Simple Summary
The extracellular products (ECPs) secreted by virulent Vibrio mediterranei strains are pathogenic to bivalves. However, it remains unclear whether differences in pathogenicity between different V. mediterranei strains are associated with ECP compositions. In this study, genomics and proteomics approaches were integrated to compare the virulence factors in the ECP proteins secreted by two high-virulence V. mediterranei strains and one low-virulence strain. We found that the ECPs secreted by high-virulence V. mediterranei strains had a greater number and variety of virulence factors compared to low-virulence strains, and that 73 of the 95 specific ECP proteins in the high-virulence strains were not expressed in the low-virulence strain. These findings provide further insights into the role that virulent ECPs play in the pathogenesis of V. mediterranei to bivalves.
Abstract
Vibrio mediterranei, a bacterial pathogen of bivalves, has exhibited strain-dependent virulence. The mechanisms behind the variations in bivalve pathogenicity between V. mediterranei strains have remained unclear. However, a preliminary analysis of the extracellular product (ECP) proteomes has revealed differences in protein compositions between low- and high-virulence strains; in addition to 1265 shared proteins, 127 proteins have been identified to be specific to one low-virulence strain and 95 proteins to be specific to two high-virulence strains. We further studied the ECP proteins of the three V. mediterranei strains from functional perspectives using integrated genomics and proteomics approaches. The results showed that lipid metabolism, transporter activity and membrane transporter pathways were more enriched in the ECPs of the two high-virulence strains than in those of the low-virulence strain. Additionally, 73 of the 95 high-virulence strain-specific proteins were found to have coding genes in the genome but were not expressed in the low-virulence strain. Moreover, comparisons with known virulence factors in the Virulence Factor Database (VFDB) and the Pathogen–Host Interactions Database (PHI-base) allowed us to predict more than 10 virulence factors in the categories of antimicrobial activity/competitive advantage, the effector delivery system and immune modulation, and the high-virulence strain-specific ECP proteins consisted of a greater percentage of known virulence factors than the low-virulence strain. Particularly, two virulence factors, MtrC and KatG, were identified in the ECPs of the two high-virulence strains but not in those of the low-virulence strain. Most coding genes of the ECP proteins including known virulence factors were identified on chromosome 1 of V. mediterranei. Our findings indicate that variations in virulence factor composition in the bacterial ECPs may partially account for the differences in the bivalve pathogenicity between V. mediterranei strains.
1. Introduction
Vibrios are rod-shaped, Gram-negative bacteria naturally present in freshwater, estuarine and marine environments [1,2]. While most Vibrio species constitute a normal component of the microbial ecology in the related environments, some species are causative agents of vibriosis in plants, animals and humans [3,4]. At least 10 Vibrio species, for example, have been reported to be pathogenic or opportunistically pathogenic to humans [4,5].
Vibrios also encompass important pathogens of aquatic vertebrates and invertebrate animals, especially bivalves [6]. Vibrios infection causes larval vibriosis that represents a major threat to hatchery-reared larvae and spats of several bivalves [7,8,9,10,11]. In 2007, a Vibrio tubiashii-caused vibriosis resulted in an 80% mortality, leading to a 59% production reduction in oyster larvae in hatcheries on the West Coast of North America [8]. Vibrio infections also give rise to mortalities in adult bivalves. Vibrio alginolyticus, for example, was identified as the causative agent for a summer mortality incident in cultured Pacific oysters Crassostrea gigas in China [12]. Other Vibrio species that are reported to cause vibriosis in bivalves and other aquatic animals include Vibrio mediterranei [13], Vibrio carchariae [14], Vibrio splendidus [15], Vibrio aestuarianus [16], Vibrio neptunius [10], Vibrio anguillarum [17], Vibrio coralliilyticus [18] and Vibrio parahaemolyticus [19].
However, pathogenicity is not a species-specific quality for these pathogenic Vibrios. Significant differences in the virulence and the degree of pathogenicity are observed between strains of the above-mentioned pathogenic or opportunistically pathogenic Vibrio species [20,21]. This difference in virulence between strains within a group or species is determined by the qualitative or quantitative variations in virulence factors [22]. Virulence factors are defined as pathogen components that damage the host [23,24]. Bacterial virulence factors consist of extremely diverse components that are involved in the entire host infection process including adherence, invasion and survival in the host and environments. The latest version of the Virulence Factor Database (VFDB), for example, classifies 11,027 bacterial virulence factors into 14 categories (accessed on November 2023) [25]. A wide variety of virulence factors associated with the adherence, motility, invasion, iron uptake subcategory of the nutritional/metabolic factor, exotoxin, effector delivery system, exoenzyme, regulation and biofilm have been identified in Vibrio species pathogenic to aquatic animals [12,26,27,28,29,30].
Pathogenic bacteria secrete virulence factors as compositions of extracellular products (ECPs), which are believed to play a role in the bacteria’s pathogenicity. Aquatic pathogen ECPs have been reported to contain proteases, lipases, DNases, chitinases, hemolysins and polysaccharides [31,32,33]. These factors contribute to extensive tissue damage, enabling bacteria to acquire nutrients and proliferate within hosts [33,34]. Certain components, such as ECPs, serine proteases, and metalloproteases, have been documented as lethal to shrimp and oyster larvae [35,36]. Haemolysins are also implicated in the pathogenicity of bacteria toward shrimp and fish [37,38]. Additionally, ECP chitinases play a role in facilitating bacterial penetration of crustacean shells [39]. However, it remains to be further studied whether the qualitative and/or quantitative differences in ECP virulence factors represent a determinant factor for the intra-specific variations in the pathogenicity of a bacterial pathogen.
