
Effect of Dietary Benzoic Acid Supplementation on Growth Performance, Rumen Fermentation, and Rumen Microbiota in Weaned Holstein Dairy Calves


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Diets, and Experimental Design
2.2. Sample Collection and Analysis
2.2.1. Body Weight and Feed Intake
2.2.2. Determination of Nutrient Levels in Diet
2.2.3. Rumen Fluid Sampling
2.2.4. Determination of VFAs
2.2.5. DNA Extraction, PCR Amplification, and 16S rRNA Gene Sequencing
2.2.6. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Rumen Fermentation Parameters
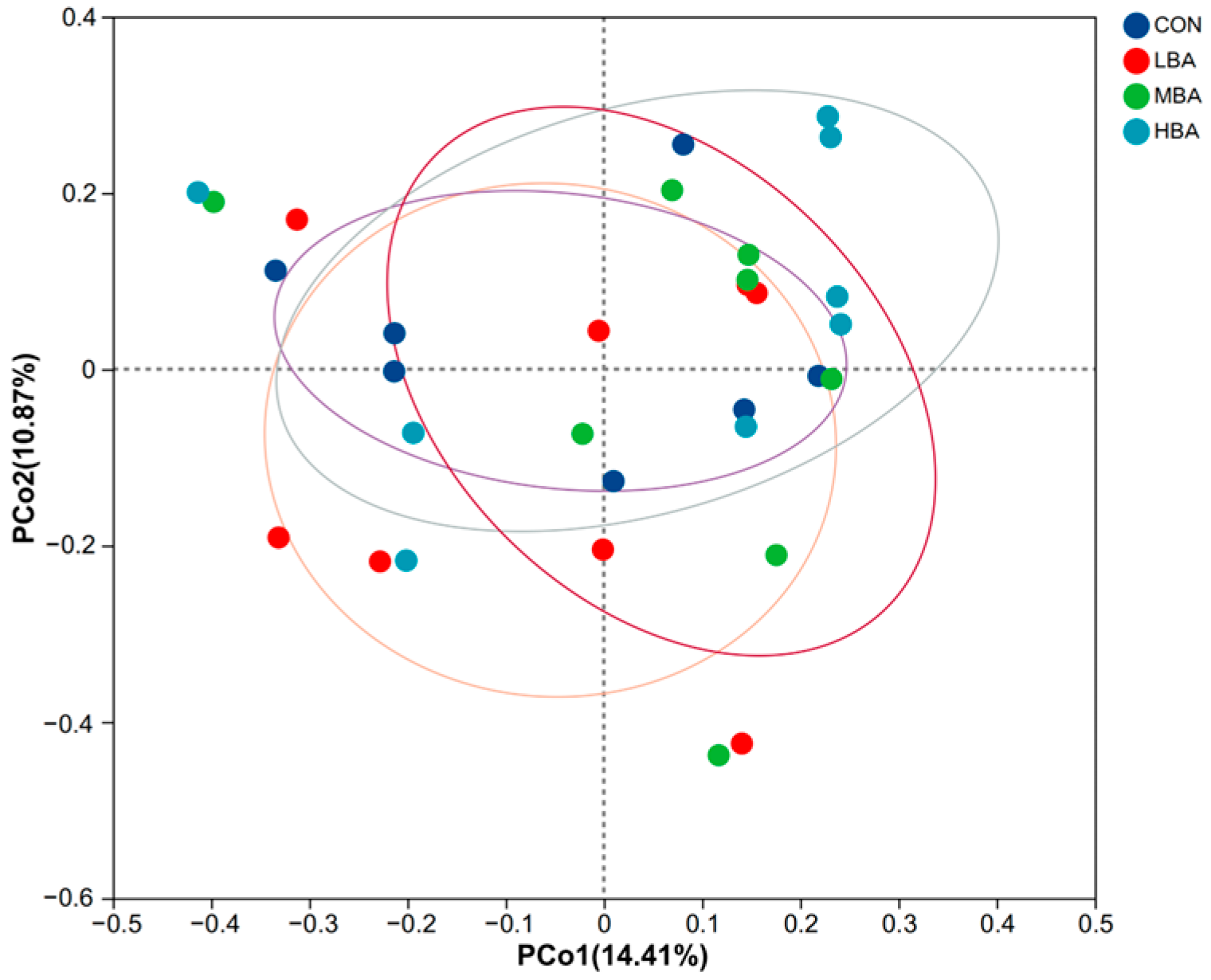
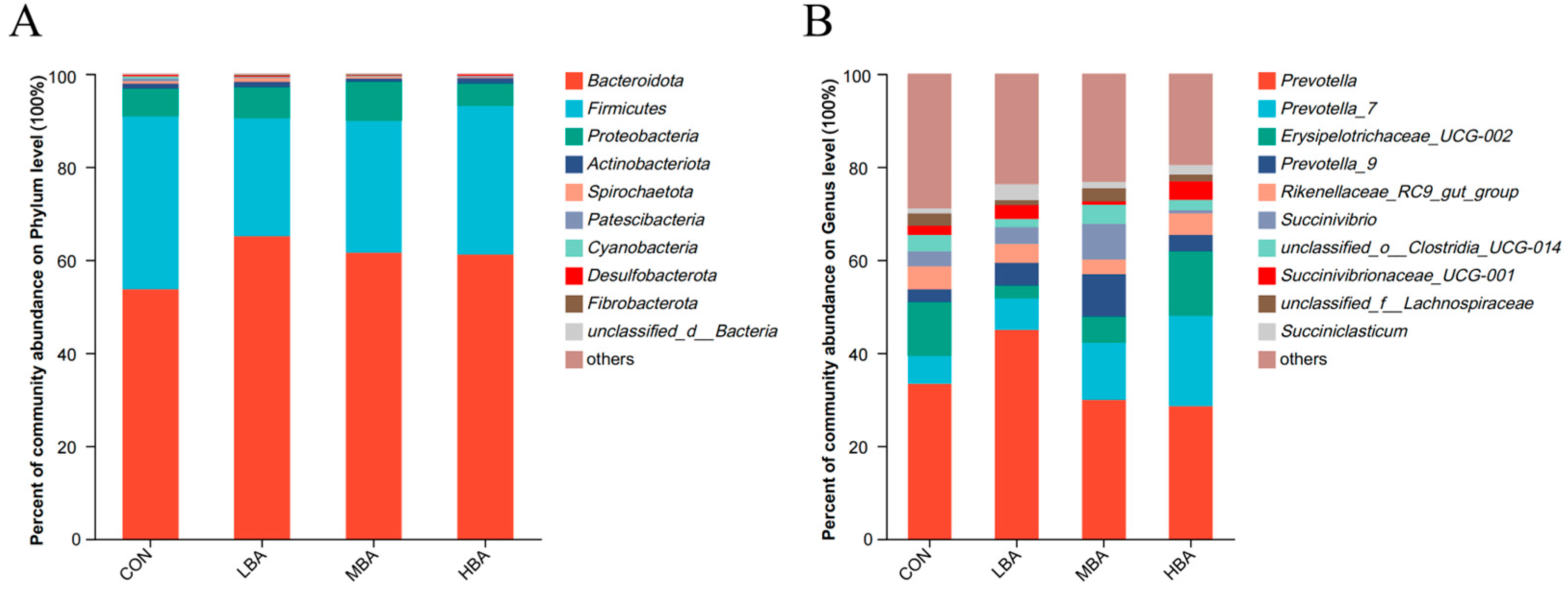
3.3. Composition of Rumen Microbiota
3.4. Comparison of Rumen Microbiota at the Genus Level
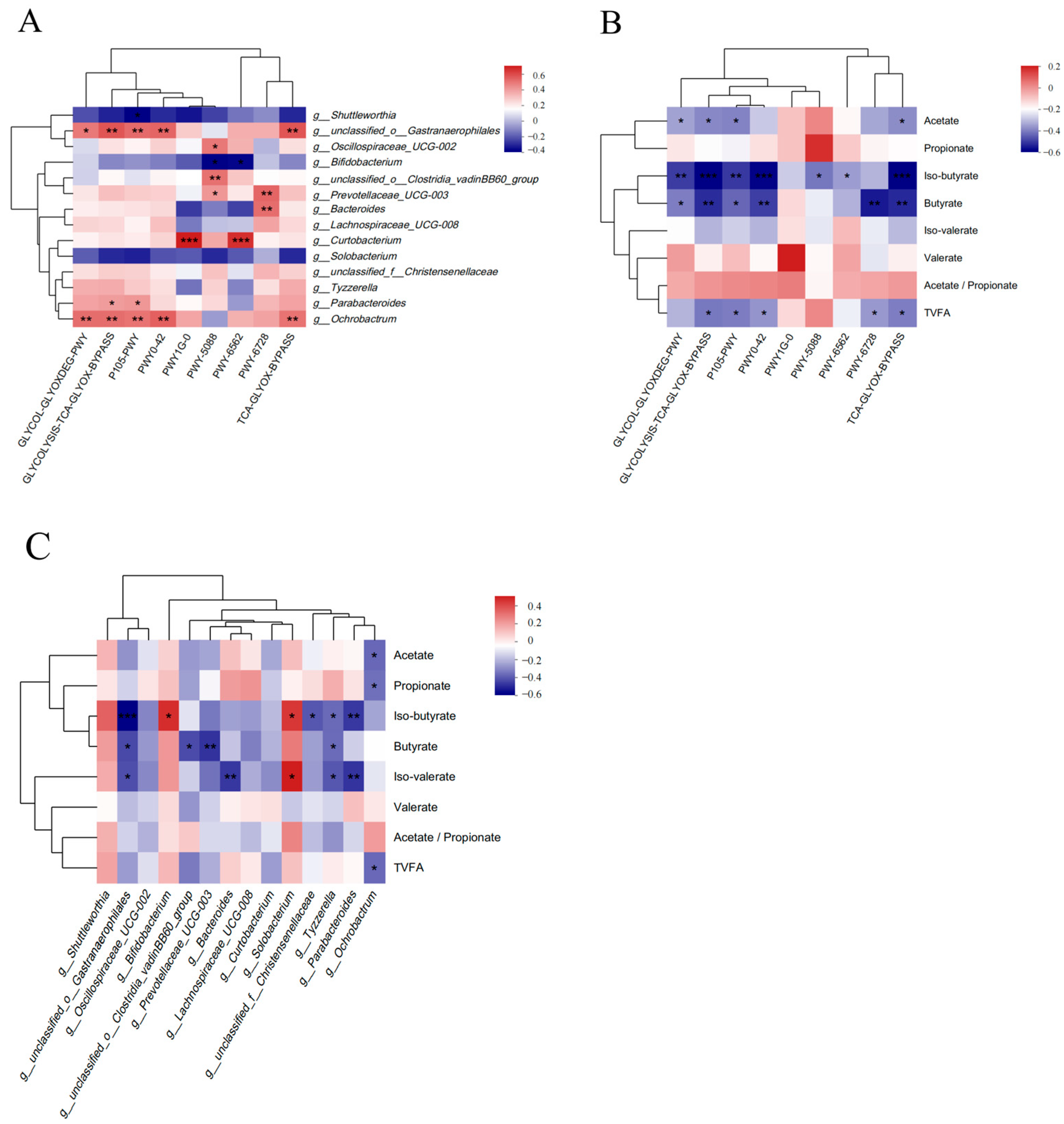
3.5. Functional Differences in Rumen Microbiota
3.6. Correlation Analysis of Differential Pathways, Differential Rumen Microbiota, and VFAs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wickramasinghe, J.; Kaya, C.A.; Beitz, D.; Appuhamy, R. Early Stepdown Weaning of Dairy Calves with Glutamine and Branched-Chain Amino Acid Supplementations. Animals 2022, 12, 1474. [Google Scholar] [CrossRef] [PubMed]
- Wickramasinghe, H.; Kaya, C.A.; Baumgard, L.H.; Appuhamy, J. Early step-down weaning of dairy calves from a high milk volume with glutamine supplementation. J. Dairy Sci. 2022, 105, 1186–1198. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Shah, A.M.; Shao, Y.; Wang, Z.; Zou, H.; Kang, K. Dietary supplementation of yeast cell wall improves the gastrointestinal development of weaned calves. Animal Nutrition. 2020, 6, 507–512. [Google Scholar] [CrossRef]
- Wang, S.; Wang, F.; Kong, F.; Cao, Z.; Wang, W.; Yang, H.; Wang, Y.; Bi, Y.; Li, S. Effect of Supplementing Different Levels of L-Glutamine on Holstein Calves during Weaning. Antioxidants 2022, 11, 542. [Google Scholar] [CrossRef]
- Dai, D.; Qi, G.; Wang, J.; Zhang, H.; Qiu, K.; Han, Y.; Wu, Y.; Wu, S. Dietary organic acids ameliorate high stocking density stress-induced intestinal inflammation through the restoration of intestinal microbiota in broilers. J. Anim. Sci. Biotechnol. 2022, 13, 124. [Google Scholar] [CrossRef]
- Abu, E.N.; Ragaa, N.M. Eubiotic effect of a dietary acidifier (potassium diformate) on the health status of cultured Oreochromis niloticus. J. Adv. Res. 2015, 6, 621–629. [Google Scholar]
- Willing, B.P.; Pepin, D.M.; Marcolla, C.S.; Forgie, A.J.; Diether, N.E.; Bourrie, B.C.T. Bacterial resistance to antibiotic alternatives: A wolf in sheep’s clothing. Anim. Front. 2018, 8, 39–47. [Google Scholar] [CrossRef]
- Burgess, K.; Li, H.; Abo-Zeid, Y.; Fatimah; Williams, G.R. The Effect of Molecular Properties on Active Ingredient Release from Electrospun Eudragit Fibers. Pharmaceutics 2018, 10, 103. [Google Scholar] [CrossRef]
