

Behavioral Coding of Captive African Elephants (Loxodonta africana): Utilizing DeepLabCut and Create ML for Nocturnal Activity Tracking
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Objectives of Wildlife Conservation
1.2. Captive Elephant Behavior and Welfare
1.3. Machine Learning as a Tool for Behavioral Analysis
1.4. Aim of This Paper
- This study expects that the machine learning models can predict selected behaviors on the same level as manual scoring;
- This study expects that behavioral differences between the elephants and behavioral differences between days can be demonstrated using selected computer vision models.
2. Materials and Methods
2.1. Subjects and Enclosure
2.2. Data Collection
2.3. Data Analysis
| Algorithm 1. DeepLabCut operation |
| Input: Video containing the subject to be tracked |
| 1. Load dataset. Load the video into DeepLabCut; |
| 2. Define keypoints. Specify keypoints of interest (body parts like head, tail, limbs); |
| 3. Annotate frames. Annotate a subset of frames manually by marking the keypoints; |
| 4. Model training. Use annotated frames to train the pose estimation model; |
| 5. Pose estimation. Apply the trained model to new video material; |
| 6. Refine model (optional). Correct predictions and retrain the model; |
| 7. Process coordinates. Extract CSV file and filter the coordinates for desired body parts; |
| 8. Classify behaviors. Set limits for each coordinate corresponding to a desired behavior and filter frames that fulfill the criteria. |
| Output: Subset of data points that can be classified as a specific behavior. |
| Algorithm 2. Create ML operation |
| Input: Video containing the subject to be tracked. |
| 1. Load dataset. Load the video into RectLabel and extract images; |
| 2. Define bounding boxes. Specify categories of each bounding box of interest (behaviors, such as lying down or standing); |
| 3. Annotate frames. Annotate a subset of frames manually by drawing bounding boxes; |
| 4. Model training. Use annotated frames to train the model; |
| 5. Pose estimation. Apply the trained model to new video material; |
| 6. Refine model (optional). Correct predictions and retrain the model; |
| 7. Process coordinates. Extract CSV file containing frames annotated with behaviors. |