We have characterized V. mediterranei as the causative agent of a recent massive mortality incident in a razor clam hatchery [13]. Differences in pathogenicity to larval and juvenile razor clams and oysters have been observed between bacterial strains in experimental infections with cultured bacteria [13]. ECPs of one low-virulence strain (DT02) and two high-virulence strains (DT07 and RW01) also demonstrate correspondent virulence differences [13]. In addition, DT02 ECPs contain higher metalloproteinase and chitinase activities than the DT07 and RW01 ECPs [13]. Intriguingly, proteomic analyses of the ECPs of the three V. mediterranei strains using liquid chromatography–tandem mass spectrometry (LC-MS/MS) revealed 1265 proteins shared by all three strains, 95 ECP proteins specific to the two high-virulence strains (i.e., DT07 and RW01), and 127 ECP proteins specific to the one low-virulence strain (i.e., DT02) [13]. However, the functional significance of the differences in the proteome composition between V. mediterranei strains with different virulence remained to be investigated.
In the present research, we performed a comprehensive function analysis of the ECP proteins identified in the previous research [13] with combined genomics and proteomics approaches. Our goals were to: (1) functionally annotate the 1265 proteins shared by all three strains, the 95 proteins specific to the two high-virulence strains and the 127 proteins specific to the one low-virulence strain using KEGG pathway and GO term analyses; (2) identify known virulence factors in the ECP proteins through searching published virulence factor databases; (3) determine the coding genes of the ECP proteins in the bacterial genomes; and (4) assess the expression of the related coding genes in different V. mediterranei strains. The results may provide important information about the mechanisms underlying the intra-specific virulence differences in V. mediterranei and other bacterial pathogens.
2. Materials and Methods
2.1. V. mediterranei Strains, Proteomic Sequence Datasets and Genomic Sequences
V. mediterranei strain isolation and characterization, ECP preparation and proteomics analysis were reported in detail previously [13]. Bacterial strain DT02, a low-virulence strain isolated from a diseased razor clam juvenile, and strains DT07 and RW01, 2 high-virulence strains isolated from a diseased razor clam juvenile and rearing water, respectively, were used in this research. The proteomics datasets selected for analysis included the 1265 ECP proteins shared by all three bacterial strains, the 95 ECP proteins specific to the two high-virulence strains and the 127 ECP proteins specific to the one low-virulence strain.
Genome sequencing of the three V. mediterranei strains was carried out using high-throughput sequencing in the Illumina NovaSeq 6000 system and nanopore sequencing in an Oxford Nanopore PromethION 48 platform. The genomes consisted of 2 circular chromosomes of 3,762,826 bp and 2,025,510 bp, respectively, and 10 plasmids for strain DT02, 2 circular chromosomes of 3,666,428 bp and 2,211,640 bp, respectively, 1 integron and 5 plasmids for strain DT07, and 2 circular chromosomes of 3,771,825 bp and 2,182,795 bp, respectively, and 6 plasmids for strain RW01. The bacterial genomes were de novo assembled using SMRT Link v5.1.0 (https://www.pacb.com/support/software-downloads/), (accessed on 9 November 2021) and Unicycle (https://github.com/rrwick/Unicycler), (accessed on 9 November 2021), and further optimized to correct the misassembled regions using arrow software v2.2.1 [40,41]. Protein-coding genes were predicted using the program Glimmer 3 [42]. Predicted coding gene sequences were compared with bacterial proteins in the UniProt database using the program BLASTX [43]. The average nucleotide identity (ANI) of the genomes between strains was determined using the program ANItools (http://ani.mypathogen.cn/), (accessed on 13 June 2023) [44].
2.2. Identification of ECP Protein-Coding Genes in the Genomes
Coding genes of the ECP proteins determined by proteomics were identified by aligning the amino acid sequence to the genome sequences of the three V. mediterranei strains using the program BLAST GUI Wrapper in TBtools [45]. The number of genes distributed on each chromosome was plotted as a histogram using Chiplot Tools (https://www.chiplot.online/), (accessed on 24 July 2023).
2.3. Functional Annotations of the ECP Proteins
General biological functions of the V. mediterranei ECP proteins were predicted using Gene Ontology (GO) enrichment analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis [46,47]. GO analyses were carried out using the Gene Ontology Annotation (GOA) database at the UniProt Knowledgebase (UniProtKB) (http://www.ebi.ac.uk/GOA/), (accessed on 22 July 2023). KEGG pathway analyses were done first by getting each protein’s KEGG annotation using the KEGG Automatic Annotation Server (KAAS) (http://www.genome.jp/kaas-bin/kaas_main), (accessed on 21 July 2023) and then by mapping the annotated proteins into KEGG pathways using the KEGG Mapper online (http://www.kegg.jp/kegg/mapper.html), (accessed on 21 July 2023). GO enrichment and KEGG pathway analysis results were compared between protein sets and plotted in histograms using the OmicShare Tools (https://www.omicshare.com/tools), (accessed on 23 July 2023).
2.4. Prediction of Virulence Factors
Bacterial virulence factors were predicted by online searching the Virulence Factor Database (VFDB) (www.mgc.ac.cn/VFs/main.htm), (accessed on 9 October 2023) [48] and the Pathogen–Host Interactions Database (PHI-base) (www.phi-base.org/), (accessed on 12 October 2023) [49]. VFDB categorizes all reported bacterial virulence factors according to their functions in the pathogenesis process, and PHI-base uses pathogen–host interactions as virulence attributes and classifies them into 9 high-level phenotypes according to the interaction outcomes [25,50].
2.5. Quantitation of ECP Protein Expression
Protein expression was assessed based on the peak intensity of each protein in the LC-MS/MS detection. Peak intensities of each protein were normalized in a globally fashion [51,52]. The normalized values were then treated as protein expression levels, and the results were presented in Heatmap.
3. Results
3.1. Coding Genes of the ECP Proteins
For the 1265 ECP proteins shared by all three strains, a total of 1165 coding genes were identified in the three genomes, but each genome was determined to host different numbers of coding genes, with 1154 in DT07, 1153 in RW01 and 1151 in DT02. Differences in specific genes contained in each genome were also observed. The DT07 genome contained 9 and 12 genes that were not identified in RW01 and DT02, respectively. These DT07-specific coding genes included known virulence factors (plr/gapA, purCD, plr/gapA, kasB and nmpC). In the meantime, the DT07 genome lacked eight genes contained in the RW01 genome and eight genes in the DT02 genome (Table 1). One hundred of the shared ECP proteins were not identified to have coding genes in any of the three genomes (Table S1).

