seasonal epizootics of avian influenza virus. J. Anim. Ecol. 2014, 83, 266–275. [Google Scholar] [CrossRef]
- Elbers, A.R.W.; Gonzales, J.L. Quantification of visits of wild fauna to a commercial free-range layer farm in the Netherlands located in an avian influenza hot-spot area assessed by video-camera monitoring. Transbound. Emerg. Dis. 2020, 67, 661–677. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Chen, H.; Yuan, H.; Gao, R.; Zhang, J.; Wang, D.; Xiong, Y.; Fan, G.; Yang, F.; Li, X.; Zhou, J.; et al. Clinical and epidemiological characteristics of a fatal case of avian influenza A H10N8 virus infection: A descriptive study. Lancet 2014, 383, 714–721. [Google Scholar] [CrossRef] [PubMed]
- Reid, A.H.; Taubenberger, J.K. The origin of the 1918 pandemic influenza virus: A continuing enigma. J. Gen. Virol. 2003, 84, 2285–2292. [Google Scholar] [CrossRef] [PubMed]
- Arnal, A.; Vittecoq, M.; Pearce-Duvet, J.; Gauthier-Clerc, M.; Boulinier, T.; Jourdain, E. Laridae: A neglected reservoir that could play a major role in avian influenza virus epidemiological dynamics. Crit. Rev. Microbiol. 2015, 41, 508–519. [Google Scholar] [CrossRef]
- Postnikova, Y.; Treshchalina, A.; Boravleva, E.; Gambaryan, A.; Ishmukhametov, A.; Matrosovich, M.; Fouchier, R.A.M.; Sadykova, G.; Prilipov, A.; Lomakina, N. Diversity and Reassortment Rate of Influenza A Viruses in Wild Ducks and Gulls. Viruses 2021, 13, 1010. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ineson, K.M.; Hill, N.J.; Clark, D.E.; MacKenzie, K.G.; Whitney, J.J.; Laskaris, Y.; Ronconi, R.A.; Ellis, J.C.; Giroux, J.F.; Lair, S.; et al. Age and season predict influenza A virus dynamics in urban gulls: Consequences for natural hosts in unnatural landscapes. Ecol. Appl. 2022, 32, e2497. [Google Scholar] [CrossRef] [PubMed]
- Hill, N.J.; Takekawa, J.Y.; Cardona, C.J.; Meixell, B.W.; Ackerman, J.T.; Runstadler, J.A.; Boyce, W.M. Cross-Seasonal Patterns of Avian Influenza Virus in Breeding and Wintering Migratory Birds: A Flyway Perspective. Vector-Borne Zoonotic Dis. 2012, 12, 243–253. [Google Scholar] [CrossRef]
- Anderson, C.M.; Gilchrist, H.G.; Ronconi, R.A.; Shlepr, K.R.; Clark, D.E.; Fifield, D.A.; Robertson, G.J.; Mallory, M.L. Both short and long distance migrants use energy-minimizing migration strategies in North American herring gulls. Mov. Ecol. 2020, 8, 26. [Google Scholar] [CrossRef]
- Rasmussen, E.A.; Czaja, A.; Cuthbert, F.J.; Tan, G.S.; Lemey, P.; Nelson, M.I.; Culhane, M.R. Influenza A viruses in gulls in landfills and freshwater habitats in Minnesota, United States. Front. Genet. 2023, 14, 1172048. [Google Scholar] [CrossRef]