- Ivanovic, M.; Mirkovic, N.; Mirkovic, M.; Miocinovic, J.; Radulovic, A.; Solevic Knudsen, T.; Radulovic, Z. Autochthonous Enterococcus durans PFMI565 and Lactococcus lactis subsp. lactis BGBU1-4 in Bio-Control of Listeria monocytogenes in Ultrafiltered Cheese. Foods 2021, 10, 1448. [Google Scholar] [CrossRef]
- Zhang, Y.; Wei, J.; Qiu, Y.; Niu, C.; Song, Z.; Yuan, Y.; Yue, T. Structure-Dependent Inhibition of Stenotrophomonas maltophilia by Polyphenol and Its Impact on Cell Membrane. Front. Microbiol. 2019, 10, 2646. [Google Scholar] [CrossRef]
- Stratford, M.; Vallieres, C.; Geoghegan, I.A.; Archer, D.B.; Avery, S.V. The Preservative Sorbic Acid Targets Respiration, Explaining the Resistance of Fermentative Spoilage Yeast Species. mSphere 2020, 5, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.P.; Li, H.; Engmann, M.L.; Bischof, K.M.; Kunka, K.S.; Harris, M.E.; Tancredi, A.C.; Ditmars, F.S.; Basting, P.J.; George, N.S.; et al. Inverted Regulation of Multidrug Efflux Pumps, Acid Resistance, and Porins in Benzoate-Evolved Escherichia coli K-12. Appl. Environ. Microbiol. 2019, 85, e00966-19. [Google Scholar] [CrossRef] [PubMed]
- Song, J.H.; Kim, H.; Jeong, M.; Kong, M.J.; Choi, H.K.; Jun, W.; Kim, Y.; Choi, K.C. In Vivo Evaluation of Dendropanax morbifera Leaf Extract for Anti-Obesity and Cholesterol-Lowering Activity in Mice. Nutrients 2021, 13, 1424. [Google Scholar] [CrossRef] [PubMed]
- Yue, L.; Li, Y.; Zhong, M.; Chai, X.; Zhao, P.; Huang, R.; Kang, Y.; Yang, X. Benzoic Acid, Chlorine Dioxide, and 1-Methylcyclopropene Induce Flavonoid Metabolic Shifts in Postharvest Flowering Chinese Cabbage Revealed by High-Dimensional Analytical Data. Int. J. Mol. Sci. 2022, 23, 6011. [Google Scholar] [CrossRef]
- Zhai, H.; Ren, W.; Wang, S.; Wu, J.; Guggenbuhl, P.; Kluenter, A.M. Growth performance of nursery and grower-finisher pigs fed diets supplemented with benzoic acid. Anim. Nutr. 2017, 3, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Pu, J.; Chen, D.; Tian, G.; He, J.; Zheng, P.; Mao, X.; Yu, J.; Huang, Z.; Luo, J.; Luo, Y.; et al. Effects of benzoic acid, Bacillus coagulans and oregano oil combined supplementation on growth performance, immune status and intestinal barrier integrity of weaned piglets. Anim. Nutr. 2020, 6, 152–159. [Google Scholar] [CrossRef]
- Giannenas, I.; Papaneophytou, C.P.; Tsalie, E.; Tsalie, E.; Pappas, I.; Triantafillou, E.; Tontis, D.; Kontopidis, G.A. Dietary Supplementation of Benzoic Acid and Essential Oil Compounds Affects Buffering Capacity of the Feeds, Performance of Turkey Poults and Their Antioxidant Status, pH in the Digestive Tract, Intestinal Microbiota and Morphology. Asian-Australas. J. Anim. Sci. 2014, 27, 225–236. [Google Scholar] [CrossRef]