Table 1.
Information for 30 ECP proteins with different coding genes for the 1265 ECP proteins shared by all 3 strains.
For the 95 ECP proteins specific to the two high-virulence strains, 72 coding genes were determined in the DT07 genome and 73 in the RW01 genome. The low-virulence DT02 genome was also found to contain 66 coding genes for the 95 ECP proteins specific to the two high-virulence strains. The DT02 genome lacked seven genes, including the coding genes for the virulence factor katG, compared to the two high-virulence strains. In addition, the RW01 genome contained a coding gene for the virulence factor tcdA, which was not found in the DT07 genome (Table 2). A total of 22 proteins did not have coding genes in any of the three genomes (Table S2).
Table 2.
Differential presence of 7 ECP protein-encoding genes in the DT02 genome compared to DT07 and RW01 genomes.
For the 127 ECP proteins specific to the low-virulence strain DT02, 91 coding genes were determined and 28 genes were missing in the strain’s genome (Table S3). The genomes of the two high-virulence strains contained 92 coding genes. The DT02 genome contained seven genes that were not identified in the DT07 genome and six genes that were missing in the RW01 genome, but it lacked eight genes that were present in the DT07 genome and seven genes that were absent in the RW01 genome (Table 3).
Table 3.
Information for the 15 out of 127 ECP proteins specific to the 1 low-virulence strain with different coding genes in the all 3 strains genomes.
3.2. Functional Annotation and Pathway Assignment of ECP Proteins
GO enrichment analysis annotates proteins in GO terms and categorizes them in three aspects, biological process, cellular component and molecular function. For all three sets of V. mediterranei ECP proteins, the first two most-enriched GO terms were the cellular process and the metabolic process in the biological process category, the cellular anatomical entity and the protein-containing complex in the cellular component category, and catalytic activity and binding in the molecular function category (Figure 1). Compared to the 1265 ECP proteins shared by all three strains and 127 proteins specific to the one low-virulence strain, however, the 95 proteins specific to the two high-virulence strains showed a relatively higher percentage enrichment in the GO terms of response to stimulus, biological regulation, homeostatic process, interspecies interaction between organisms, detoxification, viral process, protein-containing complex, transporter activity, antioxidant activity and molecular transducer activity.
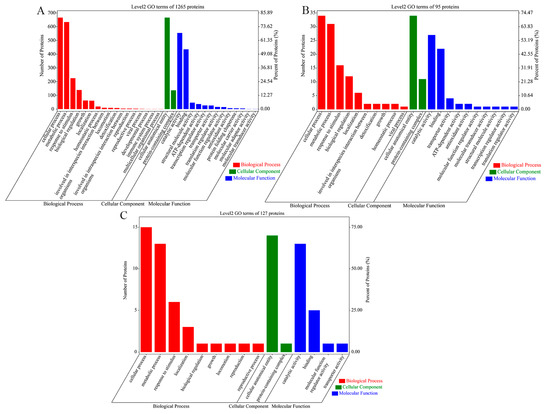
Figure 1.
GO analysis of the 3 sets of V. mediterranei ECP proteins. The numbers of ECP proteins and their percentages among the proteins analyzed are shown. (A) The 1265 ECP proteins shared by all 3 strains; (B) 95 proteins specific to the 2 high-virulence strains; (C) 127 proteins specific to the 1 low-virulence strain.
KEGG pathway analysis functionally annotates proteins into different pathways that are further classified in categories of metabolism, genetic information processing, environment information processing, cellular process and organismal systems. For all three V. mediterranei ECP protein sets, the most annotated pathways were in the metabolism category, with global and overview maps and carbohydrate metabolism being the most abundant (Figure 2). The 1265 ECP proteins shared by all three strains involved five categories with 23 pathways (Figure 2A). The 95 proteins specific to the two high-virulence strains and the 127 proteins specific to the one low-virulence strain were identified to have significantly fewer pathways than the 1265 shared proteins, and particularly, neither involved the organismal systems category (Figure 2B,C). Compared to the three strains’ shared proteins and the low-virulence strain-specific proteins, the 95 proteins specific to the two high-virulence strains showed higher relative abundance for the pathways of metabolism of cofactors and vitamins, metabolism of other amino acids, lipid metabolism, folding, sorting and degradation, replication and repair, membrane transport, and cellular community-prokaryotes (Table 4).
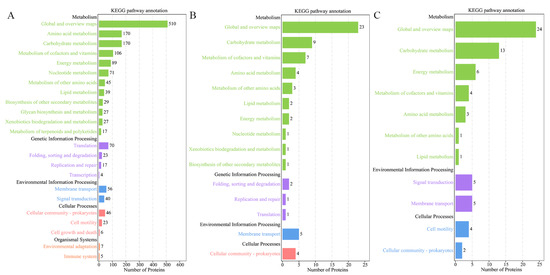
Figure 2.
KEGG analysis of the 3 sets of V. mediterranei ECP proteins. (A) The 1265 ECP proteins shared by all 3 strains; (B) 95 proteins specific to the 2 high-virulence strains; (C) 127 proteins specific to the 1 low-virulence strain.
Table 4.
Comparison of the relative abundance (percentage) of each KEGG pathway in the 3 sets of V. mediterranei ECP proteins.
3.3. Virulence Factors in ECP Proteins
Virulence factors in each of the three sets of V. mediterranei ECP proteins were predicted by comparison of known virulence factors via searches of the databases VFDB and PHI-base (Figure 3). VFDB searches identified 246 virulence factors belonging to 13 classes in the 1265 ECP proteins shared by all three strains, and most of them involved the nutritional/metabolic factor, immune modulation, adherence and motility (Figure 3A and Table S4). The searches also identified 13 virulence factors in the 95 ECP proteins specific to the two high-virulence strains, which belonged to six classes, and most of them involved the nutritional/metabolic factor, immune modulation, the effector delivery system and adherence (Figure 3A and Table S5). In the 127 proteins specific to the one low-virulence strain, VFDB searches identified 15 virulence factors that belonged to seven classes, and most of them involved the nutritional/metabolic factor, motility and the effector delivery system (Figure 3A and Table S6). The virulence factors that were related to antimicrobial activity/competitive advantage, the effector delivery system and immune modulation exhibited a higher percentage in the 95 proteins specific to the two high-virulence strains than in the 1265 ECP proteins shared by all three strains and 127 proteins specific to the one low-virulence strain (Figure 3A). In addition, the high-virulence ECP proteins contained a membrane fusion protein, MtrC, that was not identified in the two ECP protein sets (Table S5).