| Output: Dataset containing the predicted behaviors at all analyzed frames. |
3. Results
3.1. Comparability of Manual and Automatic Behavioural Observations
3.1.1. A General Overview
3.1.2. Investigating the Reliability of Two Machine Learning Models
3.2. Using Machine Learning Models for Behavioural Analysis
3.2.1. Assessing Behavioral Differences
3.2.2. Investigating Further Applications of Automatic Behavioral Coding
4. Discussion
4.1. Performance and Limitations of the Two Machine Learning Models
4.2. Nocturnal Behavioral Differences of the Two Subjects
4.3. Other Applications of Machine Learning Models for Behavioral Coding
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Floor Plan of Elephant Enclosure
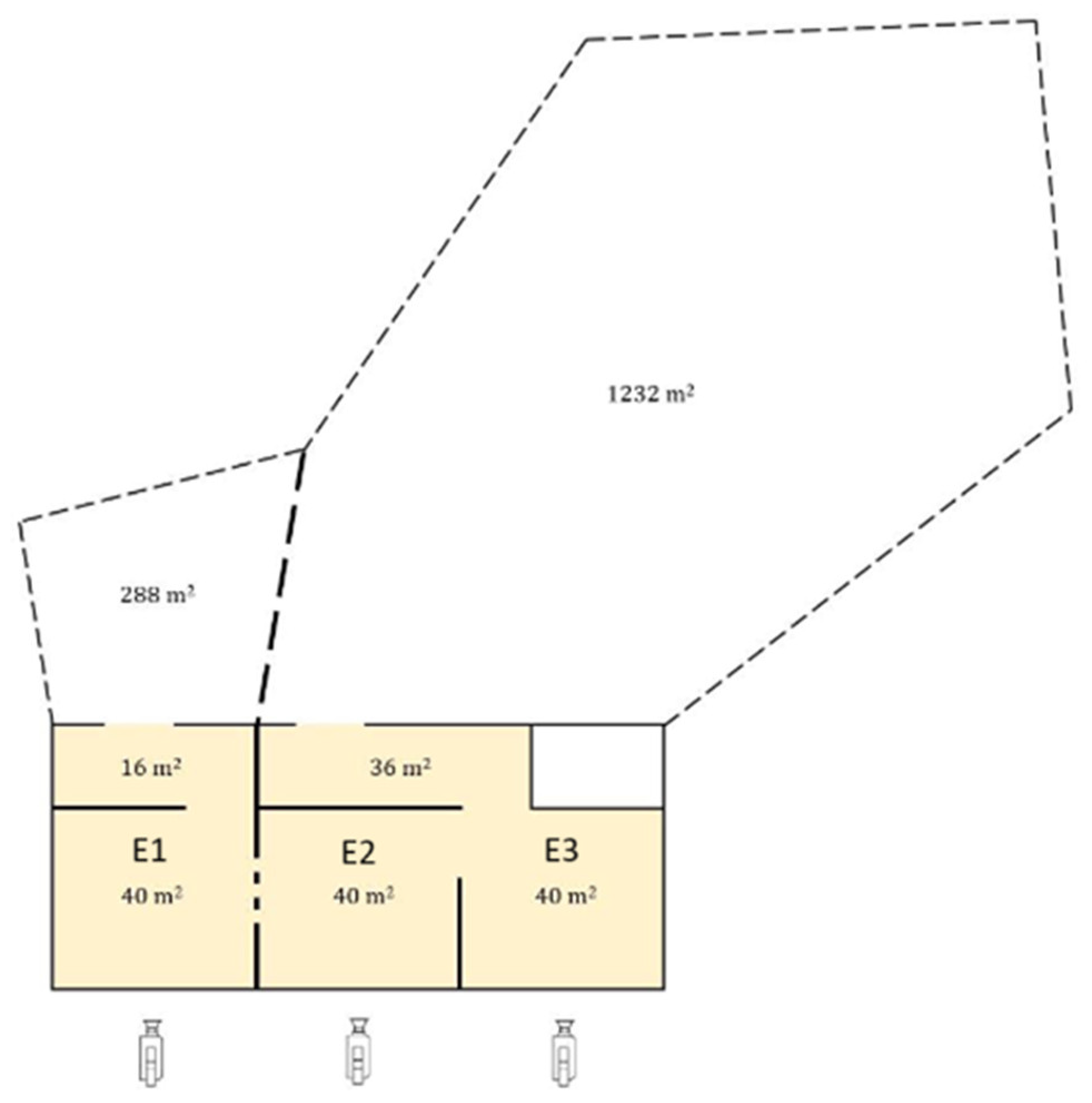
Appendix B. Selected Behaviors Displayed in Subject Enclosures


Appendix C. Confusion Matrix Comparison of Models

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject A | Manually Observed | ||||||
|---|---|---|---|---|---|---|---|
| Predicted by DeepLabCut model | Behavior: | Standing | Lying down | Foraging | Drinking | Hay-net | Out of view |
| Standing | 0.86820 | 0.00016 | 0.13258 | 1.00000 | 0.39926 | ||
| Lying down | 0.00036 | 0.89418 | |||||
| Foraging | 0.00701 | 0.84651 | |||||
| Drinking | |||||||
| Hay-net | 0.00005 | 0.16883 | |||||
| Out of view | 0.12437 | 0.10567 | 0.02091 | 0.43190 | |||
| Subject B | Manually Observed | ||||||
|---|---|---|---|---|---|---|---|
| Predicted by DeepLabCut model | Behavior: | Standing | Lying down | Foraging | Drinking | Hay-net | Out of view |
| Standing | 0.64320 | 0.00170 | 0.10185 | 0.76200 | 0.58339 | 0.18203 | |
| Lying down | 0.02517 | 0.99657 | 0.06867 | 0.00271 | |||
| Foraging | 0.02384 | 0.66607 | 0.14721 | ||||
| Drinking | |||||||
| Hay-net | 0.00016 | ||||||
| Out of view | 0.30763 | 0.00173 | 0.23209 | 0.16933 | 0.26669 | 0.81797 | |
| Subject A | Manually Observed | ||||||
|---|---|---|---|---|---|---|---|
| Predicted by Create ML model | Behavior: | Standing | Lying down | Foraging | Drinking | Hay-net | Out of view |
| Standing | 0.94253 | 0.29061 | 0.84503 | 0.07534 | |||
| Lying down | 0.00051 | 0.88842 | |||||
| Foraging | 0.00718 | 0.53114 | |||||
| Drinking | 0.00038 | 0.07602 | |||||
| Hay-net | 0.00051 | 0.91096 | |||||
| Out of view | 0.04887 | 0.11158 | 0.17825 | 0.07895 | 0.01370 | ||
