1. Introduction
In modern pig production, piglets are weaned at 3~4 weeks, which is much earlier than the weaning age in the normal state [
1]. Due to changes in the environment, social structure, and food sources, weaned piglets often encounter a variety of stressors, which can culminate in diarrhea, intestinal inflammation and pathogenic bacteria infection and which pose a threat to the health of piglets [
2]. Diarrhea of weaned piglets is the main cause of growth performance decline and even death [
3]. Due to the insufficient secretion of stomach acid and digestive enzymes at weaning, nutrients cannot be fully digested, the source of lactose is reduced, and the content of lactic acid is decreased; this can lead to diarrhea caused by indigestion [
4]. Additionally, the levels of maternal antibodies decline after weaning, leaving piglets with an immature immune system and making them vulnerable to infection from pathogenic bacteria. This susceptibility often results in the onset of diarrhea [
5]. The gastrointestinal tract, or gut, serves multiple essential functions in piglets. It facilitates the digestion and absorption of nutrients while acting as a critical barrier against external factors and potential diseases. The structural integrity and proper functioning of the intestines significantly influence the overall health of piglets. After weaning, the intestinal microecological environment of piglets undergoes significant changes. The composition of the intestinal microbial community plays a crucial role in maintaining the overall health of the host. Therefore, promoting the healthy development of the intestine is a paramount task during the weaning stage of piglets and is integral to their overall health.
Zn is a highly essential trace element for the control of several physiological functions, including animal growth and health, immune system, and bone development, since it is a component of more than 300 different enzymes and proteins [
6,
7]. Zinc oxide (ZnO) is often used as one of the sources of zinc in feed; the pharmacological dose of ZnO is used as a means to control diarrhea and improve the growth performance in piglets [
8]. This supplementation has been found to have a notable impact on the intestinal morphological development of piglets, bolster their immune function within the intestines, and facilitate the colonization of beneficial bacteria. Currently, zinc oxide is incorporated into feed in different forms, including coated with zinc oxide and nano-zinc oxide. However, studies have found that long-term feeding of high-dose zinc oxide can have adverse effects on the immune metabolism and intestinal function of piglets, and cause ZnO-dependent diarrhea, as well as soil and water pollution [
9]. Therefore, more low-zinc piglet diets and zinc oxide substitutes have been investigated [
10]. Tetrabasic zinc chloride (TBZC) is an inorganic zinc prepared by the crystallization process, which was officially approved as a feed additive in 2003. Its chemical properties are more stable and can reduce the risk of oxidation of nutrients during dietary storage. Compared with ZnO, TBZC has higher relative bioavailability and better palatability [
11], and contains lower heavy metal impurities compared to other conventional inorganic zinc. It has been reported that the addition of TBZC to feed rations in place of other zinc additives such as ZnO significantly improved the growth performance of fattening pigs [
12]. In previous experiments, the addition of 1000 mg/kg TBZC to the diet of piglets resulted in significant improvements in growth performance and nutrient digestibility [
13]. Building on these findings, the present study aimed to investigate the effects of replacing zinc oxide with TBZC on intestinal morphology, immune status, and intestinal microbes of weaned piglets. To accomplish this, three treatment groups were designed in this experiment, receiving supplementation of 800, 1000, and 1200 mg/kg TBZC.
2. Materials and Methods
The experimental design and procedures used in this study were approved by the animal ethical committee of China Agricultural University (Beijing, China; No. AW11104202-1-3) according to the Chinese Guidelines for Animal Welfare. The experiment was conducted at the FengNing Swine Research Unit of China Agricultural University (Chengdejiuyun Agricultural and Livestock Co., Ltd., Chengde, China).
2.1. Animals and Experimental Designs
A total of 240 piglets (Duroc × Landrace × Yorkshire, BW = 6.59 ± 0.05 kg) weaned at 21 d of age were randomly divided into 5 treatments based on body weight (BW) and sex, and each treatment included 6 replicate pens and 8 piglets per pen. The 5 treatments included a control diet: (CON) control group; (T1) CON + 1500 mg/kg zinc in ZnO; (T2) CON + 800 mg/kg zinc in TBZC; (T3) CON + 1000 mg/kg zinc in TBZC; (T4) CON + 1200 mg/kg zinc in TBZC. The ZnO and TBZC used in this study were provided by Xinjia Bio-Engineeriong Co., Ltd. (Changsha, China).