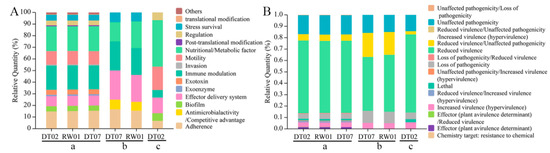
Figure 3.
Percentage of virulence factor classes in the 3 sets of V. mediterranei ECP proteins identified by searches of the databases VFDB (A) and PHI-base (B). a: the 1265 ECP proteins shared by all 3 strains; b: 95 proteins specific to the 2 high-virulence strains; c: 127 proteins specific to the 1 low-virulence strain.
The PHI-base search predicted 583 virulence factors that were categorized into seven classes in the 1265 ECP proteins shared by all three strains, and most of them were related to the classes of reduced virulence, unaffected pathogenicity/loss of pathogenicity, increased virulence (hypervirulence) and reduced virulence/unaffected pathogenicity (Figure 3B and Table S7). The PHI-base search identified 20 virulence factors that belonged to four classes, and most of them were associated with the classes of reduced virulence and reduced virulence/unaffected pathogenicity (Figure 3B and Table S8). In the 127 proteins specific to the one low-virulence strain, the PHI-base search found 35 virulence factors that involved five classes, and most of them belonged to the classes of reduced virulence, unaffected pathogenicity and loss of pathogenicity (Figure 3B and Table S9).
3.4. Distributions of ECP Protein-Coding Genes in Genomes
Analysis of the genomes of the three V. mediterranei strains disclosed that the coding genes for the ECP proteins identified in this study, including the predicted virulence factors, were present on the two chromosomes. No ECP protein-coding genes were found in non-chromosome genome elements. In addition, chromosome 1 contained most of the ECP protein-coding genes (Figure 4).
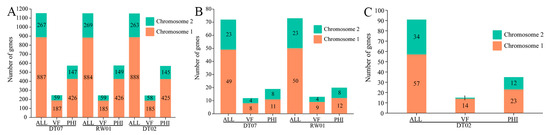
Figure 4.
Distributions of the coding genes for the ECP proteins shared by all 3 V. mediterranei (A), specific to the high-virulence strains (B) and the low-virulence strain (C). ALL: all ECP protein-coding genes in the strain; VF: virulence factors in ECP proteins searched in VFDB; PHI: virulence factors in ECP proteins searched in PHI-base.
3.5. Expression of Genes Coding for ECP Proteins
The coding genes of 1165 out of 1265 ECP proteins, which are shared by all three strains, were found to be expressed in all strains. However, among the 95 ECP proteins specific to the two high-virulence strains, 73 had coding genes present in three genomes and were expressed only in the two high-virulence strains. Additionally, variations in the expression levels of coding genes were observed between the two high-virulence strains. Specifically, 17 coding genes exhibited higher expression levels in DT07 compared to RW01, while 13 coding genes showed higher expression levels in RW01 compared to DT07 (Figure 5).
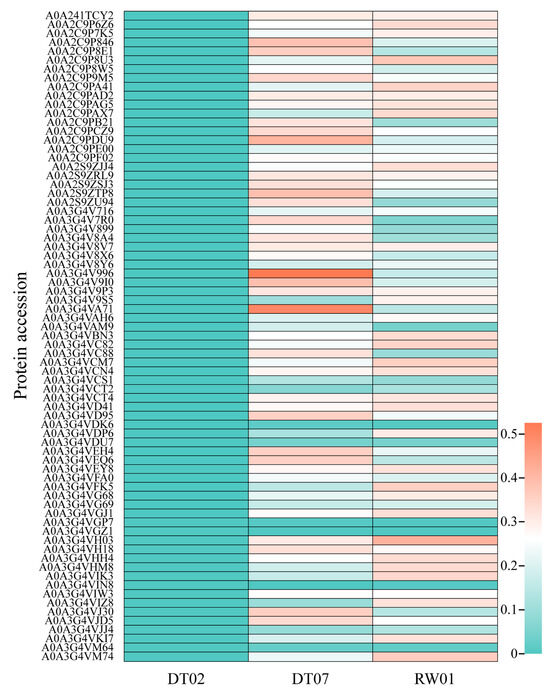
Figure 5.
Expression of 73 out of 95 ECP proteins specific to the 2 high-virulence strains with coding genes in the 3 V. mediterranei strains.
4. Discussion
ECPs secreted by Vibrios are associated with the bacteria’s pathogenicity to bivalves [13,16,53]. Some components such as proteases are believed to play a role in this attribute of bivalve-pathogenic Vibrios [16,54]. We have characterized V. mediterranei as the causative agent of larval vibriosis in hatchery-reared clams and oysters, and uncovered variations in pathogenicity between the bacterial isolates. Additionally, the differences in enzyme activities and in the proteomic compositions of the ECPs from bacterial strains differing in virulence also suggested the possible association with the bacterium’s pathogenicity [13]. Results of a combined genomic and proteomic analysis and particularly the discovery of quantitative and qualitative variations in virulence factors in ECPs provide further insights into the mechanism underlying the intra-specific variations in bivalve pathogenicity of V. mediterranei in this research.
