Serum 25(OH)D Analysis in Captive Pachyderms (Loxodonta africana, Elephas maximus, Diceros bicornis, Rhinoceros unicornis, Tapirus indicus) in Europe



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Vitamin D Analysis
2.3. Statistical Methods
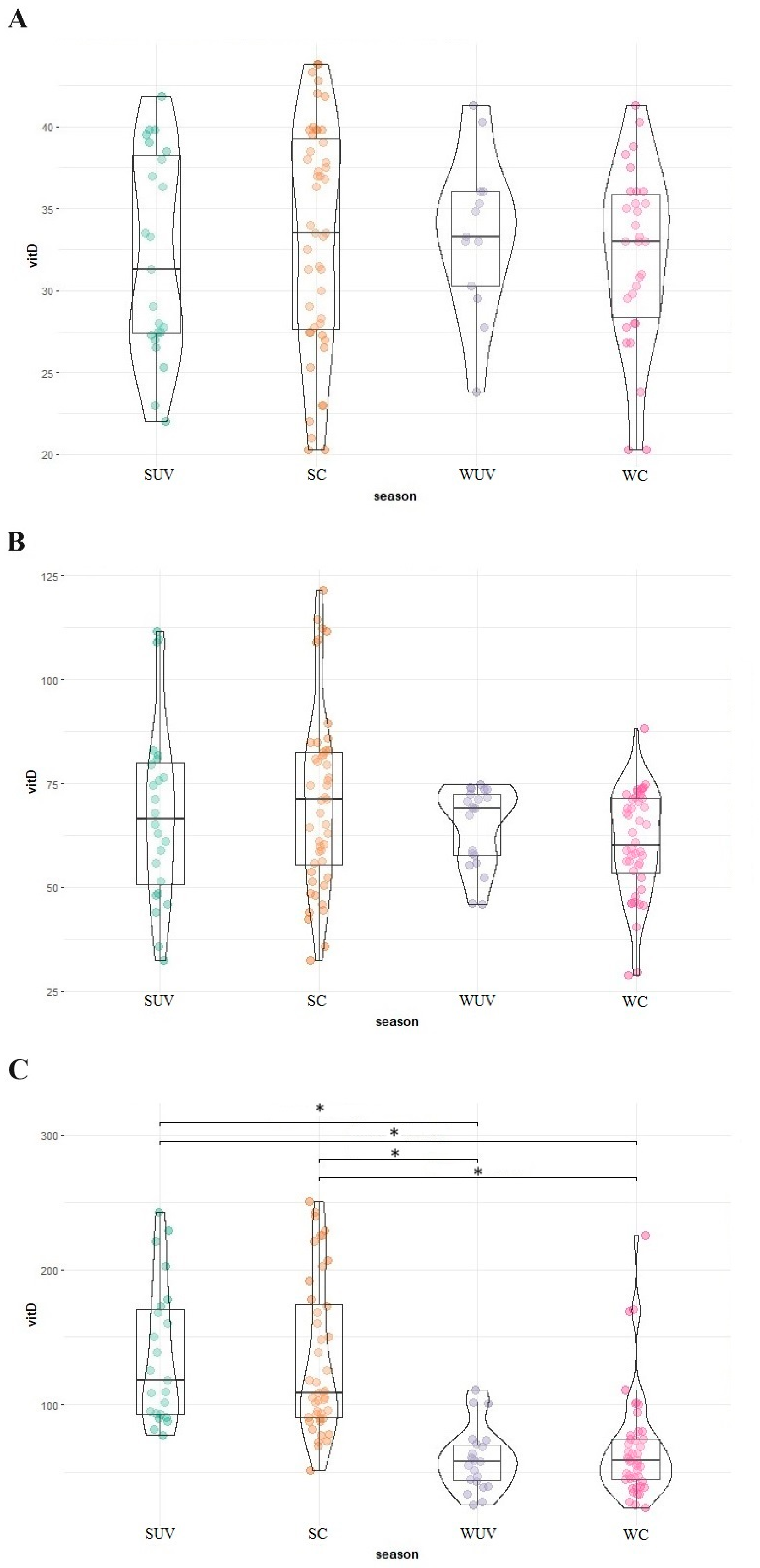
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roach, J.O.; Benyon, S. Nutrition. In Metabolism and Nutrition, 2nd ed.; Roach, J.O., Ed.; Elsevier Ltd.: London, UK; New York, NY, USA, 2003; pp. 154–156. [Google Scholar]
- Thitaram, C.; Pongsopawijit, P.; Thongtip, N.; Angkavanich, T.; Chansittivej, S.; Wongkalasin, W. Dystocia following prolonged retention of a dead fetus in an Asian elephant (Elephas maximus). Theriogenology 2006, 66, 1284–1291. [Google Scholar] [CrossRef] [PubMed]
- Mahasawangkul, S.; Angkawanich, T. Elephant health status in Thailand—The role of elephant hospitals and mobile elephant clinics. Eu-Asia link project symposium. In Managing the Health and Reproduction of Elephant Populations in Asia; Elephant Hospital: Lampang, Thailand; National Elephant Institute: Lampang, Thailand; Forest Industry Organization: Lampang, Thailand, 2007; pp. 32–37. [Google Scholar]
- Sanyathitisaeree, P.; Yartbantoong, N.; Thongthipsiridej, S.; Theeraphan, W. Elephant health problems: An accumulative case report from the Kasetsart university veterinary teaching hospital—Kamphaengsaen. Eu-Asia link project symposium. In Managing the Health and Reproduction of Elephant Populations in Asia; Faculty of Veterinary Medicine, Kasetsart University: Bangkok, Thailand, 2007; pp. 41–48. [Google Scholar]
- Hermes, R.; Saragusty, J.; Schaftenaar, W.; Göritz, F.; Schmitt, D.L.; Hildebrandt, T.B. Obstetrics in elephants. Theriogenology 2008, 70, 131–144. [Google Scholar] [CrossRef] [PubMed]
- van der Kolk, J.H.; van Leeuwen, J.P.T.M.; van den Belt, A.J.M.; van Schaik, R.H.N.; Schaftenaar, W. Subclinical hypocalcaemia in captive Asian elephants (Elephas maximus). Vet. Rec. 2008, 162, 475–479. [Google Scholar] [CrossRef] [PubMed]