The non-medicated corn-soybean basal diets were formulated to meet requirements recommended by the National Research Council (NRC, 2012) for 5 to 7 and 7 to 11 kg BW pigs and are shown in
Table 1.
2.2. Feeding and Management
All piglets were housed in a temperature-controlled nursery (temperature 26~28 °C; humidity 55%~70%) and had ad libitum access to feed and water for 28 days. Feeders were checked and additional feed was added as needed to achieve ad libitum access at 08:30 and 15:30 daily, and their daily feed intake, feces, diarrhea situation, and feed intake were observed and recorded.
2.3. Diarrhea Scores
The anus of pigs was examined every day during the experimental period, and the number of pigs with diarrhea was observed and counted. The scoring system was applied to determine the rate of diarrhea as follows: 0 = hard feces; 1 = slightly soft feces; 2 = soft, partially formed feces; 3 = loose, semiliquid feces; 4 = watery, mucous-like feces. When the average score was over 2 for 2 consecutive days, pigs were identified as having diarrhea [
14]. The diarrhea rate was calculated according to the following equation:
2.4. Sample Collections
During this experiment, feed samples of each treatment were collected at the beginning of this study and stored at −20 °C. On days 12 to 14 and 26 to 28, fresh fecal samples were collected from each pen. After mixing, the representative fresh fecal samples were baked at 65 °C for 72 h, rehumidified at natural conditions for 24 h, ground passed through a 40-mesh sieve, and stored at −20 °C until analysis.
On day 15, 6 pigs from each treatment group close to the median BW were selected and euthanized. The small intestinal tracts were taken from the abdominal cavity, and divided according to physiological features into three parts: duodenum, jejunum, and ileum [
15]. A 2 cm segment from each of duodenum, jejunum and ileum were taken, and the intestinal specimens were washed with normal saline and stored in 4% paraformaldehyde for 24 h for morphological examination. Jejunum, ileum, cecum, and colon contents were collected in cryopreserved tubes and immediately frozen in liquid nitrogen and stored at −80 °C until testing for microbial composition analysis. The stomach was cut open during dissection, and the pH of the stomach contents was measured using the pH detector, then the jejunum and ileum were selected, and the contents were placed in a paper cup, and the metal probe of the detector was completely submerged in the contents for 3 s and then the values were recorded after stabilization. Then, the remaining duodenum, jejunum, and ileum segments were rinsed with cold normal saline, the intestinal segments were cut completely, the mucosal layer was carefully scraped off with slides, and the segments were immediately frozen in liquid nitrogen and stored at −80 °C for detection of IgG and IgA.
2.5. Chemical Analysis for Diet and Feces
Diets and feces were analyzed for crude protein (CP) and chromium using AOAC methods [
16]. ATTD of nutrients is as follows:
2.6. Histological Analysis
Intestinal tissues were stored in 4% paraformaldehyde at room temperature. The samples were cut into small pieces with a width of 2 cm. Intestinal samples were dehydrated and dealcoholized, embedded in paraffin, and sectioned. The slides were deparaffinized, rehydrated, and subjected to H&E staining. Samples were examined using an Olympus IX51 inverted phase contrast microscope.
2.7. Immunity of the Intestinal Tract
Firstly, IgG, IgA, and IgM in the intestinal mucosa were detected using an ELISA kit (Leibertech-nik, Beijing, China). Blank control wells were spiked with color developer A&B and termination solution, following the same steps as the other wells. A volume of 50 μL standard substance and streptavidin-HRP had been added into the standard pore. A volume of 40 μL of each sample was used for the sample wells, then 10 μL of secondary antibody and 50 μL of streptavidin-HRP were added to each well, covered with plate sealing membrane, gently shaken and mixed, and incubated at 37 °C for 60 min. The sealing plate film was removed, and the liquid was discarded. The plate was then shaken dry. Each well was filled with washing solution and allowed to stand for 30 s before discarding. This process was repeated five times. The plate was then patted dry. Subsequently, 50 μL of chromogenic agent and chromogenic agent B were added consecutively to each well and mixed. Finally, the color development was carried out at 37 °C for 10 min. A volume of 50 μL termination solution was added to each well, and the termination reaction was zero with blank conditioning. Measured the absorbance (OD) of each well sequentially at 450 nm. The measurement should be carried out within 10 min after the addition of the termination solution.