Our analyses revealed the differences in the general proteomic composition and virulence factors between ECPs from different V. mediterranei strains, indicating the contribution of the bacterial ECPs to the bacterial virulence. At the proteomic level, both low- and high-virulence strains were observed to have proteins in their ECPs [13]. Annotation of the shared and strain-specific proteins further suggests their functional differences. We found in the GO enrichment and KEGG pathway analyses that the pathways involved in lipid metabolism and transporter activity and the membrane transporter pathways are more enriched in the 95 high-virulence strain-specific proteins as compared to those in the 1265 shared and 127 low-virulence-specific proteins. The lipid metabolism pathway is thought to be important for pathogenic bacteria to establish stable infections in hosts [55]. In addition, the membrane transporter KEGG pathway and the transporter activity GO term are associated with increased virulence in pathogenic bacteria [56].
More importantly, differences in the ECPs containing known virulence factors were found between the two types of bacterial strains. For example, 13.7% (13/95) of the high-virulence strain-specific ECP proteins are predicted to be virulence factors. This ratio was 11.8% (15/127) for the low-virulence strain. It appears that the high-virulence ECPs contain more virulence factors that are involved in pathogen adherence, the effector delivery system and host immune modulation. Several predicted virulence factors that are present in the ECPs of high-virulence V. mediterranei strains but not in the low-virulence strain are particularly worth noting. These include the stress survival-related virulence factor KatG, the antimicrobial activity/competitive advantage-related virulence factor MtrC, and the unaffected pathogenicity-related virulence factor ToxR. KatG is a catalase that degrades hydrogen peroxide and organic peroxides, and has been reported to protect pathogenic bacteria from the toxic effects of organic peroxides and enhance drug resistance, and thereby facilitate their survival and growth in hosts [57,58,59,60]. MtrC is a component of the MtrCDE efflux system [61]. The system is found to protect pathogenic bacteria from antimicrobial substances on mucosal surfaces and promotes their survival during infection [62]. ToxR is a transmembrane transcriptional regulator [63,64]. V. harveyi strains with higher toxR expression levels exhibit higher virulence to shrimp [65]. Thus, these high-virulence-specific virulence factors likely play a part in the increased pathogenicity in the related V. mediterranei strains. This phenomenon has been observed in other pathogenic Vibrios; the virulence factor pJM1 is important to the pathogenicity of the high-virulence V. anguillarum strain [66]. It should also be noted that there were differences in virulence factor composition between the two high-virulence V. mediterranei strains in this research. The virulence factors plr/gapA and purCD contained in strain DT07 were not found in strain RW01. A comparable observation has been made in the human pathogen Vibrio cholerae, wherein virulence factors such as ctxA, ctxB, and tcpA, present in the pathogenic non-O1 strain, are notably absent in the pathogenic O1 strain [67]. These findings suggest that virulence factors may function differently in different pathogenic bacteria.
Genomic analysis determined that the coding genes of all the ECP proteins from the three V. mediterranei strains, including those for the virulence factors, are present in the two chromosomal genomes. It is speculated that genes in the chromosomes are usually related to the phenotypes at the species level of a bacterium [68]. In contrast, some strain-specific phenotypes such as antibiotic resistance and virulence factors are often controlled by genes carried by plasmids [69]. This appears to be inconsistent with our results regarding the distribution of ECP protein-coding genes in the bacterial genomes. One explanation for the phenomenon may be related to the regulation at the gene expression level. Indeed, we found that most (66/95) of the coding genes for the high-virulence strain-specific ECP proteins were also present in the genome of the one low-virulence strain, which is, in fact, close to the number of coding genes identified in the genomes of the two high-virulence strains (72 and 73). It is apparent that the expression of those related genes in the low-virulence genome is inhibited by certain regulators. The mechanisms underlying the regulation of gene expression should be approached in future studies. On the other hand, differences in the number of coding genes and the specific genes involved in the different strains may involve the species evolution of V. mediterranei, and the associated mechanisms also need to be further studied. Interestingly, the coding genes for some ECP proteins identified in the proteomic analysis are found in the corresponding genomes. Studies will be carried out to investigate whether this is related to technical errors, unidentified genes in the genomes or environmental contaminations.
5. Conclusions
To summarize, a comprehensive analysis of the ECP protein compositions of three V. mediterranei strains that vary in pathogenicity to bivalves using a combination of genomics and proteomics resulted in the discovery of functional variations between low- and high-virulence strains. Particularly, differences in virulence factor composition in ECPs are observed. These findings provide solid evidence for the speculation that bacterial ECPs play an important role in the previously observed virulence differences between V. mediterranei strains. In addition, the results of our present research suggest that the inter-strain variations of ECP composition, including the virulence factors, are likely controlled by the regulation of expression of related coding genes. Future studies on the evolution and regulation of expression of the ECP protein-coding genes will generate more information about the pathogenesis of V. mediterranei and other Vibrio species in aquatic organisms.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ani14050692/s1, Table S1: Information about the 100 ECP proteins without coding genes for the 1265 ECP proteins shared by all 3 strains. Table S2: Information about the 22 ECP proteins without coding genes in the 95 ECP proteins specific to the 2 high-virulence strains. Table S3: Information about the 28 ECP proteins without coding genes in the 127 ECP proteins specific to the 1 low-virulence strain. Table S4: Virulence factors in the 1265 ECP proteins shared by all 3 bacterial strains and their classification in VFDB. Table S5: Virulence factors in the 95 ECP proteins specific to the 2 high-virulence strains and their classification in VFDB. Table S6: Virulence factors in the 127 ECP proteins specific to the low-virulence strain and their classification in VFDB. Table S7: Virulence factors in the 1265 ECP proteins shared by all 3 bacterial strains and their classification in PHI-base. Table S8: Virulence factors in the 95 ECP proteins specific to the 2 high-virulence strains and their classification in PHI-base. Table S9: Virulence factors in the 127 ECP proteins specific to the low-virulence strain and their classification in PHI-base.