- van Sonsbeek, G.R.; van der Kolk, J.H.; van Leeuwen, J.P.T.M.; Everts, H.; Marais, J.; Schaftenaar, W. Effect of calcium and cholecalciferol supplementation on several parameters of calcium status in plasma and urine of captive Asian (Elephas maximus) and African elephants (Loxodonta africana). J. Zoo Wildl. Med. 2013, 44, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Ensley, P.T.; Anderson, M.; Osborn, K.; Bissonnette, S.; Deftos, L. Osteodystrophy in an orphan Asian elephant. In Proceedings of the AAZV, Pittsburgh, PA, USA, 23–28 October 1994; pp. 142–143. [Google Scholar]
- Sach, F.; Dierenfeld, E.S.; Langley-Evans, S.C.; Watts, M.J.; Yon, L. African savanna elephants (Loxodonta africana) as an example of a herbivore making movement choices based on nutritional needs. PeerJ 2019, 7, e6260. [Google Scholar] [CrossRef]
- Childs-Sanford, S.E.; Makowski, A.J.; Wakshlag, J.J. The vitamin D status of Asian elephants (Elephas maximus) managed in a northern temperate climate. J. Zoo Wildl. Med. 2020, 51, 1–12. [Google Scholar] [CrossRef]
- Miller, M.; Chen, T.C.; Holick, M.F.; Mikota, S.; Dierenfeld, E. Serum concentrations of calcium, phosphorus, and 25-hydroxyvitamin D in captive African elephants (Loxodonta africana). J. Zoo Wildl. Med. 2009, 40, 302–305. [Google Scholar] [CrossRef]
- Dosi, M.C.M.C.; McGorum, B.C.; Kirton, R.D.; Cillán-García, E.; Mellanby, R.J.; Keen, J.A.; Hurst, E.A.; Morgan, R.A. The effect of season, management and endocrinopathies on vitamin D status in horses. Equine Vet. J. 2022, 55, 672–680. [Google Scholar] [CrossRef]
- Ullrey, D.E.; Crissey, S.D.; Hintz, H.F. Elephants: Nutrition and dietary husbandry. In Nutrition Advisory Group Handbook; American Zoo and Aquarium Association: Silver Spring, MD, USA, 1997; pp. 1–20. [Google Scholar]
- Pilgrim, M.; Biddle, R. EAZA Best Practice Guidelines Black Rhinoceros (Diceros bicornis), 2nd ed.; EAZA: Amsterdam, The Netherlands, 2020. [Google Scholar]
- AZA Tapir TAG. Tapir (Tapiridae) Care Manual; Association of Zoos and Aquariums: Silver Spring, MD, USA, 2013. [Google Scholar]
- BioMérieux. Vidas® 25 OH Vitamin D Total (vitD); Biomerieux: Maharashtra, India, 2016; pp. 1–9. [Google Scholar]
- KNMI/ESA UV Index. Available online: https://www.esa.int/Applications/Observing_the_Earth/Thanks_to_ESA_KNMI_offers_a_UV_forecasting_Service (accessed on 22 November 2023).