2.8. Microbiological Analysis of Intestinal Contents
Jejunum, ileum, cecum, and colon contents were collected in cryopreserved tubes and immediately stored at −80 °C in liquid nitrogen until testing for microbial composition analysis. Total microbial genomic DNA was extracted from the samples using the E.Z.N.A.
® soil DNA Kit (Omega Bio-tek, Norcross, GA, USA) according to the manufacturer’s instructions. The quality and concentration of DNA were determined by 1.0% agarose gel electrophoresis and a NanoDrop
® ND-2000 spectrophotometer (Thermo Scientific Inc., Waltham, MA, USA) and kept at −80 °C prior to further use. The hypervariable region V3-V4 of the bacterial 16S rRNA gene were amplified with primer pairs 338F (5’-ACTCCTACGGGAGGCAGCAG-3’) and 806R (5’-GGACTACHVGGGTWTCTAAT-3’) by an ABI GeneAmp
® 9700 PCR thermocycler (ABI, Los Angeles, CA, USA). The amplification cycles conditions were as follows: 95 °C pre-denaturation for 3 min, followed by 27 cycles of 95 °C denaturation for 30 s, 55 °C annealing for 30 s and 72 °C extending for 30 s, followed by 72 °C extending for 10 min. All samples were amplified in triplicate. The sequencing was performed using Illumina’s Miseq PE300 platform (Illumina, San Diego, CA, USA). The raw sequenced sequences were sequenced using fastp (
https://github.com/OpenGene/fastp, version 0.19.6, accessed on 16 March 2023) software and FLASH (
http://www.cbcb.umd.edu/software/flash, version 1.2.11, accessed on 16 March 2023) software for Quality control and splicing. The splicing was performed for subsequent analysis. Then, using the UPARSE software (
http://drive5.com/uparse/, version 7.1, accessed on 16 March 2023), the quality control spliced sequences were subjected to operational taxonomic unit (OTU) removal of chimeras and standard clustering (with 97% confidence level) based on 97% similarity, then the representative OTUs sequences were selected for annotation. Finally, the composition of microorganisms in the gut contents was analyzed based on the standardized OTUs. The grouped samples were analyzed for differences in species composition and characteristic flora using Mothur software (
http://www.mothur.org/wiki/Calculators, accessed on 16 March 2023) and the results were presented in pictures.
2.9. Statistical Analysis
Bioinformatic analysis of the feces microbiota was carried out using the Majorbio Cloud platform (
https://cloud.majorbio.com, accessed on 16 March 2023). Based on the OTU information, the alpha diversity indices of Ace, Chao, and Shannon were calculated using Mother (
http://www.mothur.org/wiki/Calculators, accessed on 16 March 2023). Bray–Curtis dissimilarity-based principal coordinates analysis (PCoA) was used to determine the similarity between microbial communities in different samples. Linear discriminant analysis (LDA) effect size (LEfSe) (
http://huttenhower.sph.harvard.edu/LEfSe, accessed on 16 March 2023) was used to analyze bacterial taxa (phylum to species) that differed significantly between groups (LDA score > 2,
p < 0.05). Differences in diarrhea rates were analyzed with the Pen (replicate) as the experimental unit. Piglets were used as experimental units to analyze crude protein digestibility, intestinal morphology, intestinal immunity and intestinal flora. Statistical analysis was performed using the unpaired t-test procedure in SAS 9.4 statistical software, ANOVA was used for analysis of variance, and Tukey’s test was used for multiple comparisons.
p < 0.05 indicates a significant difference; 0.05 <
p < 0.1 indicates a tendency to have a significant impact.
4. Discussion
Studies have shown that the addition of pharmacological doses of zinc to weaned piglet diets can alleviate weaning stress and improve growth performance [
17,
18,
19,
20,
21]. In addition to its inhibitory effect on piglet growth performance, long-term feeding of ZnO caused dependent diarrhea and soil contamination [
22]. TBZC is a novel inorganic source of Zn manufactured through the reaction of high purified forms of the metal with water and hydrochloric acid [
11]. It has a crystalline structure made up of covalent connections between the soluble metal ion, many hydroxyl groups, and chloride ions [
23]. Mavromichalis et al. showed that weaned piglets fed 1500 mg/kg of TBZC could achieve the same growth-promoting effect as high-dose zinc oxide and were not affected by the addition of antibacterial agents, effectively improving feed conversion efficiency, the growth performance of piglets remained unaffected when TBZC was administered at levels lower than 1500 mg/kg. Furthermore, it was observed that the inclusion of 1200 mg/kg TBZC did not yield any improvements in growth performance [
12]. However, the growth performance of weaned pigs decreased to the same level as the control group when TBZC was supplemented at 3000 mg/kg, which is consistent with previous report that pigs fed 3000 mg/kg Zn-deficient treatment [
24,
25].