Author Contributions
Data curation, C.F.; investigation, Q.X., Z.L., W.D., H.Z. and S.L.; project administration, Q.X.; writing—original draft, C.F. and W.D; writing—review and editing, C.F., Q.X. and W.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (32273110, 32073010, 32202989) and the 3315 Innovative Team Project of Ningbo City.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Baker-Austin, C.; Trinanes, J.; Gonzalez-Escalona, N.; Martinez-Urtaza, J. Non-Cholera Vibrios: The Microbial Barometer of Climate Change. Trends Microbiol. 2017, 25, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Thompson, F.L.; Iida, T.; Swings, J. Biodiversity of vibrios. Microbiol. Mol. Biol. Rev. 2004, 68, 403–431. [Google Scholar] [CrossRef] [PubMed]
- Grimes, D.J. The Vibrios: Scavengers, Symbionts, and Pathogens from the Sea. Microb. Ecol. 2020, 80, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Oliver, J.D.; Alam, M.; Ali, A.; Waldor, M.K.; Qadri, F.; Martinez-Urtaza, J. Vibrio spp. infections. Nat. Rev. Dis. Primers 2018, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Nair, G.B.; Shinoda, S. Pathogenic Vibrios in the Natural Aquatic Environment. Rev. Environ. Health 1997, 12, 63–80. [Google Scholar] [CrossRef] [PubMed]
- Paillard, C.; Le Roux, F.; Borrego, J.J. Bacterial disease in marine bivalves, a review of recent studies: Trends and evolution. Aquat. Living Resour. 2004, 17, 477–498. [Google Scholar] [CrossRef]
- Elston, R.; Leibovitz, L.; Relyea, D.; Zatila, J. Diagnosis of vibriosis in a commercial oyster hatchery epizootic: Diagnostic tools and management features. Aquaculture 1981, 24, 53–62. [Google Scholar] [CrossRef]
- Elston, R.A.; Hasegawa, H.; Humphrey, K.L.; Polyak, I.K.; Häse, C.C. Re-emergence of Vibrio tubiashii in bivalve shellfish aquaculture: Severity, environmental drivers, geographic extent and management. Dis. Aquat. Organ. 2008, 82, 119–134. [Google Scholar] [CrossRef]
- Prado, S.; Dubert, J.; Barja, J.L. Characterization of pathogenic vibrios isolated from bivalve hatcheries in Galicia, NW Atlantic coast of Spain. Description of Vibrio tubiashii subsp. europaensis subsp. nov. Syst. Appl. Microbiol. 2015, 38, 26–29. [Google Scholar] [CrossRef]
- Prado, S.; Romalde, J.L.; Montes, J.; Barja, J.L. Pathogenic bacteria isolated from disease outbreaks in shellfish hatcheries. First description of Vibrio neptunius as an oyster pathogen. Dis. Aquat. Organ. 2005, 67, 209–215. [Google Scholar] [CrossRef]
- Sakatoku, A.; Hatano, K.; Tanaka, S.; Isshiki, T. Isolation and Characterization of a Vibrio Sp. Strain MA3 Possibly Associated with Mass Mortalities of the Pearl Oyster Pinctada fucata. Arch. Microbiol. 2021, 203, 5267–5273. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Zhai, S.; Li, X.; Tian, J.; Liu, S. Identification of Vibrio alginolyticus as a causative pathogen associated with mass summer mortality of the Pacific Oyster (Crassostrea gigas) in China. Aquaculture 2021, 535, 736363. [Google Scholar] [CrossRef]
- Fan, C.; Liu, S.; Dai, W.; He, L.; Xu, H.; Zhang, H.; Xue, Q. Characterization of Vibrio mediterranei Isolates as Causative Agents of Vibriosis in Marine Bivalves. Microbiol. Spectr. 2023, 11, e0492322. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.K.; Liu, P.C.; Chen, Y.C.; Huang, C.Y. The implication of ambient temperature with the outbreak of vibriosis incultured small abalone Haliotis diversicolor supertexta Lischke. J. Therm. Biol. 2001, 26, 585–587. [Google Scholar] [CrossRef]
- Sugumar, G.; Nakai, T.; Hirata, Y.; Matsubara, D.; Muroga, K. Vibrio splendidus biovar II as the causative agent of bacillary necrosis of Japanese oyster Crassostrea gigas larvae. Dis. Aquat. Organ. 1998, 33, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Labreuche, Y.; Soudant, P.; Gonçalves, M.; Lambert, C.; Nicolas, J.-L. Effects of extracellular products from the pathogenic Vibrio aestuarianus strain 01/32 on lethality and cellular immune responses of the oyster Crassostrea gigas. Dev. Comp. Immunol. 2006, 30, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Liu, H.; Fu, S.; Dong, W.; Bu, W. Transcriptome-wide identification and characterization of Toll-like receptors response to Vibrio anguillarum infection in Manila clam (Ruditapes philippinarum). Fish Shellfish Immun. 2021, 111, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Kehlet-Delgado, H.; Hse, C.; Mueller, R.S. Comparative genomic analysis of Vibrios yields insights into genes associated with virulence towards C. gigas larvae. BMC Genom. 2020, 21, 599. [Google Scholar] [CrossRef]
- Liu, P.C.; Chen, Y.C.; Huang, C.Y.; Lee, K.K. Virulence of Vibrio parahaemolyticus isolated from cultured small abalone, Haliotis diversicolor supertexta, with withering syndrome. Lett. Appl. Microbiol. 2000, 31, 433–437. [Google Scholar] [CrossRef]
- Le Roux, F.; Wegner, K.M.; Polz, M.F. Oysters and Vibrios as a Model for Disease Dynamics in Wild Animals. Trends Microbiol. 2016, 24, 568–580. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Fuxa, J.R.; Lacey, L.A.; Onstad, D.W.; Kaya, H.K. Definitions of pathogenicity and virulence in invertebrate pathology. J. Invertebr. Pathol. 2005, 184, 1–7. [Google Scholar] [CrossRef]