| Subject B | Manually Observed | ||||||
|---|---|---|---|---|---|---|---|
| Predicted by Create ML model | Behavior: | Standing | Lying down | Foraging | Drinking | Hay-net | Out of view |
| Standing | 0.91524 | 0.00121 | 0.04310 | 0.36667 | 0.06907 | 0.29412 | |
| Lying down | 0.99868 | ||||||
| Foraging | 0.05509 | 0.94939 | 0.50390 | ||||
| Drinking | 0.00890 | 0.00011 | 0.61667 | ||||
| Hay-net | 0.00130 | 0.01667 | 0.38559 | ||||
| Out of view | 0.01948 | 0.00751 | 0.04144 | 0.70588 | |||
Appendix D. Time Budgets for Each Night


Appendix E. Cumulative Graphs for Every Behavior Each Night

Appendix F. Spearman Rank Correlation between Days
| 16-04-2024 | 17-04-2024 | 18-04-2024 | 19-04-2024 | 20-04-2024 | 21-04-2024 | 22-04-2024 | |
|---|---|---|---|---|---|---|---|
| 16-04-2024 | 1.000 | ||||||
| 17-04-2024 | 0.820 | 1.000 | |||||
| 18-04-2024 | 0.756 | 0.965 | 1.000 | ||||
| 19-04-2024 | 0.465 | 0.779 | 0.791 | 1.000 | |||
| 20-04-2024 | 0.367 | 0.307 | 0.237 | 0.207 | 1.000 | ||
| 21-04-2024 | 0.716 | 0.938 | 0.924 | 0.700 | 0.276 | 1.000 | |
| 22-04-2024 | 0.786 | 0.612 | 0.520 | 0.250 | –0.053 | 0.450 | 1.000 |
| 16-04-2024 | 17-04-2024 | 18-04-2024 | 19-04-2024 | 20-04-2024 | 21-04-2024 | 22-04-2024 | |
|---|---|---|---|---|---|---|---|
| 16-04-2024 | 1.000 | ||||||
| 17-04-2024 | 0.930 | 1.000 | |||||
| 18-04-2024 | 0.860 | 0.777 | 1.000 | ||||
| 19-04-2024 | 0.299 | 0.115 | 0.573 | 1.000 | |||
| 20-04-2024 | 0.297 | 0.401 | 0.357 | 0.019 | 1.000 | ||
| 21-04-2024 | 0.997 | 0.924 | 0.872 | 0.315 | 0.314 | 1.000 | |
| 22-04-2024 | 0.843 | 0.760 | 0.999 | 0.601 | 0.372 | 0.856 | 1.000 |
| 16-04-2024 | 17-04-2024 | 18-04-2024 | 19-04-2024 | 20-04-2024 | 21-04-2024 | 22-04-2024 | |
|---|---|---|---|---|---|---|---|
| 16-04-2024 | 1.000 | ||||||
| 17-04-2024 | 0.902 | 1.000 | |||||
| 18-04-2024 | 0.872 | 0.918 | 1.000 | ||||
| 19-04-2024 | 0.759 | 0.841 | 0.859 | 1.000 | |||
| 20-04-2024 | 0.222 | 0.043 | 0.102 | –0.135 | 1.000 | ||
| 21-04-2024 | 0.896 | 0.934 | 0.970 | 0.855 | 0.112 | 1.000 | |
| 22-04-2024 | 0.737 | 0.741 | 0.501 | 0.473 | 0.048 | 0.516 | 1.000 |
| 16-04-2024 | 17-04-2024 | 18-04-2024 | 19-04-2024 | 20-04-2024 | 21-04-2024 | 22-04-2024 | |
|---|---|---|---|---|---|---|---|
| 16-04-2024 | 1.000 | ||||||
| 17-04-2024 | 0.933 | 1.000 | |||||