- IUCN—Number of Animals in Countries of Origin All Species Current Knowhow of the Different IUCN Specialist Groups. 2023. Available online: https://www.iucnredlist.org/ (accessed on 8 January 2024).
- Martini, P. General considerations and highlights of low-lying coastal zones: Passive continental margins from the poles to the tropics. In Sedimentary Coastal zones from High to Low Latitudes: Similarities and Differences; Martini, P., Walness, H.R., Eds.; Geological Society of London Special Publications: London, UK, 2014; Volume 388, pp. 1–32. [Google Scholar]
- Liczano, D.J.; Cavelier, J. Daily and seasonal activity of the mountain tapir (Tapirus pinchaque) in the Central Andes of Colombia. J. Zool. 2000, 252, 429–435. [Google Scholar]
- Zimmerman, D.M.; Hernandez, S. Tapiridae. In Zoo Wild Animal Medicine, 8th ed.; Miller, R.E., Fowler, M.E., Eds.; Elsevier Mosby Saunders: St. Louis, MO, USA, 2015; pp. 547–558. [Google Scholar]
- Van Sonsbeek, G.R.; van der Kolk, J.H.; van Leeuwen, J.P.T.M.; Schaftenaar, W. Preliminary validation of assays to measure parameters of calcium metabolism in captive Asian and African elephants in Western Europe. J. Vet. Diagn. Investig. 2011, 23, 504–510. [Google Scholar] [CrossRef]
- Olds, J.; Oltman, W.; Makowski, M.; Householder, H.; Keeley, L.L. Seasonal variation of serum 25-hydroxy-vitamin D in two captive Eastern black rhinoceros (Diceros bicornis michaeli) housed in a North American zoo. J. Zoo Wildl. Med. 2018, 49, 943–951. [Google Scholar] [PubMed]
- Hamilton, S. Rhinoceros Nutrition: A Comparative Approach. Bachelor’s Thesis, Department of Animal Science, University of Sydney, Sydney, Australia, 1999; p. 87. [Google Scholar]
- Clauss, M.; Jessup, D.A.; Norkus, E.B.; Chen, T.C.; Holick, M.F.; Streich, J.; Dierenfeld, E.S. Fat soluble vitamins in blood and tissues of free-ranging and captive rhinoceros. J. Wildl. Dis. 2002, 38, 402–413. [Google Scholar] [CrossRef] [PubMed]
- Perumbilly, A.J.; Balagangatharathilagar, M.; Kalaivanan, M.; Vishnurahav, R.B.; Begum, M.M. Quantitaive determination of 25-hydroxyvitamin D in adult captive Asian elephants (Elephas maximus) from Southern tropical regions of Tamil Nadu. Int. J. Adv. Biochem. Res. 2024, SP-8, 406–408. [Google Scholar] [CrossRef]