- Casadevall, A.; Pirofski, L.-A. Host-Pathogen Interactions: The Attributes of Virulence. J. Infect. Dis. 2001, 184, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A.; Pirofski, L.-A. Host-Pathogen Interactions: Redefining the Basic Concepts of Virulence and Pathogenicity. Infect. Immun. 1999, 67, 3703–3713. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A.; Pirofski, L.A. Virulence factors and their mechanisms of action: The view from a damage-response framework. J. Water Health 2009, 7, S2. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zheng, D.; Zhou, S.; Chen, L.; Yang, J. VFDB 2022: A general classification scheme for bacterial virulence factors. Nucleic Acids Res. 2022, 50, D912–D917. [Google Scholar] [CrossRef] [PubMed]
- Frans, I.; Michiels, C.W.; Bossier, P.; Willems, K.A.; Lievens, B.; Rediers, H. Vibrio anguillarum as a fish pathogen: Virulence factors, diagnosis and prevention. J. Fish Dis. 2011, 34, 643–661. [Google Scholar] [CrossRef]
- Ceccarelli, D.; Hasan, N.A.; Huq, A.; Colwell, R.R. Distribution and dynamics of epidemic and pandemic Vibrio parahaemolyticus virulence factors. Front. Cell. Infect. Microbiol. 2013, 3, 97. [Google Scholar] [CrossRef] [PubMed]
- Osei-Adjei, G.; Huang, X.; Zhang, Y. The extracellular proteases produced by Vibrio parahaemolyticus. World J. Microb. Biot. 2016, 34, 68. [Google Scholar] [CrossRef]
- Natrah, F.M.I.; Ruwandeepika, H.A.D.; Pawar, S.; Karunasagar, I.; Sorgeloos, P.; Bossier, P.; Defoirdt, T. Regulation of virulence factors by quorum sensing in Vibrio harveyi. Vet. Microbiol. 2011, 154, 124–129. [Google Scholar] [CrossRef]
- Choi, G.; Choi, S.H. Complex regulatory networks of virulence factors in Vibrio vulnificus. Trends Microbiol. 2022, 30, 1205–1216. [Google Scholar] [CrossRef]
- Nurhafizah Wan Ibrahim, W.; Kok Leong, L.; Abdul Razzak, L.; Musa, N.; Danish Daniel, M.; Catherine Zainathan, S.; Musa, N. Virulence properties and pathogenicity of multidrug-resistant Vibrio harveyi associated with luminescent vibriosis in Pacific white shrimp, Penaeus vannamei. J. Invertebr. Pathol. 2021, 186, 107594. [Google Scholar] [CrossRef] [PubMed]
- Vanmaele, S.; Defoirdt, T.; Cleenwerck, I.; Vos, P.D.; Bossier, P. Characterization of the virulence of Harveyi clade vibrios isolated from a shrimp hatchery in vitro and in vivo, in a brine shrimp (Artemia franciscana) model system. Aquaculture 2015, 435, 28–32. [Google Scholar] [CrossRef]
- Defoirdt, T.J. Virulence mechanisms of bacterial aquaculture pathogens and antivirulence therapy for aquaculture. Rev. Aquacult. 2013, 5, 100–114. [Google Scholar] [CrossRef]
- Maeda, H.; Yamamoto, T. Pathogenic Mechanisms Induced by Microbial Proteases in Microbial Infections. Chem. Hoppe-Seyler 1996, 377, 217–226. [Google Scholar]
- Lee, K.-K.; Yu, S.-R.; Liu, P.-C. Alkaline serine protease is an exotoxin of Vibrio alginolyticus in kuruma prawn, Penaeus japonicus. Curr. Microbiol. 1997, 34, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Saulnier, D.; Decker, S.D.; Haffner, P.; Cobret, L.; Robert, M.; Garcia, C. A Large-Scale Epidemiological Study to Identify Bacteria Pathogenic to Pacific Oyster Crassostrea gigas and Correlation Between Virulence and Metalloprotease-like Activity. Microb. Ecol. 2010, 59, 787–798. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Zhang, X.-H.; Tang, X.; Wang, S.; Zhong, Y.; Chen, J.; Austin, B. A Single Residue Change in Vibrio harveyi Hemolysin Results in the Loss of Phospholipase and Hemolytic Activities and Pathogenicity for Turbot (Scophthalmus maximus). J. Bacteriol. 2007, 189, 2479–2575. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Liu, P.C.; Lee, K.K.; Chen, S.N. Pathogenicity of different isolates of Vibrio harveyi in tiger prawn, Penaeus monodon. Lett. Appl. Microbiol. 1996, 22, 413–416. [Google Scholar]
- Vogan, C.L.; Costa-Ramos, C.; Rowley, A.F. Shell disease syndrome in the edible crab, Cancer pagurus—Isolation, characterization and pathogenicity of chitinolytic bacteria. Microbiology 2002, 148, 743–754. [Google Scholar] [CrossRef][Green Version]
- Berlin, K.; Koren, S.; Chin, C.-S.; Drake, J.P.; Landolin, J.M.; Phillippy, A.M. Assembling large genomes with single-molecule sequencing and locality-sensitive hashing. Nat. Biotechnol. 2015, 33, 623–630. [Google Scholar] [CrossRef]
- Chin, C.S.; Alexander, D.H.; Marks, P.; Klammer, A.A.; Drake, J.; Heiner, C.; Clum, A.; Copeland, A.; Huddleston, J.; Eichler, E.E.; et al. Nonhybrid, finished microbial genome assemblies from long-read SMRT sequencing data. Nat. Methods 2013, 10, 563. [Google Scholar] [CrossRef]
- Delcher, A.L.; Bratke, K.A.; Powers, E.C.; Salzberg, S.L. Identifying bacterial genes and endosymbiont DNA with Glimmer. Bioinformatics 2007, 23, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Han, N.; Qiang, Y.; Zhang, W. ANItools web: A web tool for fast genome comparison within multiple bacterial strains. Database-Oxford 2016, 2016, baw084. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef]
- The Gene Ontology Consortium. Expansion of the Gene Ontology knowledgebase and resources. Nucleic Acids Res. 2017, 45, D331–D338. [Google Scholar] [CrossRef]