- Gong, H.; Yang, Z.; Celi, P.; Yan, L.; Ding, X.; Bai, S.; Zeng, Q.; Xu, S.; Su, Z.; Zhuo, Y.; et al. Effect of benzoic acid on production performance, egg quality, intestinal morphology, and cecal microbial community of laying hens. Poult. Sci. 2021, 100, 196–205. [Google Scholar] [CrossRef]
- Wang, L.M.; Huang, S.; Chalupa-Krebzdak, S.; Vásquez Mejía, S.M.; Mandell, I.B.; Bohrer, B.M. Effects of essential oils and(or) benzoic acid in beef finishing cattle diets on the fatty acid profile and shelflife stability of ribeye steaks and ground beef. Meat Sci. 2020, 168, 108195. [Google Scholar] [CrossRef]
- Williams, M.S.; Mandell, I.B.; Bohrer, B.M.; Wood, K.M. The effects of feeding benzoic acid and/or live active yeast (Saccharomyces cerevisiae) on beef cattle performance, feeding behavior, and carcass characteristics. Transl. Anim. Sci. 2021, 5, txab143. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Dairy Cattle; National Academy Press: Washington, DC, USA, 2021. [Google Scholar]
- AOAC International. Official Methods of Analysis, 18th ed.; AOAC International: Washington, DC, USA, 2005. [Google Scholar]
- AOAC International. Official Methods of Analysis, 17th ed.; AOAC International: Arlington, MA, USA, 2000. [Google Scholar]
- AOAC International. Official Methods of Analysis, 17th ed.; AOAC International: Gaithersburg, MD, USA, 2003. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis, 15th ed.; AOAC International: Arlington, MA, USA, 1990. [Google Scholar]
- Li, Z.; Wang, M.; Usman, S.; Khan, A.; Zhang, Y.; Li, F.; Bai, J.; Chen, M.; Zhang, Y.; Guo, X. Effects of nisin on bacterial community and fermentation profiles, in vitro rumen fermentation, microbiota, and methane emission of alfalfa silage. J. Sci. Food Agric. 2023, 103, 6706–6718. [Google Scholar] [CrossRef] [PubMed]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every base matter: Assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Nan, X.; Zhao, Y.; Jiang, L.; Wang, H.; Hua, D.; Zhang, F.; Wang, Y.; Liu, J.; Yao, J.; et al. Dietary supplementation with inulin improves lactation performance and serum lipids by regulating the rumen microbiome and metabolome in dairy cows. Anim. Nutr. 2021, 7, 1189–1204. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Xu, Q.; Wang, L.; Wang, J.; Guo, W.; Zhou, M. The impact of diet on the composition and relative abundance of rumen microbes in goat. Asian-Australas. J. Anim. Sci. 2017, 30, 531–537. [Google Scholar] [CrossRef]
- Yu, X.; Wo, Y.; Ma, F.; Shan, Q.; Wei, J.; Sun, P. Zinc methionine improves the lipid metabolism that is associated with the alteration of intestine mucosal proteomes and microbiota of newborn Holstein dairy calves. Anim. Res. One Health 2024, 2, 71–85. [Google Scholar] [CrossRef]
- Duthie, C.A.; Haskell, M.; Hyslop, J.J.; Waterhouse, A.; Wallace, R.J.; Roehe, R.; Rooke, J.A. The impact of divergent breed types and diets on methane emissions, rumen characteristics and performance of finishing beef cattle. Animal 2017, 11, 1762–1771. [Google Scholar] [CrossRef]