- Arriero, E.; Müller, I.; Juvaste, R.; Martínez, F.J.; Bertolero, A. Variation in Immune Parameters and Disease Prevalence among Lesser Black-Backed Gulls (Larus fuscus sp.) with Different Migratory Strategies. PLoS ONE 2015, 10, e0118279. [Google Scholar] [CrossRef]
- Dusek, R.J.; Hallgrimsson, G.T.; Ip, H.S.; Jónsson, J.E.; Sreevatsan, S.; Nashold, S.W.; TeSlaa, J.L.; Enomoto, S.; Halpin, R.A.; Lin, X.; et al. North Atlantic Migratory Bird Flyways Provide Routes for Intercontinental Movement of Avian Influenza Viruses. PLoS ONE 2014, 9, e92075. [Google Scholar] [CrossRef]
- Huang, Y.; Wille, M.; Benkaroun, J.; Munro, H.; Bond, A.L.; Fifield, D.A.; Robertson, G.J.; Ojkic, D.; Whitney, H.; Lang, A.S. Perpetuation and reassortment of gull influenza A viruses in Atlantic North America. Virology 2014, 456–457, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Guinn, K.; Fojtik, A.; Davis-Fields, N.; Poulson, R.L.; Krauss, S.; Webster, R.G.; Stallknecht, D.E. Antibodies to Influenza A Viruses in Gulls at Delaware Bay, USA. Avian Dis. 2016, 60, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Reeves, A.B.; Hall, J.S.; Poulson, R.L.; Donnelly, T.; Stallknecht, D.E.; Ramey, A.M. Influenza A virus recovery, diversity, and intercontinental exchange: A multi-year assessment of wild bird sampling at Izembek National Wildlife Refuge, Alaska. PLoS ONE 2018, 13, e0195327. [Google Scholar] [CrossRef]
- Bevins, S.N.; Pedersen, K.; Lutman, M.W.; Baroch, J.A.; Schmit, B.S.; Kohler, D.; Gidlewski, T.; Nolte, D.L.; Swafford, S.R.; DeLiberto, T.J. Large-Scale Avian Influenza Surveillance in Wild Birds throughout the United States. PLoS ONE 2014, 9, e104360. [Google Scholar] [CrossRef] [PubMed]
- Krauss, S.; Stucker, K.M.; Schobel, S.A.; Danner, A.; Friedman, K.; Knowles, J.P.; Kayali, G.; Niles, L.J.; Dey, A.D.; Raven, G.; et al. Long-term surveillance of H7 influenza viruses in American wild aquatic birds: Are the H7N3 influenza viruses in wild birds the precursors of highly pathogenic strains in domestic poultry? Emerg. Microbes Infect. 2015, 4, 1–9. [Google Scholar] [CrossRef]
- Criado, M.F.; Moresco, K.A.; Stallknecht, D.E.; Swayne, D.E. Low-pathogenicity influenza viruses replicate differently in laughing gulls and mallards. Influenza Other Respir. Viruses 2021, 15, 701–706. [Google Scholar] [CrossRef]
- USDA-APHIS. Epidemiologic and other Analyses of HPAI Affected Poultry Flocks: July 1, 2022 Interim Report; Technical report; USDA:APHIS:VS: Center for Epidemiology and Animal Health: Fort Collins, CO, USA, 2022.