Since the digestive system of weaned piglets is not fully developed, the digestibility of CP may be affected by a variety of factors; the composition of the diet, the development of the subjects, and the use of feed additives will all affect the apparent digestibility of nutrients [
26]. Supplementation of 1200 mg/kg TBZC in the diet could effectively improve the digestion and absorption ability of crude protein in weaned piglets, this result is consistent with previous studies, piglets fed the TBZC diet had higher (
p < 0.05) digestibility of crude protein and gross energy than those fed the CON diet [
26]. Swine protein digestion starts in the stomach with the aid of the pepsin enzyme, which is created by the major stomach cells and transforms from an inactive form to pepsin in an acidic environment [
13].
The low pH is necessary for the conversion of gastric zymogen into active enzymes. The pH of the stomach contents of the weaned piglets was higher than that of the lactation period. The increased acidity of the digestive tract affects the activity of intestinal digestive enzymes and causes proliferation of pathogenic bacteria. The feeds’ high buffering/binding capacity contributes to even higher stomach pH, it may alter gastrointestinal pH, which may have an effect on how proteins are digested and health of the gut flora [
27]. The acidic properties of TBZC are comparatively lower than that of ZnO. As a result, the degree of stomach acid or acidifier consumption is also lower when using TBZC compared to ZnO. The results indicate that the addition of TBZC in the supplementation leads to a lower pH value in the stomach contents compared to the ZnO supplementation. This observation further supports the idea that TBZC provides better protection for the digestive environment in the stomach.
The small intestine is the site for digestion and absorption of nutrients in pigs. Villus height, crypt depth, and villus height to crypt depth ratio directly reflect the health of the small intestine [
28]. The intestinal villi atrophy as the epithelium decreases, and the crypts deepen as cell differentiation decreases [
29]. In this study, the addition of 1200 mg/kg of TBZC to the feed significantly improved the ratio of villi height to crypt depth in the ileal intestinal segment (
p < 0.05), and there was a tendency to decrease the depth of duodenal and ileal crypt. These results indicate that TBZC has an improvement effect on intestinal morphology. However, this experiment also demonstrated that the addition of TBZC did not significantly affect the morphology of the jejunum. Previous studies have also reported that high doses of zinc did not have a significant impact on intestinal morphology at the jejunum stage of piglets [
30], which aligns with the findings of this paper. Consequently, further investigation is required to fully understand the impact of TBZC on intestinal morphology.
In addition, Zinc deficiency can injure the intestinal mucosal barrier [
31]. It has been shown that treating Caco2 cells with TPEN for zinc deprivation led to decreased expression of tight junction proteins, increased intestinal epithelial permeability, and impaired intestinal mucosal barrier function [
32]. In this study, TBZC also tended to increase the IgA content in the duodenal and ileal mucosa, further demonstrating the positive effect of TBZC on improving the intestinal barrier. With the decrease in intestinal permeability, it also hinders the transport of harmful bacteria or toxins to a certain extent, further reducing the incidence of diarrhea and enteritis [
33]. Intestinal immune indicators can reflect the health status of the intestinal tract. The results of this experiment indicate the effects of dietary supplementation with 1000 mg/kg of TBZC on the IgA content in the ileum mucosa. Although there was a noticeable increase, it did not reach statistical significance. The levels of immunoglobulin in the duodenum and jejunum mucosa were not significantly impacted. Hence, in order to gain a better understanding of the effects of TBZC on intestinal immunity, further exploration and comprehensive studies are necessary.