| 18-04-2024 | 0.913 | 0.842 | 1.000 | ||||
| 19-04-2024 | 0.747 | 0.662 | 0.882 | 1.000 | |||
| 20-04-2024 | 0.311 | 0.414 | 0.379 | 0.558 | 1.000 | ||
| 21-04-2024 | 1.000 | 0.933 | 0.917 | 0.752 | 0.313 | 1.000 | |
| 22-04-2024 | 0.852 | 0.775 | 0.988 | 0.892 | 0.373 | 0.857 | 1.000 |
Appendix G. Spearman Rank Correlation between Individuals
| Subject B | ||||||||
|---|---|---|---|---|---|---|---|---|
| Subject A | 16-04-2024 | 17-04-2024 | 18-04-2024 | 19-04-2024 | 20-04-2024 | 21-04-2024 | 22-04-2024 | |
| 16-04-2024 | 0.476 | |||||||
| 17-04-2024 | 0.772 | 0.700 | ||||||
| 18-04-2024 | 0.839 | 0.762 | 0.975 | |||||
| 19-04-2024 | 0.884 | 0.790 | 0.783 | 0.407 | ||||
| 20-04-2024 | 0.372 | 0.508 | 0.194 | –0.361 | 0.717 | |||
| 21-04-2024 | 0.654 | 0.622 | 0.881 | 0.551 | 0.672 | 0.672 | ||
| 22-04-2024 | 0.203 | 0.068 | 0.441 | 0.879 | 0.197 | 0.224 | 0.474 | |
| Subject B | ||||||||
|---|---|---|---|---|---|---|---|---|
| Subject A | 16-04-2024 | 17-04-2024 | 18-04-2024 | 19-04-2024 | 20-04-2024 | 21-04-2024 | 22-04-2024 | |
| 16-04-2024 | 0.690 | |||||||
| 17-04-2024 | 0.673 | 0.563 | ||||||
| 18-04-2024 | 0.855 | 0.785 | 0.977 | |||||
| 19-04-2024 | 0.532 | 0.470 | 0.790 | 0.920 | ||||
| 20-04-2024 | 0.206 | 0.314 | 0.112 | 0.052 | 0.602 | |||
| 21-04-2024 | 0.743 | 0.711 | 0.918 | 0.874 | 0.491 | 0.748 | ||
| 22-04-2024 | 0.347 | 0.194 | 0.419 | 0.512 | 0.066 | 0.349 | 0.472 | |
References
- Veasey, J.S. Differing animal welfare conceptions and what they mean for the future of zoos and aquariums, insights from an animal welfare audit. Zoo Biol. 2022, 41, 292–307. [Google Scholar] [CrossRef] [PubMed]
- Barongi, R.; Fisken, F.A.; Parker, M.; Gusset, M. Committing to Conservation: The World Zoo and Aquarium Conservation Strategy; World Association of Zoos and Aquariums (WAZA) Executive Office: Gland, Switzerland, 2015. [Google Scholar]
- European Association of Zoos and Aquaria. Standards for the Accommodation and Care of Animals in Zoos and Aquaria; EAZA: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Danish Association of Zoos and Aquaria. DAZA Etiske Retningslinjer; DAZA: Aalborg, Denmark, 2022; Available online: https://www.daza.dk/pdf/DAZA%20etiske%20retningslinjer%20-%20godkendt%20GF%2015.06.2022-underskrevet.pdf (accessed on 9 April 2023).