- Froberg, T.; Cuthbert, F.; Jennelle, C.S.; Cardona, C.; Culhane, M. Avian Influenza Prevalence and Viral Shedding Routes in Minnesota Ring-Billed Gulls (Larus delawarensis). Avian Dis. 2018, 63, 120–125. [Google Scholar] [CrossRef]
- Howell, S.N.G.; Dunn, J. A Reference Guide to Gulls of the Americas; Houghton Mifflin Harcourt: Boston, MA, USA, 2007. [Google Scholar]
- Pyle, P. Identification Guide to North American Birds. Part II: Anatidae to Alcidae; Slate Creek Press: Point Reyes Station, CA, USA, 2008. [Google Scholar]
- Pollet, I.L.; Shutler, D.; Chardine, J.W.; Ryder, J.P. Ring-billed Gull (Larus delawarensis), version 2.0. In The Birds of North America; Cornell Lab of Ornithology: Ithaca, NY, USA, 2012. [Google Scholar] [CrossRef]
- Velarde, R.; Calvin, S.E.; Ojkic, D.; Barker, I.K.; Nagy, E. Avian Influenza Virus H13 Circulating in Ring-Billed Gulls (Larus delawarensis) in Southern Ontario, Canada. Avian Dis. 2010, 54, 411–419. [Google Scholar] [CrossRef]
- Boersma, D.; Ryder, J.P. Reproductive Performance and Body Condition of Earlier and Later Nesting Ring-Billed Gulls. J. Field Ornithol. 1983, 54, 374–380. [Google Scholar]
- Kelly, L.M.; Alworth, L.C. Techniques for collecting blood from the domestic chicken. Lab Anim. 2013, 42, 359–361. [Google Scholar] [CrossRef]
- Owen, J.C. Collecting, processing, and storing avian blood: A review. J. Field Ornithol. 2011, 82, 339–354. [Google Scholar] [CrossRef]
- Spackman, E.; Suarez, D.L. Type A Influenza Virus Detection and Quantitation by Real-Time RT-PCR. In Avian Influenza Virus; Spackman, E., Ed.; Methods in Molecular BiologyTM; Humana Press: Totowa, NJ, USA, 2008; pp. 19–26. [Google Scholar] [CrossRef]
- Munster, V.J.; Baas, C.; Lexmond, P.; Bestebroer, T.M.; Guldemeester, J.; Beyer, W.E.; de Wit, E.; Schutten, M.; Rimmelzwaan, G.F.; Osterhaus, A.D.; et al. Practical considerations for high-throughput influenza A virus surveillance studies of wild birds by use of molecular diagnostic tests. J. Clin. Microbiol. 2009, 47, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.D.; Stallknecht, D.E.; Berghaus, R.D.; Luttrell, M.P.; Velek, K.; Kistler, W.; Costa, T.; Yabsley, M.J.; Swayne, D. Evaluation of a Commercial Blocking Enzyme-Linked Immunosorbent Assay To Detect Avian Influenza Virus Antibodies in Multiple Experimentally Infected Avian Species. Clin. Vaccine Immunol. 2009, 16, 824–829. [Google Scholar] [CrossRef] [PubMed]
- Minnesota Department of Natural Resources. National Wetland Inventory for Minnesota. 2019. Available online: https://gisdata.mn.gov/dataset/water-nat-wetlands-inv-2009-2014 (accessed on 29 August 2022).
- McKelvey, R.D.; Zavoina, W. A statistical model for the analysis of ordinal level dependent variables. J. Math. Sociol. 1975, 4, 103–120. [Google Scholar] [CrossRef]
- Veall, M.R.; Zimmermann, K.F. Evaluating Pseudo-R2’s for binary probit models. Qual. Quant. 1994, 28, 151–164. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024. [Google Scholar]
- Robinson, D.; Hayes, A.; Couch, S. broom: Convert Statistical Objects into Tidy Tibbles; R Package Version 1.0.6. 2024. Available online: https://CRAN.R-project.org/package=broom (accessed on 29 August 2022).
- Robinson, D. fuzzyjoin: Join Tables Together on Inexact Matching; R Package Version 0.1.6. 2020. Available online: https://CRAN.R-project.org/package=fuzzyjoin (accessed on 29 August 2022).
- Schloerke, B.; Cook, D.; Larmarange, J.; Briatte, F.; Marbach, M.; Thoen, E.; Elberg, A.; Crowley, J. GGally: Extension to ’ggplot2’; R Package Version 2.2.1. 2024. Available online: https://CRAN.R-project.org/package=GGally (accessed on 29 August 2022).
- Clarke, E.; Sherrill-Mix, S.; Dawson, C. ggbeeswarm: Categorical Scatter (Violin Point) Plots; R Package Version 0.7.2. 2023. Available online: https://cran.r-project.org/package=ggbeeswarm (accessed on 29 August 2022).