The colony composition and microbial diversity of the microbial community in the gut are closely related to the health of the host, which can ensure the stability and health of the intestinal function, and have a significant role in maintaining the stability of the intestinal ecosystem and reducing the stress caused by the external environment [
34]. High doses of zinc can effectively inhibit the growth of harmful bacteria in the intestine [
35], decrease the utilization of nutrients in the feed by microorganisms, reduce the nutrient antagonism between intestinal microorganisms and the host, thereby improving feed conversion efficiency in weaned piglets [
21]. Feeding 2500 mg/kg ZnO to weaned piglets could effectively increase the microbial diversity of intestinal digestion [
36]. The experimental study revealed that TBZC has the potential to enhance microbial diversity in the intestinal contents of piglets. As the microbial community becomes more diverse, there is an increase in the relative abundance of beneficial bacteria, which may contribute to improvements in piglets’ diarrhea, digestion and absorption capabilities, as well as their overall intestinal health [
10]. This is consistent with the conclusion of this study that TBZC could effectively increase the abundance of beneficial bacteria and reduce the abundance of harmful bacteria in the gut of weaned pigs. However, long-term exposure to a high-zinc diet can lead to poisoning of weaned piglets, change the integrity of intestinal mucosal epithelial structure, reduce growth performance and intestinal mucosal immune function [
37]. Thus, the lower dose of TBZC caused less intestinal irritation and fewer side effects in weanling piglets than ZnO.