- Ma, Y.; Chen, X.; Khan, M.Z.; Xiao, J.; Cao, Z. A Combination of Novel Microecological Agents and Molasses Role in Digestibility and Fermentation of Rice Straw by Facilitating the Ruminal Microbial Colonization. Front. Microbiol. 2022, 13, 948049. [Google Scholar] [CrossRef]
- Ali, O.; Szabo-Fodor, J.; Febel, H.; Mézes, M.; Balogh, K.; Glávits, R.; Kovács, M.; Zantomasi, A.; Szabó, A. Porcine Hepatic Response to Fumonisin B(1) in a Short Exposure Period: Fatty Acid Profile and Clinical Investigations. Toxins 2019, 11, 655. [Google Scholar] [CrossRef]
- Roques, S.; Deborde, C.; Guimas, L.; Marchand, Y.; Richard, N.; Jacob, D.; Skiba-Cassy, S.; Moing, A.; Fauconneau, B. Integrative Metabolomics for Assessing the Effect of Insect (Hermetia illucens) Protein Extract on Rainbow Trout Metabolism. Metabolites 2020, 10, 83. [Google Scholar] [CrossRef]
- Seibt, K.D.; Ghaffari, M.H.; Scheu, T.; Koch, C.; Sauerwein, H. Characteristics of the Oxidative Status in Dairy Calves Fed at Different Milk Replacer Levels and Weaned at 14 Weeks of Age. Antioxidants 2021, 10, 260. [Google Scholar] [CrossRef] [PubMed]
- Diao, H.; Gao, Z.; Yu, B.; Zheng, P.; He, J.; Yu, J.; Huang, Z.; Chen, D.; Mao, X. Effects of benzoic acid (VevoVitall(R)) on the performance and jejunal digestive physiology in young pigs. J. Anim. Sci. Biotechnol. 2016, 7, 32. [Google Scholar] [CrossRef] [PubMed]
- Kluge, H.; Broz, J.; Eder, K. Effect of benzoic acid on growth performance, nutrient digestibility, nitrogen balance, gastrointestinal microflora and parameters of microbial metabolism in piglets. J. Anim. Physiol. Anim. Nutr. 2006, 90, 316–324. [Google Scholar] [CrossRef]
- Jozefiak, D.; Kaczmarek, S.; Rutkowski, A. The effects of benzoic acid supplementation on the performance of broiler chickens. J. Anim. Physiol. Anim. Nutr. 2010, 94, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Chen, X.; Tu, Y.; Wang, B.; Lou, C.; Ma, T.; Diao, Q. Effects of mulberry leaf flavonoid and resveratrol on methane emission and nutrient digestion in sheep. Anim. Nutr. 2015, 1, 362–367. [Google Scholar] [CrossRef]
- Bergman, E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef]
- Li, L.P.; Qu, L.; Li, T. Supplemental dietary Selenohomolanthionine affects growth and rumen bacterial population of Shaanbei white cashmere wether goats. Front. Microbiol. 2022, 13, 942848. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Lv, J.; Dou, X.; Zhang, Y. Effects of Dietary Supplementation with Clostridium butyricum on the Amelioration of Growth Performance, Rumen Fermentation, and Rumen Microbiota of Holstein Dairys. Front. Nutr. 2021, 8, 763700. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.W.; Bauman, D.E. Adaptations of glucose metabolism during pregnancy and lactation. J. Mammary Gland. Biol. Neoplasia 1997, 2, 265–278. [Google Scholar] [CrossRef]