- Kahle, D.; Wickham, H. ggmap: Spatial Visualization with ggplot2. R J. 2013, 5, 144–161. [Google Scholar] [CrossRef]
- Kassambara, A. ggpubr: ’ggplot2’ Based Publication Ready Plots; R Package Version 0.6.0. 2023. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 29 August 2022).
- Rizopoulos, D. GLMMadaptive: Generalized Linear Mixed Models Using Adaptive Gaussian Quadrature; R Package Version 0.9-1. 2023. Available online: https://cloud.r-project.org/web/packages/GLMMadaptive/index.html (accessed on 29 August 2022).
- Firke, S. janitor: Simple Tools for Examining and Cleaning Dirty Data; R Package Version 2.2.0. 2023. Available online: https://CRAN.R-project.org/package=janitor (accessed on 29 August 2022).
- Zhu, H. kableExtra: Construct Complex Table with ’kable’ and Pipe Syntax; R Package Version 1.4.0. 2024. Available online: https://CRAN.R-project.org/package=kableExtra (accessed on 29 August 2022).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Grolemund, G.; Wickham, H. Dates and Times Made Easy with lubridate. J. Stat. Softw. 2011, 40, 1–25. [Google Scholar] [CrossRef]
- Bache, S.M.; Wickham, H. magrittr: A Forward-Pipe Operator for R; R Package Version 2.0.3. 2022. Available online: https://CRAN.R-project.org/package=magrittr (accessed on 29 August 2022).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Pedersen, T.L. patchwork: The Composer of Plots; R Package Version 1.2.0. 2024. Available online: https://CRAN.R-project.org/package=patchwork (accessed on 29 August 2022).
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.C.; Müller, M. pROC: An open-source package for R and S+ to analyze and compare ROC curves. BMC Bioinform. 2011, 12, 77. [Google Scholar] [CrossRef]
- Yu, G. scatterpie: Scatter Pie Plot; R Package Version 0.2.3. 2024. Available online: https://CRAN.R-project.org/package=scatterpie (accessed on 29 August 2022).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Michel, A.L.; Van Heerden, H.; Crossley, B.M.; Al Dahouk, S.; Prasse, D.; Rutten, V. Pathogen detection and disease diagnosis in wildlife: Challenges and opportunities. Rev. Sci. Tech. (Int. Off. Epizoot.) 2021, 40, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.D.; Luttrell, M.P.; Berghaus, R.D.; Kistler, W.; Keeler, S.P.; Howey, A.; Wilcox, B.; Hall, J.; Niles, L.; Dey, A.; et al. Prevalence of Antibodies to Type A Influenza Virus in Wild Avian Species Using Two Serologic Assays. J. Wildl. Dis. 2010, 46, 896–911. [Google Scholar] [CrossRef]
- Gilbert, A.T.; Fooks, A.R.; Hayman, D.T.S.; Horton, D.L.; Müller, T.; Plowright, R.; Peel, A.J.; Bowen, R.; Wood, J.L.N.; Mills, J.; et al. Deciphering Serology to Understand the Ecology of Infectious Diseases in Wildlife. EcoHealth 2013, 10, 298–313. [Google Scholar] [CrossRef]
- Salewski, V.; Bruderer, B. The evolution of bird migration—A synthesis. Naturwissenschaften 2007, 94, 268–279. [Google Scholar] [CrossRef]
- Johnson, J.A.; DeCicco, L.H.; Ruthrauff, D.R.; Krauss, S.; Hall, J.S. Avian Influenza Virus Antibodies in Pacific Coast Red Knots (Calidris canutus roselaari). J. Wildl. Dis. 2014, 50, 671–675. [Google Scholar] [CrossRef]