- Sutherland, W.J. The importance of behavioural studies in conservation biology. Anim. Behav. 1998, 56, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Wolfensohn, S.; Shotton, J.; Bowley, H.; Davies, S.; Thompson, S.; Justice, W.S.M. Assessment of Welfare in Zoo Animals: Towards Optimum Quality of Life. Animals 2018, 8, 110. [Google Scholar] [CrossRef] [PubMed]
- Perdue, B.M.; Sherwen, S.L.; Maple, T.L. Editorial: The Science and Practice of Captive Animal Welfare. Front. Psychol. 2020, 11, 1851. [Google Scholar] [CrossRef] [PubMed]
- Sherwen, S.L.; Hemsworth, P.H. The Visitor Effect on Zoo Animals: Implications and Opportunities for Zoo Animal Welfare. Animals 2019, 9, 366. [Google Scholar] [CrossRef] [PubMed]
- Hauenstein, S.; Jassoy, N.; Mupepele, A.; Carroll, T.; Kshatriya, M.; Beale, C.M.; Dormann, C.F. A systematic map of demographic data from elephant populations throughout Africa: Implications for poaching and population analyses. Mammal Rev. 2022, 52, 438–453. [Google Scholar] [CrossRef]
- Rees, A.P. Activity budgets and the relationship between feeding and stereotypic behaviors in Asian elephants (Elephas maximus) in a Zoo. Zoo Biol. 2009, 28, 79–97. [Google Scholar] [CrossRef] [PubMed]
- Yon, L.; Williams, E.; Harvey, N.D.; Asher, L. Development of a behavioural welfare assessment tool for routine use with captive elephants. PLoS ONE 2019, 14, e0210783. [Google Scholar] [CrossRef] [PubMed]
- Greco, B.J.; Meehan, C.L.; Heinsius, J.L.; Mench, J.A. Why pace? The influence of social, housing, management, life history, and demographic characteristics on locomotor stereotypy in zoo elephants. Appl. Anim. Behav. Sci. 2017, 194, 104–111. [Google Scholar] [CrossRef]
- Bansiddhi, P.; Nganvongpanit, K.; Brown, J.L.; Punyapornwithaya, V.; Pongsopawijit, P.; Thitaram, C. Management factors affecting physical health and welfare of tourist camp elephants in Thailand. Biodivers. Conserv. 2019, 7, e6756. [Google Scholar] [CrossRef]
- Bertelsen, S.S.; Sørensen, A.S.; Pagh, S.; Pertoldi, C.; Jensen, T.H. Nocturnal Behaviour of Three Zoo Elephants (Loxodonta africana). Genet. Biodivers. J. 2020, 4, 92–113. [Google Scholar] [CrossRef]
- Holdgate, M.R.; Meehan, C.L.; Hogan, J.N.; Miller, L.J.; Rushen, J.; de Passillé, A.M.; Soltis, J.; Andrews, J.; Shepherdson, D.J. Recumbence Behavior in Zoo Elephants: Determination of Patterns and Frequency of Recumbent Rest and Associated Environmental and Social Factors. PLoS ONE 2016, 11, e0153301. [Google Scholar] [CrossRef] [PubMed]
- Boyle, S.A.; Roberts, B.; Pope, B.M.; Blake, M.R.; Leavelle, S.R.; Marshall, J.J.; Smith, A.; Hadicke, A.; Falcone, J.F.; Knott, K.; et al. Assessment of Flooring Renovations on African Elephant (Loxodonta africana) Behavior and Glucocorticoid Response. PLoS ONE 2015, 10, e0141009. [Google Scholar] [CrossRef]
- Schiffmann, C.; Hellriegel, L.; Clauss, M.; Stefan, B.; Knibbs, K.; Wenker, C.; Hård, T.; Galeffi, C. From left to right all through the night: Characteristics of lying rest in zoo elephants. Zoo Biol. 2022, 42, 17–25. [Google Scholar] [CrossRef]