- Kamke, J.; Kittelmann, S.; Soni, P.; Li, Y.; Tavendale, M.; Ganesh, S.; Janssen, P.H.; Shi, W.; Froula, J.; Rubin, E.M.; et al. Rumen metagenome and metatranscriptome analyses of low methane yield sheep reveals a Sharpea-enriched microbiome characterised by lactic acid formation and utilisation. Microbiome 2016, 4, 56. [Google Scholar] [CrossRef]
- Laarman, A.H.; Ruiz-Sanchez, A.L.; Sugino, T.; Guan, L.L.; Oba, M. Effects of feeding a calf starter on molecular adaptations in the ruminal epithelium and liver of Holstein dairy calves. J. Dairy Sci. 2012, 95, 2585–2594. [Google Scholar] [CrossRef] [PubMed]
- Lane, M.A.; Jesse, B.W. Effect of volatile fatty acid infusion on development of the rumen epithelium in neonatal sheep. J. Dairy Sci. 1997, 80, 740–746. [Google Scholar] [CrossRef]
- Sakata, T.; Tamate, H. Rumen epithelial cell proliferation accelerated by rapid increase in intraruminal butyrate. J. Dairy Sci. 1978, 61, 1109–1113. [Google Scholar] [CrossRef] [PubMed]
- Sakata, T.; Tamate, H. Rumen epithelium cell proliferation accelerated by propionate and acetate. J. Dairy Sci. 1979, 62, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Kuhla, S.; Zitnan, R.; Seyfert, H.M.; Schneider, F.; Hagemeister, H.; Chudy, A.; Löhrke, B.; Blum, J.W.; Hammon, H.M. Intraruminal infusion of n-butyric acid induces an increase of ruminal papillae size independent of IGF-1 system in castrated bulls. Arch. Anim. Nutr. 2005, 59, 213–225. [Google Scholar] [CrossRef]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly-Y, M.; Glickman, J.N.; Garrett, W.S. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Li, R.W.; Li, C. Butyrate induces profound changes in gene expression related to multiple signal pathways in bovine kidney epithelial cells. BMC Genom. 2006, 7, 234. [Google Scholar] [CrossRef]
- Li, C.J.; Li, R.W.; Wang, Y.H.; Elsasser, T.H. Pathway analysis identifies perturbation of genetic networks induced by butyrate in a bovine kidney epithelial cell line. Funct. Integr. Genom. 2007, 7, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Marinova, Z.; Leng, Y.; Leeds, P.; Chuang, D.M. Histone deacetylase inhibition alters histone methylation associated with heat shock protein 70 promoter modifications in astrocytes and neurons. Neuropharmacology 2011, 60, 1109–1115. [Google Scholar] [CrossRef]
- Yang, C.M. Response of forage fiber degradation by ruminal microorganisms to branched-chain volatile fatty acids, amino acids, and dipeptides. J. Dairy Sci. 2002, 85, 1183–1190. [Google Scholar] [CrossRef]