- Verhagen, J.H.; Munster, V.J.; Majoor, F.; Lexmond, P.; Vuong, O.; Stumpel, J.B.G.; Rimmelzwaan, G.F.; Osterhaus, A.D.M.E.; Schutten, M.; Slaterus, R.; et al. Avian Influenza A Virus in Wild Birds in Highly Urbanized Areas. PLoS ONE 2012, 7, e38256. [Google Scholar] [CrossRef]
- Wilson, H.M.; Hall, J.S.; Flint, P.L.; Franson, J.C.; Ely, C.R.; Schmutz, J.A.; Samuel, M.D. High Seroprevalence of Antibodies to Avian Influenza Viruses among Wild Waterfowl in Alaska: Implications for Surveillance. PLoS ONE 2013, 8, e58308. [Google Scholar] [CrossRef]
- Lu, L.; Lycett, S.J.; Leigh Brown, A.J. Reassortment patterns of avian influenza virus internal segments among different subtypes. BMC Evol. Biol. 2014, 14, 16. [Google Scholar] [CrossRef] [PubMed]
- Morse, S.S. Public Health Surveillance and Infectious Disease Detection. Biosecurity Bioterrorism Biodefense Strateg. Pract. Sci. 2012, 10, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Fuller, T.L.; Gilbert, M.; Martin, V.; Cappelle, J.; Hosseini, P.; Njabo, K.Y.; Abdel Aziz, S.; Xiao, X.; Daszak, P.; Smith, T.B. Predicting Hotspots for Influenza Virus Reassortment. Emerg. Infect. Dis. 2013, 19, 581–588. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Fereidouni, S. Ecology of Avian Influenza Viruses in Wild Birds. In Ecology of Wild Bird Diseases; CRC Press: Boca Raton, FL, USA, 2024; p. 18. [Google Scholar]
- Ljubojević, D.; Radosavljević, V.; Milanov, D. The role of gulls (Laridae) in the emergence and spreading of antibiotic resistance in the environment. World’S Poult. Sci. J. 2016, 72, 853–864. [Google Scholar] [CrossRef]
- Endo, A.; Nishiura, H. The Role of Migration in Maintaining the Transmission of Avian Influenza in Waterfowl: A Multisite Multispecies Transmission Model along East Asian-Australian Flyway. Can. J. Infect. Dis. Med. Microbiol. 2018, 2018, 3420535. [Google Scholar] [CrossRef]
- Raven, S.; Coulson, J. The distribution and abundance of Larus gulls nesting on buildings in Britain and Ireland. Bird Study 1997, 44, 13–34. [Google Scholar] [CrossRef]
- Zhang, Y.; Aevermann, B.D.; Anderson, T.K.; Burke, D.F.; Dauphin, G.; Gu, Z.; He, S.; Kumar, S.; Larsen, C.N.; Lee, A.J.; et al. Influenza Research Database: An integrated bioinformatics resource for influenza virus research. Nucleic Acids Res. 2017, 45, D466–D474. [Google Scholar] [CrossRef]
- Hatch, J.J. Threats to public health from gulls (Laridae). Int. J. Environ. Health Res. 1996, 6, 5–16. [Google Scholar] [CrossRef]
- Ahlstrom, C.A.; van Toor, M.L.; Woksepp, H.; Chandler, J.C.; Reed, J.A.; Reeves, A.B.; Waldenström, J.; Franklin, A.B.; Douglas, D.C.; Bonnedahl, J.; et al. Evidence for continental-scale dispersal of antimicrobial resistant bacteria by landfill-foraging gulls. Sci. Total Environ. 2021, 764, 144551. [Google Scholar] [CrossRef]
- Bevins, S.N.; Shriner, S.A.; Cumbee, J.C.; Dilione, K.E.; Douglass, K.E.; Ellis, J.W.; Killian, M.L.; Torchetti, M.K.; Lenoch, J.B. Intercontinental Movement of Highly Pathogenic Avian Influenza A(H5N1) Clade 2.3.4.4 Virus to the United States, 2021. Emerg. Infect. Dis. 2022, 28, 1006–1011. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Taylor, L.U.; Ronconi, R.A.; Spina, H.A.; Jones, M.E.; Ogbunugafor, C.B.; Ayala, A.J. Limited Outbreak of Highly Pathogenic Influenza A (H5N1) in Herring Gull Colony, Canada, 2022. Emerg. Infect. Dis. 2023, 29, 2150. [Google Scholar] [CrossRef]