- Greco, B.J.; Meehan, C.L.; Hogan, J.N.; Leighty, K.A.; Mellen, J.; Mason, G.J.; Mench, J.A. The Days and Nights of Zoo Elephants: Using Epidemiology to Better Understand Stereotypic Behvaior of African Elephants (Loxodonta africana) and Asian Elephants (Elephas maximus) in North American Zoos. PLoS ONE 2016, 11, e0144276. [Google Scholar] [CrossRef]
- Broom, D.M. Welfare of Animals: Behavior as a Basis for Decisions. Encycl. Anim. Behav. 2010, 3, 580–584. [Google Scholar]
- Jacobson, S.L.; Ross, S.R.; Bloomsmith, M.A. Characterizing abnormal behavior in a large population of zoo-housed chimpanzees: Prevalence and potential influencing factors. PeerJ 2016, 4, e2225. [Google Scholar] [CrossRef] [PubMed]
- Bacon, H. Behaviour-Based Husbandry—A Holistic Approach to the Management of Abnormal Repetitive Behaviors. Animals 2018, 8, 103. [Google Scholar] [CrossRef]
- Fuktong, S.; Yuttasaen, P.; Punyapornwithaya, V.; Brown, J.L.; Thitaram, C.; Luevitoonvechakij, N.; Bansiddhi, P. A survey of stereotypic behaviors in tourist camp elephants in Chiang Mai, Thailand. Appl. Anim. Behav. Sci. 2021, 243, 105456. [Google Scholar] [CrossRef]
- Mason, G.J. Stereotypies and suffering. Behav. Process. 1991, 25, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Mason, G.J.; Rushen, J. Stereotypic Animal Behaviour: Fundamentals and Applications to Welfare; CABI Digital Library: Wallingford, UK, 2006. [Google Scholar]
- Mostard, K.E. General Understanding, Neuro-Endocrinologic and (Epi)Genetic Factors of Stereotypy; Radboud University of Nijmegen: Nijmegen, The Netherlands, 2011. [Google Scholar]
- Altmann, J. Observational Study of Behavior: Sampling Methods. Behaviour 1974, 49, 227–267. [Google Scholar] [CrossRef]
- Vanitha, V.; Thiyagesan, K.; Baskaran, N. Prevalence of stereotypies and its possible causes among captive Asian elephants (Elephas maximus) in Tamil Nadu, India. Appl. Anim. Behav. Sci. 2016, 174, 137–146. [Google Scholar]
- Adams, J.; Berg, J.K. Behavior of Female African Elephants (Loxodonta africana) in Captivity. Appl. Anim. Ethol. 1980, 6, 257–276. [Google Scholar] [CrossRef]
- Dell, A.I.; Bender, J.A.; Branson, K.; Couzin, I.D.; de Polavieja, G.G.; Noldus, L.P.; Pérez-Escudero, A.; Perona, P.; Straw, A.D.; Wikelski, M.; et al. Automated image-based tracking and its application in ecology. Trends Ecol. Evol. 2014, 29, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Marin, A.; Paton, J.J.; Kampff, A.R.; Costa, R.M.; Mainen, Z.F. Big Behavioral Data: Psychology, Ethology and the Foundations of Neuroscience. Nat. Neurosci. 2014, 17, 1455–1462. [Google Scholar] [CrossRef]
- Anderson, D.J.; Perona, P. Toward a Science of Computational Ethology. Neuron 2014, 84, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Mathis, A.; Mamidanna, P.; Cury, K.M.; Abe, T.; Murthy, V.N.; Mathis, M.W.; Bethge, M. DeepLabCut: Markerless pose estimation of user-defined body parts with deep learning. Nat. Neurosci. 2018, 21, 1281–1289. [Google Scholar] [CrossRef]
- Mirkó, E.; Dóka, A.; Miklósi, Á. Association between subjective rating and behaviour coding and the role of experience in making video assessments on the personality of the domestic dog (Canis familiaris). Appl. Anim. Behav. Sci. 2013, 149, 45–54. [Google Scholar] [CrossRef]