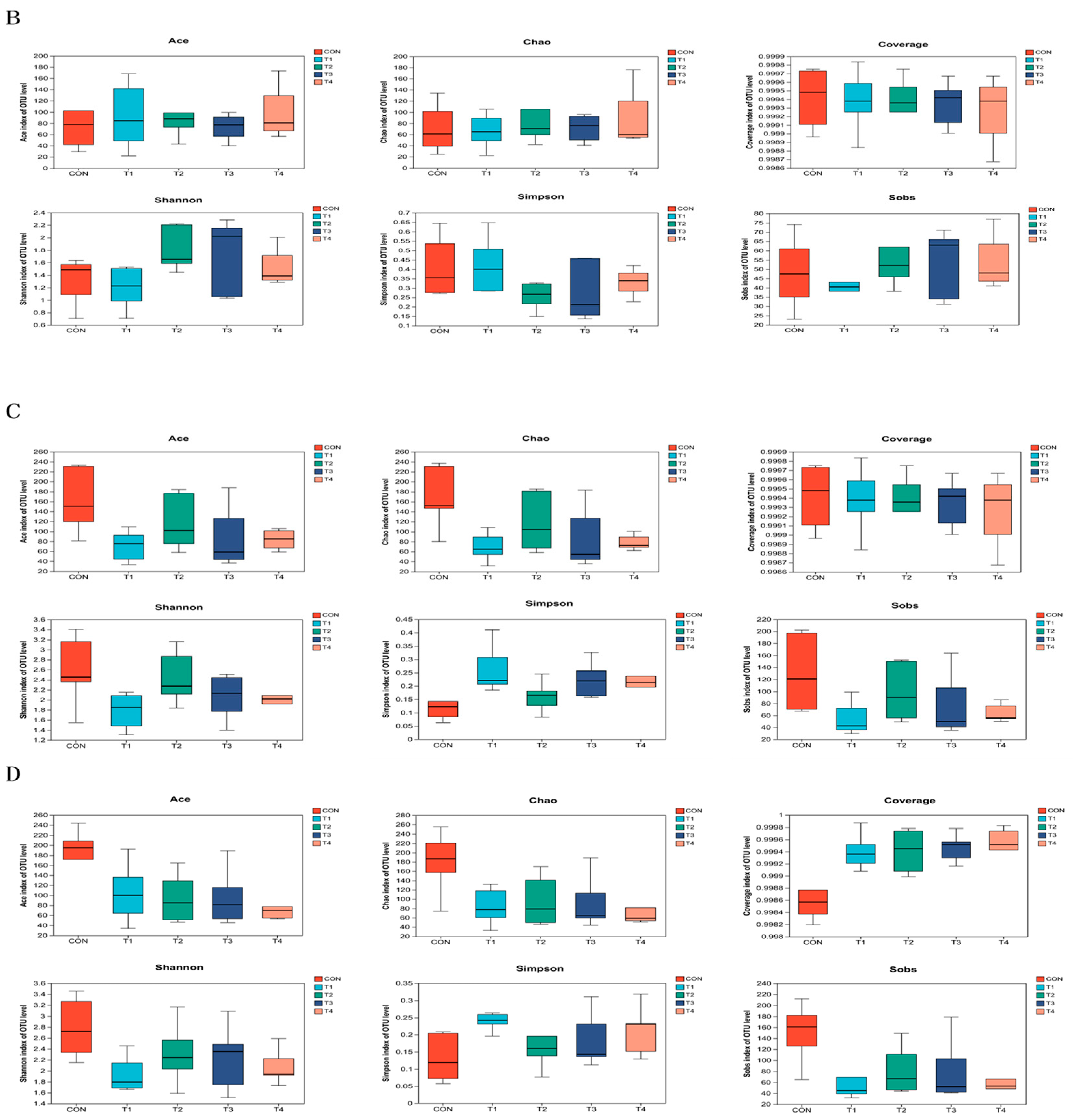
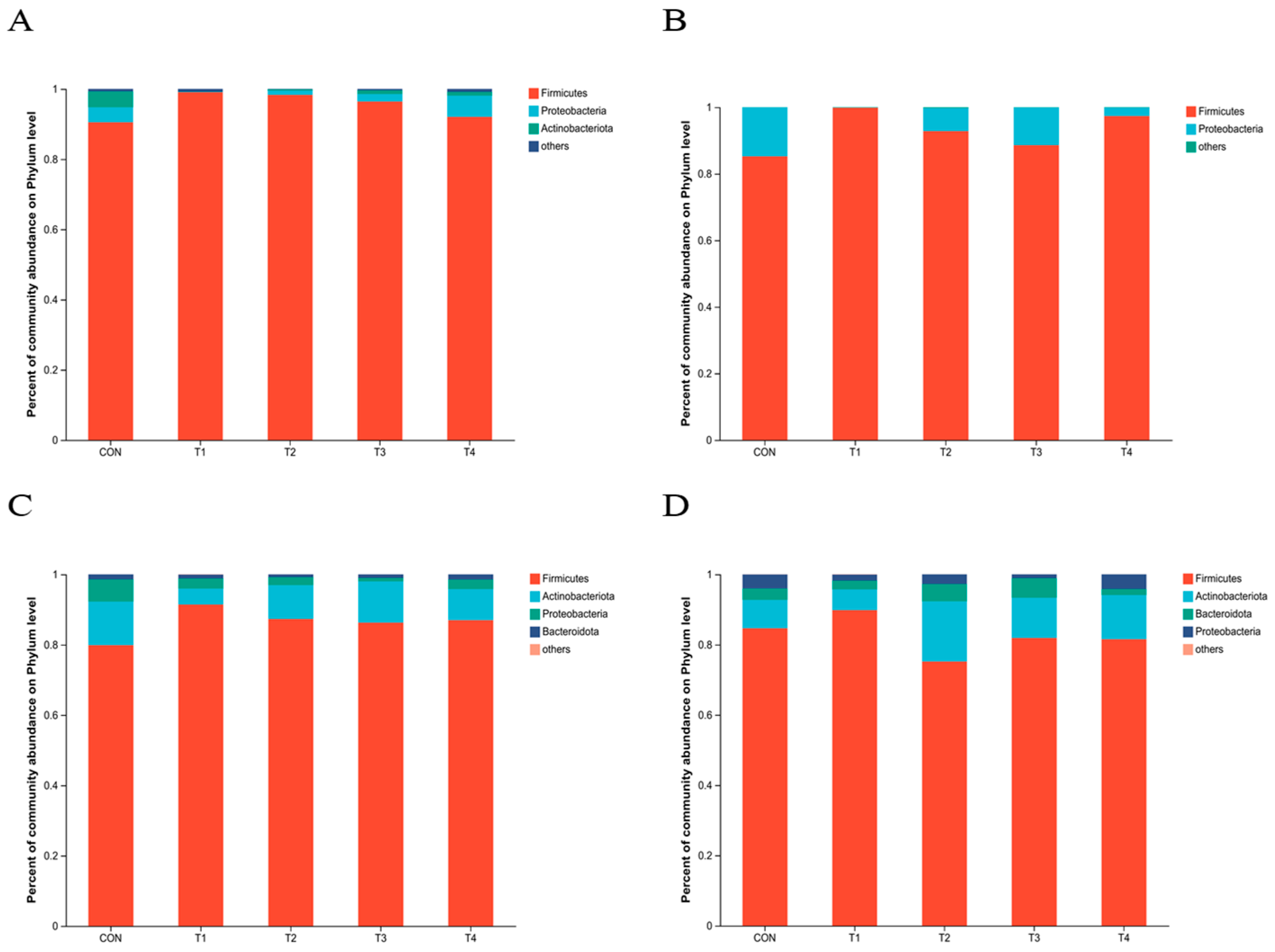
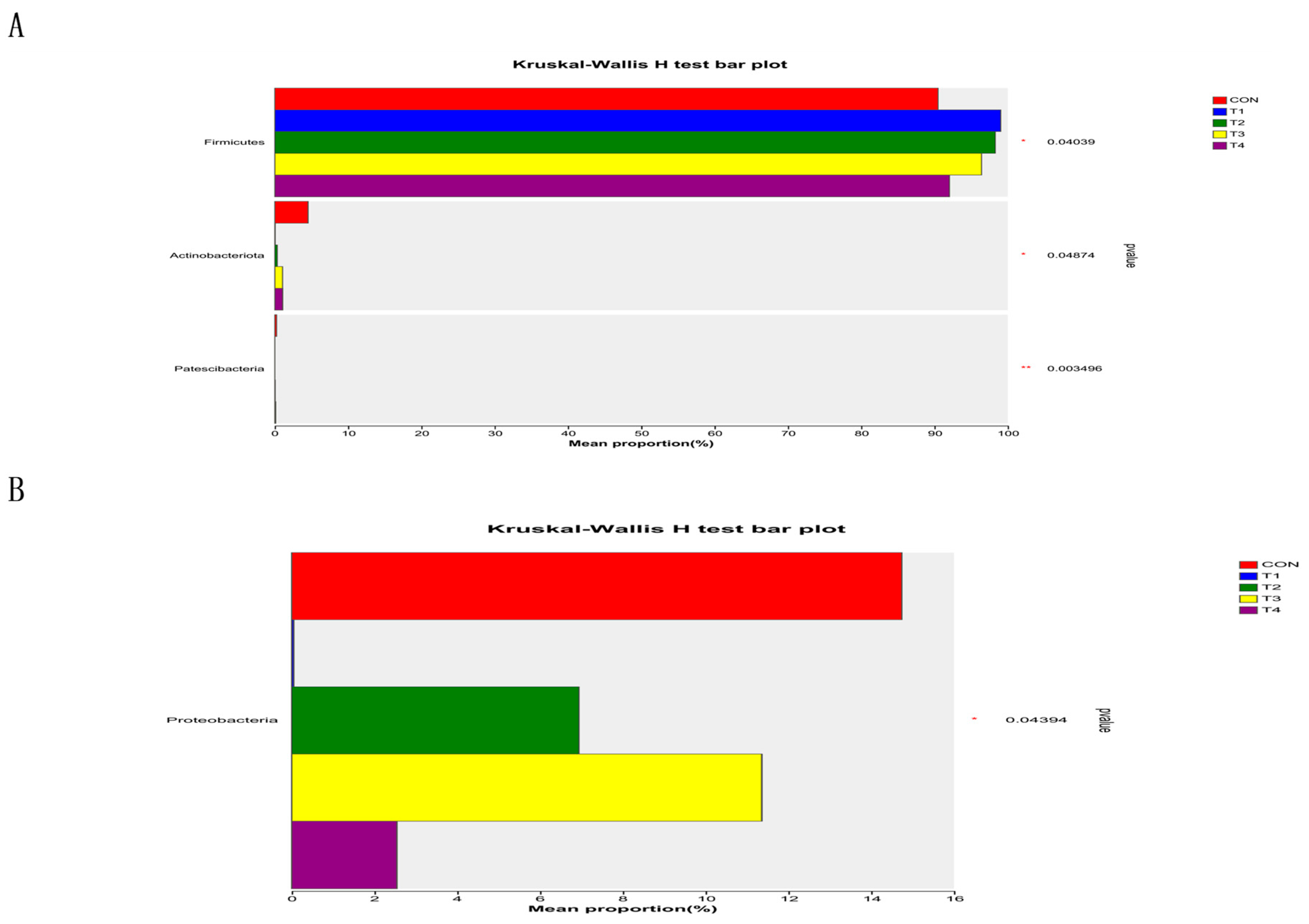
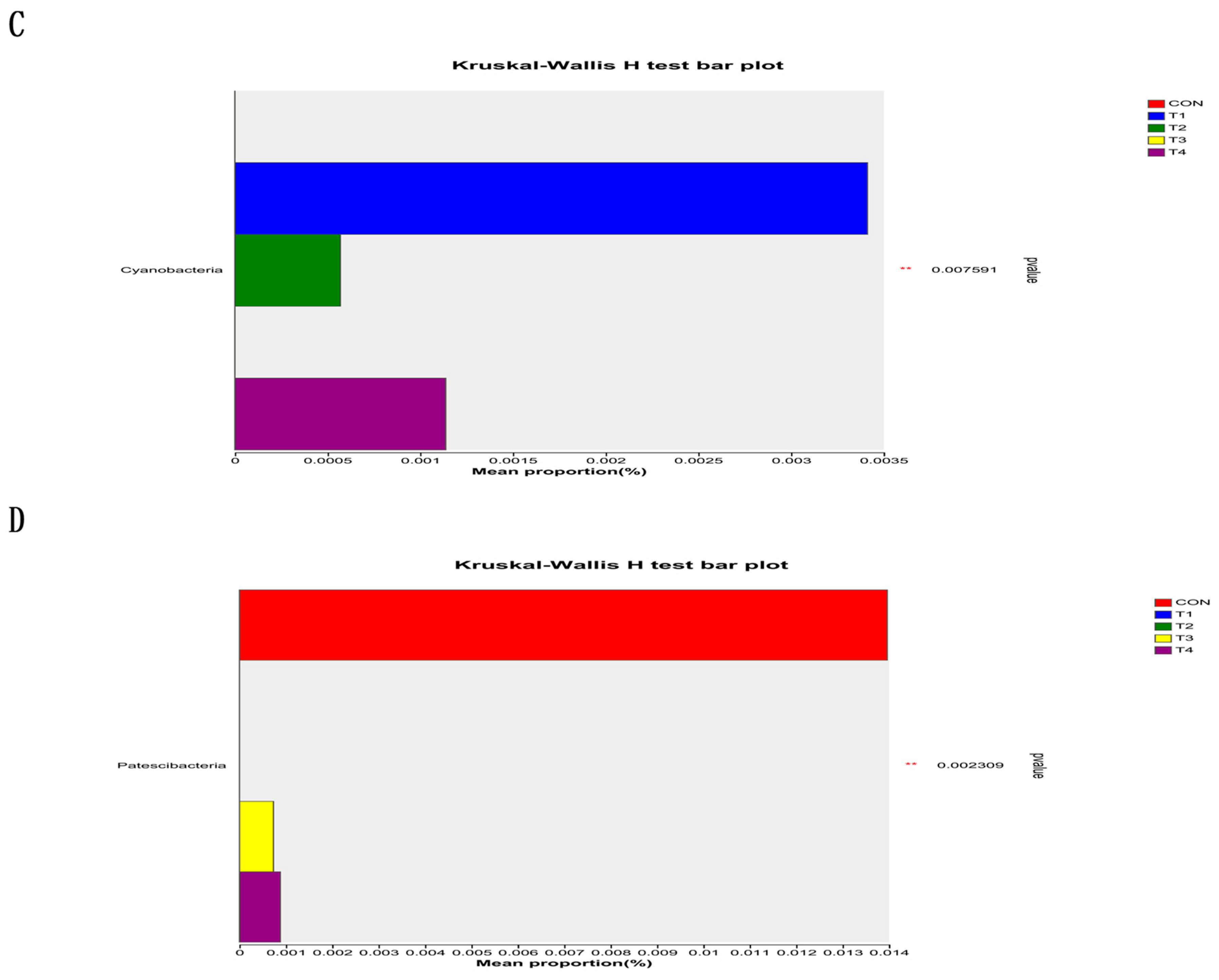
This study demonstrated that the addition of various zinc sources had a notable impact on microbial alpha diversity in the jejunum contents. It can be inferred that zinc supplementation can modulate the diversity of microorganisms in the jejunum, potentially affecting the overall gut health and function of the subjects. However, it is important to note that there was a higher Chao index and a lower Coverage index in the CON group compared to the other treatment groups. While these differences were not statistically significant, they suggest potential variations in the microbial diversity and abundance between the CON group and the other treatment groups. TBZC was also found to significantly increase the relative abundance of Firmicutes in the intestine, and Firmicutes contribute to energy uptake and storage in pigs. Increasing the relative abundance of Veillonellaceae can increase the concentration of propionic acid in food, promote the production of SCFA, and improve the immune stress of piglets, Negativicutes can effectively promote energy uptake and storage in weaned piglets. However, there are limitations in this study to exploring the effects of TBZC on intestinal health, microflora and immune stress of weaned piglets; the effects of different doses of TBZC on the function of host-specific flora are inconsistent with previous studies, and the reproducibility is low, so further investigation is still needed.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}