- Chen, L.; Yang, J.; Yu, J.; Yao, Z.; Sun, L.; Shen, Y.; Jin, Q. VFDB: A reference database for bacterial virulence factors. Nucleic Acids Res. 2005, 33, D325–D328. [Google Scholar] [CrossRef]
- Urban, M.; Pant, R.; Raghunath, A.; Irvine, A.G.; Pedro, H.; Hammond-Kosack, K.E. The Pathogen-Host Interactions database (PHI-base): Additions and future developments. Nucleic Acids Res. 2014, 43, 645–655. [Google Scholar] [CrossRef]
- Urban, M.; Cuzick, A.; Seager, J.; Wood, V.; Rutherford, K.; Venkatesh, S.Y.; Sahu, J.; Iyer, S.V.; Khamari, L.; Silva, N.D.; et al. PHI-base in 2022: A multi-species phenotype database for Pathogen–Host Interactions. Nucleic Acids Res. 2022, 50, 837–847. [Google Scholar] [CrossRef]
- Anderle, M.; Roy, S.; Lin, H.; Becker, C.; Joho, K. Quantifying reproducibility for differential proteomics: Noise analysis for protein liquid chromatography-mass spectrometry of human serum. Bioinformatics 2004, 20, 3575–3582. [Google Scholar] [CrossRef] [PubMed]
- Sandin, M.; Teleman, J.; Johan, M.; Levander, F. Data processing methods and quality control strategies for label-free LC-MS protein quantification. Biochim. Biophys. Acta 2014, 1844, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Madec, S.; Pichereau, V.; Jacq, A.; Paillard, M.; Boisset, C.; Guérard, F.; Paillard, C.; Nicolas, J.-L. Characterization of the secretomes of two vibrios pathogenic to mollusks. PLoS ONE 2014, 9, e113097. [Google Scholar] [CrossRef] [PubMed]
- Nottage, A.S.; Birkbeck, T.H. Purification of a proteinase produced by the bivalve pathogen Vibrio alginolyticus NCMB 1339. J. Fish Dis. 1987, 10, 211–220. [Google Scholar] [CrossRef]
- Rameshwaram, N.R.; Singh, P.; Ghosh, S.; Mukhopadhyay, S. Lipid metabolism and intracellular bacterial virulence: Key to next-generation therapeutics. Future Microbiol. 2018, 13, 1301–1328. [Google Scholar] [CrossRef] [PubMed]
- Guerra, P.R.; Herrero-Fresno, A.; Pors, S.E.; Ahmed, S.; Wang, D.; Thøfner, I.; Antenucci, F.; Olsen, J.E. The membrane transporter PotE is required for virulence in avian pathogenic Escherichia coli (APEC). Vet. Microbiol. 2018, 216, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Smith, I. Mycobacterium tuberculosis Pathogenesis and Molecular Determinants of Virulence. Clin. Microbiol. Rev. 2003, 16, 463–496. [Google Scholar] [CrossRef]
- Sherman, D.R.; Mdluli, K.; Hickey, M.J.; Arain, T.M.; Morris, S.L.; Barry, C.E., III; Stover, C.K. Compensatory ahpC gene expression in isoniazid-resistant Mycobacterium tuberculosis. Science 1996, 272, 1641–1643. [Google Scholar] [CrossRef]
- Li, Z.; Kelley, C.; Collins, F.; Rouse, D.; Morris, S. Expression of katG in Mycobacterium tuberculosis Is Associated with Its Growth and Persistence in Mice and Guinea Pigs. J. Infect. Dis. 1998, 177, 1030–1035. [Google Scholar] [CrossRef]
- Zhang, M.; Yan, Q.; Mao, L.; Wang, S.; Huang, L.; Xu, X.; Qin, Y. KatG plays an important role in Aeromonas hydrophila survival in fish macrophages and escape for further infection. Gene 2018, 672, 156–164. [Google Scholar] [CrossRef]
- Hagman, K.E.; Shafer, W.M. Transcriptional control of the mtr efflux system of Neisseria gonorrhoeae. J. Bacteriol. 1995, 177, 4162–4165. [Google Scholar] [CrossRef] [PubMed]
- Jerse, A.E.; Sharma, N.D.; Simms, A.N.; Crow, E.T.; Snyder, L.A.; Shafer, W.M. A Gonococcal Efflux Pump System Enhances Bacterial Survival in a Female Mouse Model of Genital Tract Infection. Infect. Immun. 2003, 71, 5576–5582. [Google Scholar] [CrossRef] [PubMed]
- Miller, V.L.; Taylor, R.K.; Mekalanos, J.J. Cholera toxin transcriptional activator ToxR is a transmembrane DNA binding protein. Cell 1987, 48, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Milton, D.L. Quorum sensing in vibrios: Complexity for diversification. Int. J. Med. Microbiol. 2006, 296, 61–71. [Google Scholar] [CrossRef]
- Ruwandeepika, H.A.D.; Defoirdt, T.; Bhowmick, P.P.; Karunasagar, I.; Karunasagar, I.; Bossier, P. In vitro and in vivo expression of virulence genes in Vibrio isolates belonging to the Harveyi clade in relation to their virulence towards gnotobiotic brine shrimp (Artemia franciscana). Environ. Microbiol. 2010, 13, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Rønneseth, A.; Castillo, D.; D’Alvise, P.; Tønnesen, Ø.; Haugland, G.; Grotkjær, T.; Engell-Sørensen, K.; Nørremark, L.; Bergh, Ø.; Wergeland, H.I.; et al. Comparative assessment of Vibrio virulence in marine fish larvae. J. Fish Dis. 2017, 40, 1373–1385. [Google Scholar] [CrossRef] [PubMed]
- Saleh, T.H.; Sabbah, M.A.; Jasem, K.A.; Hammad, Z.N. Identification of virulence factors in Vibrio cholerae isolated from Iraq during the 2007-2009 outbreak. Can. J. Microbiol. 2011, 57, 1024–1031. [Google Scholar] [CrossRef] [PubMed]
- Gerganova, V.; Berger, M.; Zaldastanishvili, E.; Sobetzko, P.; Lafon, C.; Mourez, M.; Travers, A.; Muskhelishvili, G. Chromosomal position shift of a regulatory gene alters the bacterial phenotype. Nucleic Acids Res. 2015, 43, 8215–8226. [Google Scholar] [CrossRef]
- Salyers, A.A.; Gupta, A.; Wang, Y. Human intestinal bacteria as reservoirs for antibiotic resistance genes. Trends Microbiol. 2004, 12, 412–416. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).