- Zhao, Z.-Q.; Zheng, P.; Xu, S.-T.; Wu, X. Object Detection With Deep Learning: A Review. IEEE Trans. Neural Netw. Learn. Syst. 2019, 30, 3212–3232. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.-H.; Liu, S. Machine Learning; Springer Nature: Nanjing, China, 2021. [Google Scholar]
- Lenzi, J.; Barnas, A.F.; ElSaid, A.A.; Desell, T.; Rockwell, R.F.; Ellis-Felege, S.N. Artificial intelligence for automated detection of large mammals creates path to upscale drone surveys. Sci. Rep. 2023, 13, 947. [Google Scholar] [CrossRef]
- Bain, M.; Nagrani, A.; Schofield, D.; Berdugo, S.; Bessa, J.; Owen, J.; Hockings, K.J.; Matsuzawa, T.; Hayashi, M.; Biro, D.; et al. Automated audiovisual behavior recognition in wild primates. Sci. Adv. 2021, 7, eabi4883. [Google Scholar] [CrossRef] [PubMed]
- Sakib, F.; Burghardt, T. Visual Recognition of Great Ape Behaviours in the Wild. arXiv 2020, arXiv:2011.10759. [Google Scholar]
- Hebbar, P.K.; Pullela, P.K. Deep Learning in Object Detection: Advancements in Machine Learning and AI. In Proceedings of the 2023 International Conference on the Confluence of Advancements in Robotics, Vision and Interdisciplinary Technology Management (IC-RVITM), Bangalore, India, 28–29 November 2023. [Google Scholar]
- Hardin, A.; Schlupp, I. Using machine learning and DeepLabCut in animal behavior. Acta Ethologica 2022, 25, 125–133. [Google Scholar] [CrossRef]
- Marks, M.; Jin, Q.; Sturman, O.; von Ziegler, L.; Kollmorgen, S.; von der Behrens, W.; Mante, V.; Bohacek, J.; Yanik, M.F. Deep-learning-based identification, tracking, pose estimation and behaviour classification of interacting primates and mice in complex environments. Nat. Mach. Intell. 2022, 4, 331–340. [Google Scholar] [CrossRef]
- Nath, T.; Mathis, A.; Chen, A.C.; Patel, A.; Bethge, M.; Mathis, M.W. Using DeepLabCut for 3D markerless pose estimation across species and behaviors. Nat. Protoc. 2019, 14, 2152–2176. [Google Scholar] [CrossRef] [PubMed]
- Marques, O. Machine Learning with Core ML. In Image Processing and Computer Vision in IOS; Springer International Publishing AG: Cham, Switzerland, 2020; pp. 29–40. [Google Scholar]
- Andersen, T.A.; Herskind, C.; Maysfelt, J.; Rørbæk, R.W.; Schnoor, C.; Pertoldi, C. The nocturnal behaviour of African elephants (Loxodonta africana) in Aalborg Zoo and how changes in the environment affect them. Genet. Biodivers. J. 2020, 4, 114–130. [Google Scholar] [CrossRef]
- Ruuska, S.; Hämäläinen, W.; Kajava, S.; Mughal, M.; Matilainen, P.; Mononen, J. Evaluation of the confusion matrix method in the validation of an automated system for measuring feeding behaviour of cattle. Behav. Process. 2018, 148, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Larsen, J.F.; Andersen, K.K.D.; Cuprys, J.; Fosgaard, T.B.; Jacobsen, J.H.; Krysztofiak, D.; Lund, S.M.; Nielsen, B.; Pedersen, M.E.B.; Pedersen, M.J.; et al. Behavioral analysis of a captive male Bornean orangutan (Pongo pygmaeus) when. Arch. Biol. Sci. 2023, 75, 443–458. [Google Scholar] [CrossRef]
- Field, A.P. Kendall’s Coefficient of Concordance. Encycl. Stat. Behav. Sci. 2005, 2, 1010–1011. [Google Scholar]
- Brownlee, J. Machine Learning Mastery. Guiding Tech Media. 19 August 2020. Available online: https://machinelearningmastery.com/distance-measures-for-machine-learning/ (accessed on 22 May 2024).