- Ramis, A.; Van Amerongen, G.; Van de Bildt, M.; Leijten, L.; Vanderstichel, R.; Osterhaus, A.; Kuiken, T. Experimental infection of highly pathogenic avian influenza virus H5N1 in black-headed gulls (Chroicocephalus ridibundus). Vet. Res. 2014, 45, 84. [Google Scholar] [CrossRef]
- Brown, J.D.; Stallknecht, D.E.; Beck, J.R.; Suarez, D.L.; Swayne, D.E. Susceptibility of North American ducks and gulls to H5N1 highly pathogenic avian influenza viruses. Emerg. Infect. Dis. 2006, 12, 1663. [Google Scholar] [CrossRef] [PubMed]
- Verhagen, J.H.; Fouchier, R.A.; Lewis, N. Highly pathogenic avian influenza viruses at the wild–domestic bird interface in Europe: Future directions for research and surveillance. Viruses 2021, 13, 212. [Google Scholar] [CrossRef] [PubMed]
- USDA-APHIS. Detections of Highly Pathogenic Avian Influenza in Wild Birds; USDA-APHIS: Riverdale Park, MD, USA, 2024.
- Labocha, M.K.; Hayes, J.P. Morphometric indices of body condition in birds: A review. J. Ornithol. 2012, 153, 1–22. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus Name | Subtype | Pathogenicity | Abbreviation | Lab Designator |
|---|---|---|---|---|
| A/Chicken/Mexico/1433-2/2008 | H5N2 | Low | MX/08/H5N2/LP | none |
| A/Turkey/Wisconsin/1968 | H5N9 | Low | WI/68/H5N9/LP | none |
| A/Turkey/Oregon/1971 | H7N3 | Low | OR/71/H7N3/LP | none |
| A/Turkey/Minnesota/9845-4/2015 | H5N2 | High | MN/15/H5N2/HP | 133 ADV 1501 |
| 1/4/2005 to 7/3/2013 | 7/4/2013 to 12/31/2021 | |||||
|---|---|---|---|---|---|---|
| Positive | Tested | Positivity | Positive | Tested | Positivity | |
| Anatidae | 14,624 | 164,215 | 8.9% | 29,584 | 258,631 | 11.4% |
| Laridae | 1396 | 28,225 | 4.9% | 3206 | 31,134 | 10.3% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michalska-Smith, M.; Clements, E.; Rasmussen, E.; Culhane, M.R.; Craft, M.E. Location, Age, and Antibodies Predict Avian Influenza Virus Shedding in Ring-Billed and Franklin’s Gulls in Minnesota. Animals 2024, 14, 2781. https://doi.org/10.3390/ani14192781
Michalska-Smith M, Clements E, Rasmussen E, Culhane MR, Craft ME. Location, Age, and Antibodies Predict Avian Influenza Virus Shedding in Ring-Billed and Franklin’s Gulls in Minnesota. Animals. 2024; 14(19):2781. https://doi.org/10.3390/ani14192781
Chicago/Turabian StyleMichalska-Smith, Matthew, Eva Clements, Elizabeth Rasmussen, Marie R. Culhane, and Meggan E. Craft. 2024. "Location, Age, and Antibodies Predict Avian Influenza Virus Shedding in Ring-Billed and Franklin’s Gulls in Minnesota" Animals 14, no. 19: 2781. https://doi.org/10.3390/ani14192781
APA StyleMichalska-Smith, M., Clements, E., Rasmussen, E., Culhane, M. R., & Craft, M. E. (2024). Location, Age, and Antibodies Predict Avian Influenza Virus Shedding in Ring-Billed and Franklin’s Gulls in Minnesota. Animals, 14(19), 2781. https://doi.org/10.3390/ani14192781