- Bapodra, P.; Dierenfeld, E.; Wolfe, B.A. Evaluation of season-related dietary changes on the serum profiles of fat soluble vitamins, mineral, fatty acids, and lipids in the captive greater-one horned rhinoceros (Rhinoceros unicornis). Zoo Biol. 2014, 33, 314–319. [Google Scholar] [CrossRef]
- Childs-Sanford, S.E.; Kiso, W.K.; Schmitt, D.L. Serum vitamin D and selected biomarkers of calcium homeostasis in Asian elephants (Elephas maximus) managed at a low latitude. J. Zoo Wildl. Med. 2024, 55, 430–435. [Google Scholar] [CrossRef]
- Hymøller, L.; Jensen, S.K. We know next to nothing about vitamin D in horses! J. Equine Vet. Sci. 2015, 35, 785–792. [Google Scholar] [CrossRef]
- Azarpeykan, S.; Dittmer, K.E.; Gee, E.K.; Marshall, J.C.; Walace, J.; Elder, P.; Acke, E.; Thompson, K.G. Influence of blanketing and season on vitamin D and parathyroid hormone, calcium, phosphorus, and magnesium concentrations in horses in New Zealand. Domest. Anim. Endocrinol. 2016, 56, 75–84. [Google Scholar] [CrossRef]
- Childs-Sanford, S.E.; Makowski, A.J.; Hilliard, R.L.; Wakshlag, J.J. Experimental cholecalciferol supplementation in a herd of managed Asian elephants (Elephas maximus). J. Zoo Wildl. Med. 2023, 54, 219–230. [Google Scholar] [CrossRef]
- Hurst, E.A.; Homer, N.Z.; Mellanby, J. Vitamin D metabolism and profiling in veterinary species. Metabolites 2020, 10, 371. [Google Scholar] [CrossRef] [PubMed]
- Azarpeykan, S.; Gee, E.K.; Thompson, K.G.; Dittmer, K.E. Undetectable vitamin D3 in equine skin irradiated with ultraviolet light. J. Equine Sci. 2022, 33, 45–49. [Google Scholar] [CrossRef]
- Bruins-van Sonsbeek, L.G.R. Personal observations. 2024. [Google Scholar]
- Cavaleros, M.; Buffenstein, R.; Ross, F.P.; Pettifor, J.M. Vitamin D metabolism in frugivorous nocturnal mammal, the Egyptian fruit bat (Rousettus aegyptiacus). Gen. Comp. Endocrinol. 2003, 133, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Southworth, L.O.; Holick, M.F.; Chen, T.C.; Kunz, T.H. Effects of sunlight on behaviour and 25-hydroxyvitamin D levels in two species of Old World fruit bats. Derm.-Endocrinol. 2013, 5, 192–198. [Google Scholar] [CrossRef] [PubMed]
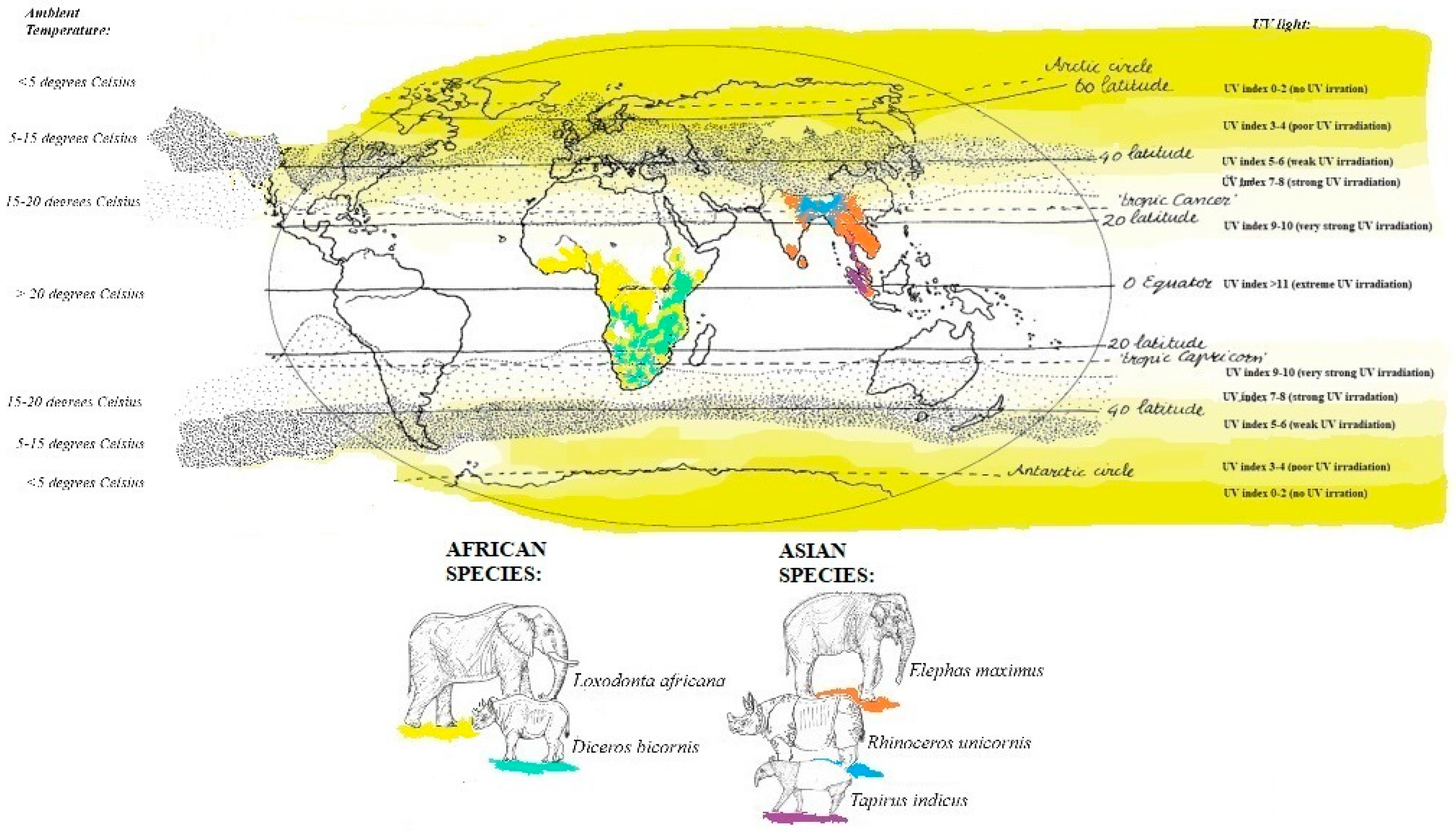


{kind=link}
{kind=link}
{kind=link}