- Malviya, M.; Buswell, N.T.; Berdanier, C.G.P. Visual and Statistical Methods to Calculate Intercoder Reliability for Time-Resolved Observational Research. Int. J. Qual. Methods 2021, 20, 16094069211002418. [Google Scholar] [CrossRef]
- Whiteway, M.R.; Biderman, D.; Friedman, Y.; Dipoppa, M.; Buchanan, E.K.; Wu, A.; Zhou, J.; Bonacchi, N.; Miska, N.J.; Noel, J.-P.; et al. Partitioning variability in animal behavioral videos using semi-supervised variational autoencoders. PLoS Comput. Biol. 2021, 17, e1009439. [Google Scholar] [CrossRef] [PubMed]
- García-Mateos, G.; Hernández-Hernández, J.; Escarabajal-Henarejos, D.; Jaén-Terrones, S.; Molina-Martínez, J. Study and comparison of color models for automatic image analysis in irrigation management applications. Agric. Water Manag. 2015, 151, 158–166. [Google Scholar] [CrossRef]
- Hernández-Hernández, J.L.; García-Mateos, G.; González-Esquiva, J.M.; Escarabajal-Henarejos, D.; Ruiz-Canales, A.; Molina-Martínez, J.M. Optimal color space selection method for plant/soil segmentation in agriculture. Comput. Electron. Agric. 2016, 122, 124–132. [Google Scholar] [CrossRef]
- Tobler, I. Fundamental Research Behavioral Sleep in the Asian Elephant in Captivity. Sleep 1992, 15, 1–12. [Google Scholar] [PubMed]
- Finch, K.; Sach, F.; Fitzpatrick, M.; Rowden, L.J. Insights into Activity of Zoo Housed Asian Elephants (Elephas maximus) during Periods of Limited Staff and Visitor Presence, a Focus on Resting Behaviour. Zool. Bot. Gard. 2021, 2, 101–114. [Google Scholar] [CrossRef]
- Casares, M.; Silván, G.; Carbonell, M.D.; Gerique, C.; Martinez-Fernandez, L.; Cáceres, S.; Illera, J.C. Circadian rhythm of salivary cortisol secretion in female zoo-kept African elephants (Loxodonta africana). Zoo Biol. 2016, 35, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Zhao, Y.; Xu, Z.; Yu, Q. Automatic detection of stereotypical behaviors of captive wild animals based on surveillance videos of zoos and animal reserves. Ecol. Inform. 2024, 79, 102450. [Google Scholar] [CrossRef]




| Behavior | Description |
|---|---|
| Standing | The elephant is standing or walking. This behavior is the default if no other selected behavior is taking place. |
| Lying down | The elephant is lying down on the floor of the enclosure. |
| Drinking | The elephant is drinking from a water bowl. |
| Foraging | The elephant is using the foraging boxes, accessed using trunks in the holes at the back of the enclosure. |
| Hay-net | The elephant is using its trunk to reach the hay-net at the top of the enclosure. |
| Swaying | The elephant is swaying from side to side for at least 5 s. |
| Out of view | The elephant is out of view of the camera. This may also include falsely unlabeled frames by the machine learning models. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lund, S.M.; Nielsen, J.; Gammelgård, F.; Nielsen, M.G.; Jensen, T.H.; Pertoldi, C. Behavioral Coding of Captive African Elephants (Loxodonta africana): Utilizing DeepLabCut and Create ML for Nocturnal Activity Tracking. Animals 2024, 14, 2820. https://doi.org/10.3390/ani14192820
Lund SM, Nielsen J, Gammelgård F, Nielsen MG, Jensen TH, Pertoldi C. Behavioral Coding of Captive African Elephants (Loxodonta africana): Utilizing DeepLabCut and Create ML for Nocturnal Activity Tracking. Animals. 2024; 14(19):2820. https://doi.org/10.3390/ani14192820
Chicago/Turabian StyleLund, Silje Marquardsen, Jonas Nielsen, Frej Gammelgård, Maria Gytkjær Nielsen, Trine Hammer Jensen, and Cino Pertoldi. 2024. "Behavioral Coding of Captive African Elephants (Loxodonta africana): Utilizing DeepLabCut and Create ML for Nocturnal Activity Tracking" Animals 14, no. 19: 2820. https://doi.org/10.3390/ani14192820