- Russell, J.B.; Hespell, R.B. Microbial rumen fermentation. J. Dairy Sci. 1981, 64, 1153–1169. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Yang, R.; Duan, C.; Wang, Y.; Hao, Q.; Ji, S.; Yan, H.; Zhang, Y.; Liu, Y. Effect of dioscorea opposite waste on growth performance, blood parameters, rumen fermentation and rumen bacterial community in weaned lambs. J. Integr. Agric. 2023, 22, 1833–1846. [Google Scholar] [CrossRef]
- Sun, C.; Chen, L.; Shen, Z. Mechanisms of gastrointestinal microflora on drug metabolism in clinical practice. Saudi Pharm. J. 2019, 27, 1146–1156. [Google Scholar] [CrossRef]
- Pokusaeva, K.; Fitzgerald, G.F.; van Sinderen, D. Carbohydrate metabolism in Bifidobacteria. Genes Nutr. 2011, 6, 285–306. [Google Scholar] [CrossRef] [PubMed]
- Bunesova, V.; Vlkova, E.; Rada, V.; Killer, J.; Musilova, S. Bifidobacteria from the gastrointestinal tract of animals: Differences and similarities. Benef. Microbes 2014, 5, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Lei, H.; Chen, G.; Chen, C.; Song, Y.; Cao, Z.; Zhang, C.; Zhang, C.; Zhou, J.; Lu, Y.; et al. In Vitro and In Vivo Studies Reveal that Hesperetin-7-O-glucoside, a Naturally Occurring Monoglucoside, Exhibits Strong Anti-inflammatory Capacity. J. Agric. Food Chem. 2021, 69, 12753–12762. [Google Scholar] [CrossRef]
- Soo, R.M.; Skennerton, C.T.; Sekiguchi, Y.; Imelfort, M.; Paech, S.J.; Dennis, P.G.; Steen, J.A.; Parks, D.H.; Tyson, G.W.; Hugenholtz, P. An expanded genomic representation of the phylum cyanobacteria. Genome Biol. Evol. 2014, 6, 1031–1045. [Google Scholar] [CrossRef]
- Feng, C.; Li, A.; Yin, C.; Wang, S.; Jin, W.; Liu, Y.; Huo, T.; Jiang, H. Realgar Alleviated Neuroinflammation Induced by High Protein and High Calorie Diet in Rats via the Microbiota-Gut-Brain Axis. Nutrients 2022, 14, 3958. [Google Scholar] [CrossRef]
- Wang, Y.; Nan, X.; Zhao, Y.; Jiang, L.; Wang, M.; Wang, H.; Zhang, F.; Xue, F.; Hua, D.; Liu, J. Rumen microbiome structure and metabolites activity in dairy cows with clinical and subclinical mastitis. J. Anim. Sci. Biotechnol. 2021, 12, 36. [Google Scholar] [CrossRef]
- Bach, A.; Lopez-Garcia, A.; Gonzalez-Recio, O.; Elcoso, G.; Fàbregas, F.; Chaucheyras-Durand, F.; Castex, M. Changes in the rumen and colon microbiota and effects of live yeast dietary supplementation during the transition from the dry period to lactation of dairy cows. J. Dairy Sci. 2019, 102, 6180–6198. [Google Scholar] [CrossRef]
- Perea, K.; Perz, K.; Olivo, S.K.; Williams, A.; Lachman, M.; Ishaq, S.L.; Thomson, J.; Yeoman, C.J. Feed efficiency phenotypes in lambs involve changes in ruminal, colonic, and small-intestine-located microbiota. J. Anim. Sci. 2017, 95, 2585–2592. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Wang, Z.; Li, D.; Zhu, B.; Xia, Y.; Wang, G.; Ai, L.; Zhang, C.; Wang, C. The gut microbiota as a target to improve health conditions in a confined environment. Front. Microbiol. 2022, 13, 1067756. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Jiao, N.; Zhu, R.; Zhang, Y.; Wu, D.; Wang, A.J.; Fang, S.; Tao, L.; Li, Y.; Cheng, S.; et al. Identification of microbial markers across populations in early detection of colorectal cancer. Nat. Commun. 2021, 12, 3063. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jin, T.; Zhang, N.; Li, J.; Wang, Y.; Kulyar, M.F.; Han, Z.; Li, Y. Effect of stocking density and age on physiological performance and dynamic gut bacterial and fungal communities in Langya hens. Microb. Cell Factories 2021, 20, 218. [Google Scholar] [CrossRef]
- Lun, H.; Yang, W.; Zhao, S.; Jiang, M.; Xu, M.; Liu, F.; Wang, Y. Altered gut microbiota and microbial biomarkers associated with chronic kidney disease. Microbiol. Open 2019, 8, e678. [Google Scholar] [CrossRef]
- Caspi, R.; Billington, R.; Keseler, I.M.; Kothari, A.; Krummenacker, M.; Midford, P.E.; Ong, W.K.; Paley, S.; Subhraveti, P.; Karp, P.D. The MetaCyc database of metabolic pathways and enzymes—A 2019 update. Nucleic Acids Res. 2020, 48, D445–D453. [Google Scholar] [CrossRef]
- Pietrocola, F.; Galluzzi, L.; Bravo-San, P.J.; Madeo, F.; Kroemer, G. Acetyl coenzyme A: A central metabolite and second messenger. Cell Metab. 2015, 21, 805–821. [Google Scholar] [CrossRef]
- Lacroux, J.; Atteia, A.; Brugiere, S.; Couté, Y.; Vallon, O.; Steyer, J.P.; van Lis, R. Proteomics unveil a central role for peroxisomes in butyrate assimilation of the heterotrophic Chlorophyte alga Polytomella sp. Front. Microbiol. 2022, 13, 1029828. [Google Scholar] [CrossRef]
- Carley, A.N.; Maurya, S.K.; Fasano, M.; Wang, Y.; Selzman, C.H.; Drakos, S.G.; Lewandowski, E.D. Short-Chain Fatty Acids Outpace Ketone Oxidation in the Failing Heart. Circulation 2021, 143, 1797–1808. [Google Scholar] [CrossRef]
- Sanchez, H.N.; Moroney, J.B.; Gan, H.; Shen, T.; Im, J.L.; Li, T.; Taylor, J.R.; Zan, H.; Casali, P. B cell-intrinsic epigenetic modulation of antibody responses by dietary fiber-derived short-chain fatty acids. Nat. Commun. 2020, 11, 60. [Google Scholar] [CrossRef]
- Mathewson, N.D.; Jenq, R.; Mathew, A.V.; Koenigsknecht, M.; Hanash, A.; Toubai, T.; Oravecz-Wilson, K.; Wu, S.R.; Sun, Y.; Rossi, C.; et al. Gut microbiome-derived metabolites modulate intestinal epithelial cell damage and mitigate graft-versus-host disease. Nat. Immunol. 2016, 17, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzi, S.C.; Sharon, I.; Wrighton, K.C.; Koren, O.; Hug, L.A.; Thomas, B.C.; Goodrich, J.K.; Bell, J.T.; Spector, T.D.; Banfield, J.F.; et al. The human gut and groundwater harbor non-photosynthetic bacteria belonging to a new candidate phylum sibling to Cyanobacteria. eLife 2013, 2, e1102. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Proportion (%) | |
|---|---|---|
| Starter | Growth Feed | |
| Diet composition (as-fed basis) | ||
| Corn | 32.00 | 39.73 |
| Soybean Meal | 17.00 | 9.73 |