| Zoo * | Individual | Species | Sex | Year of Birth | Age at Sample Collection (Years) | n | Cholecalciferol Intake (IU/kg BW) ** | Recommended Vitamin D Intake (IU/kg BW) |
|---|---|---|---|---|---|---|---|---|
| A | 1 | L. africana | f | 2015 | 5–6 | 9 | 6.5 | 8–121 |
| 2 | L. africana | f | 1985 | 35–36 | 18 | 6.5 | ||
| 3 | L. africana | f | 1986 | 34–35 | 18 | 6.5 | ||
| 4 | L. africana | f | 1992 | 28–29 | 19 | 6.5 | ||
| B | 5 | L. africana | f | 1993 | 27–29 | 6 | 3.6 | |
| 6 | L. africana | f | 2007 | 13–15 | 3 | 3.6 | ||
| 7 | L. africana | f | 2016 | 4–6 | 5 | 3.6 | ||
| 8 | L. africana | m | 2019 | 2–3 | 6 | 3.6 | ||
| 9 | L. africana | m | 1993 | 34–36 | 5 | 3.6 | ||
| 10 | L. africana | f | 1992 | 29–31 | 1 | 3.6 | ||
| C | 11 | E. maximus | f | 1970 | 50–53 | 25 | 5 | 12–151 |
| 12 | E. maximus | f | 2000 | 20–23 | 22 | 5 | ||
| 13 | E. maximus | f | 2003 | 17–19 | 20 | 5 | ||
| 14 | E. maximus | f | 2010 | 10–13 | 25 | 5 | ||
| 15 | E. maximus | m | 2021 | 1–2 | 2 | 5 | ||
| 16 | D. bicornis | m | 2001 | 19–20 | 19 | 2.5–6.6 *** | 3–92 | |
| 17 | D. bicornis | f | 2011 | 9–12 | 37 | 2.5–6.6 *** | ||
| 18 | D. bicornis | f | 2017 | 3–5 | 22 | 2.5–6.6 *** | ||
| 19 | D. bicornis | m | 2020 | 2–3 | 11 | 2.5–6.6 *** | ||
| 20 | D. bicornis | m | 2019 | 4 | 1 | 2.5–6.6 *** | ||
| 21 | R. unicornis | f | 2017 | 5–6 | 4 | 1.5 | - | |
| 22 | T. indicus | f | 2011 | 9–10, 12 | 32 | 10–30 **** | 10.53 | |
| 23 | T. indicus | m | 2016 | 7 | 8 | 5 |
| Species *** | Mean (±SD) | Median (Range) | Median Per Season (Range) | ||||
|---|---|---|---|---|---|---|---|
| Summer | Winter | ||||||
| Calendar (Apr–Sept) | UV Index > 5 (May–July) | Calendar (Oct–Mar) | UV Index < 2 (Nov–Jan) | ||||
| L. Africana * n = 90, 10 + | 34.5 (±9.0) | 33.2 (21.2–58.8) | 33.5 n = 47, 6 (20.3–43.8) | 31.3 n = 23, 6 (22.0–41.8) | 33.0 n = 30, 6 (20.3–41.3) | 33.3 n = 13, 6 (23.8–41.3) | |
| E. maximus n = 94, 5 | 75.3 (±21.1) | 68.7 (55.5–110.6) | 71.2 n = 48, 4 (32.5–122.0) | 66.5 n = 24, 4 (32.5–112.0) | 60.2 n = 44, 4 (29.0–88.3) | 69.0 n = 21, 4 (46.0–74.8) | |
| D. bicornis n = 90, 5 | 126.2 (±70.2) | 86.3 (72.9–237.0) | 109.0 † n = 44, 4 (51.5–251.0) | 118.0 † n = 23, 4 (77.5–243.0) | 58.3 n = 45, 4 (23.5–226.0) | 57.9 n = 22, 4 (26.0–111.0) | |
| R. unicornis n = 4, 1 | - | 107.9 (106.3–132.8) | 109.5 (105.3–132.8) | 121.2 (109.5–132.8) | 106.3 | 106.3 | |
| T. indicus ** n = 26, 2 | - | <20.3 (<20.3–33.5) | <20.3 (<20.3–< 20.3) | <20.3 (<20.3–< 20.3) | <20.3 (<20.3–33.5) | <20.3 (<20.3–33.5) | |
| Species | Continent * | 25(OH)D | 25(OH)D3 | 25(OH)D2 | |
|---|---|---|---|---|---|
| Horse | 0.1−69 undetectable n = 21; 40° S10 | <1.9−36.09 5−20 n = 21; 40° S10 | |||
| Captive | Captive | Wild | Captive | ||
| African elephant | Europe | 34.5 ± 9.0 n = 90, 10 +; 52° Ncurrent study | 11± 5 n = 5; 52° N12 15.6 ± 7.7 n = 6; 52° N13 | ||
| North America | 39.4 ± 18.7 n = 72, 14; 28° N5 | ||||
| Asian elephant | Europe | 75.3 ± 21.1 n = 90, 5; 52° Ncurrent study | 36 ± 11 n = 8; 52° N12 35.6 ± 11.7 n = 10; 52° N13 | ||
| North America | undetectable n = 72, 6; 43° N6,8 undetectable n = 22; 28° N8 undetectable n = 5; 43° N11 32.2 ± 8.7 ** n = 5; 43° N11 | 17.5 ± 2.2 n = 72, 6; 43° N6,8 5.8 ± 1.5 n = 33; 28° N8 12.3 ± 1.5 n = 5; 43° N11 12.1 ± 2.2 ** n = 5; 43° N11 | |||
| Asia | 27.7 ± 4.9 n = 10; 8−14° N4 | ||||
| Black rhinoceros | Europe | † 126.2 ± 70.2 n = 90, 5; 52° Ncurrent study | 0.24 ± 0 n = 22 | 139.0 ± 85 n = 28; 20° S3 | |
| North America | † 100.6 n = 40, 2; 41° N1 | ||||
| Indian rhinoceros | Europe | 107.9 (106.3−132.8) n = 4, 1; 52° Ncurrent study | |||
| North America | † 27.6 (winter 22.4 (± 2.93) - summer 32.8 (± 7.44)) n = 10, 5; 40° N7 | ||||
| Malayan tapir | < 20.3-33.5 n = 26, 2; 52° Ncurrent study | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bruins-van Sonsbeek, L.G.R.; Corbee, R.J. Serum 25(OH)D Analysis in Captive Pachyderms (Loxodonta africana, Elephas maximus, Diceros bicornis, Rhinoceros unicornis, Tapirus indicus) in Europe. Animals 2024, 14, 2843. https://doi.org/10.3390/ani14192843
Bruins-van Sonsbeek LGR, Corbee RJ. Serum 25(OH)D Analysis in Captive Pachyderms (Loxodonta africana, Elephas maximus, Diceros bicornis, Rhinoceros unicornis, Tapirus indicus) in Europe. Animals. 2024; 14(19):2843. https://doi.org/10.3390/ani14192843
Chicago/Turabian StyleBruins-van Sonsbeek, Linda G. R., and Ronald J. Corbee. 2024. "Serum 25(OH)D Analysis in Captive Pachyderms (Loxodonta africana, Elephas maximus, Diceros bicornis, Rhinoceros unicornis, Tapirus indicus) in Europe" Animals 14, no. 19: 2843. https://doi.org/10.3390/ani14192843