| Cottonseed Meal | 2.73 | 3.27 |
| DDGS | 13.18 | 13.63 |
| Bran | 10.55 | 16.09 |
| Puffed Soybean Flour | 2.73 | |
| Wheat Flour | 4.54 | |
| NaHCO3 | 0.45 | 0.73 |
| 1 Premix | 7.73 | 7.73 |
| Oat Grass | 9.09 | 9.09 |
| Total | 100 | 100 |
| Nutrient Levels (as dry matter basis) | ||
| Dry Matter | 89.03 | 88.64 |
| Crude Protein | 19.13 | 17.63 |
| Ether Extract | 2.88 | 2.48 |
| Ash | 9.47 | 6.40 |
| Neutral Detergent Fiber | 27.92 | 34.57 |
| Acid Detergent Fiber | 13.36 | 13.27 |
| Ca | 1.10 | 0.97 |
| P | 0.73 | 0.71 |
| Items | Treatment 1 | SEM 2 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| CON | LBA | MBA | HBA | Treatment | Linear | Quadratic | ||
| IBW 3 (d 60, kg) | 91.0 | 93.1 | 91.4 | 91.9 | 0.6 | 0.68 | 0.83 | 0.58 |
| FBW 4 (d 102, kg) | 137.1 | 146.6 | 144.6 | 140.9 | 1.4 | 0.08 | 0.56 | 0.02 |
| ADG 5 (kg/d) | 1.07 b | 1.22 a | 1.27 a | 1.24 a | 0.03 | 0.03 | 0.02 | 0.06 |
| ADFI 6 (kg/d DM) | 3.05 b | 3.30 a | 3.34 a | 3.18 ab | 0.03 | 0.01 | 0.27 | 0.02 |
| F/G ratio 7 | 2.85 | 2.70 | 2.63 | 2.56 | 0.05 | 0.17 | 0.03 | 0.64 |
| Items | Treatment 1 | SEM 2 | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| CON | LBA | MBA | HBA | Treatment | Linear | Quadratic | ||
| VFAs 3 (mmol/L) | ||||||||
| Acetate | 49.20 | 49.54 | 52.12 | 52.90 | 1.77 | 0.86 | 0.41 | 0.95 |
| Propionate | 32.26 | 28.28 | 28.82 | 30.67 | 1.85 | 0.94 | 0.97 | 0.54 |
| Iso-butyrate | 0.46 b | 0.71 a | 0.83 a | 0.89 a | 0.05 | <0.01 | <0.01 | 0.26 |
| Butyrate | 6.90 b | 7.89 ab | 8.56 ab | 10.52 a | 0.47 | 0.04 | <0.01 | 0.58 |
| Iso-valerate | 0.87 | 1.09 | 1.22 | 1.19 | 0.12 | 0.76 | 0.34 | 0.62 |
| Valerate | 1.83 | 1.98 | 2.17 | 1.80 | 0.15 | 0.82 | 0.95 | 0.40 |
| Acetate/Propionate | 1.43 | 1.72 | 1.83 | 1.81 | 0.06 | 0.07 | 0.02 | 0.16 |
| TVFAs 4 (mmol/L) | 91.33 | 89.48 | 93.82 | 98.26 | 3.85 | 0.86 | 0.50 | 0.70 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, H.; Huang, Q.; Li, S.; Du, D.; Yu, W.; Guo, J.; Zhao, Z.; Yu, X.; Ma, F.; Sun, P. Effect of Dietary Benzoic Acid Supplementation on Growth Performance, Rumen Fermentation, and Rumen Microbiota in Weaned Holstein Dairy Calves. Animals 2024, 14, 2823. https://doi.org/10.3390/ani14192823
Dai H, Huang Q, Li S, Du D, Yu W, Guo J, Zhao Z, Yu X, Ma F, Sun P. Effect of Dietary Benzoic Acid Supplementation on Growth Performance, Rumen Fermentation, and Rumen Microbiota in Weaned Holstein Dairy Calves. Animals. 2024; 14(19):2823. https://doi.org/10.3390/ani14192823
Chicago/Turabian StyleDai, Haonan, Qi Huang, Shujing Li, Dewei Du, Wenli Yu, Jia Guo, Zengyuan Zhao, Xin Yu, Fengtao Ma, and Peng Sun. 2024. "Effect of Dietary Benzoic Acid Supplementation on Growth Performance, Rumen Fermentation, and Rumen Microbiota in Weaned Holstein Dairy Calves" Animals 14, no. 19: 2823. https://doi.org/10.3390/ani14192823