
A Genetic Tool to Identify Predators Responsible for Livestock Attacks in South America and Recommendations for Human–Wildlife Conflict Mitigation

 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Phase One: PCR Protocol Standardization
2.2. Phase Two: Testing the Standardized Protocols in Simulated Attacks
2.3. Phase Three: Use of Standardized Protocols to Analyze DNA from Bite Wounds Resulting from Attacks on Livestock in Rural Areas of Ecuador (Field Testing)
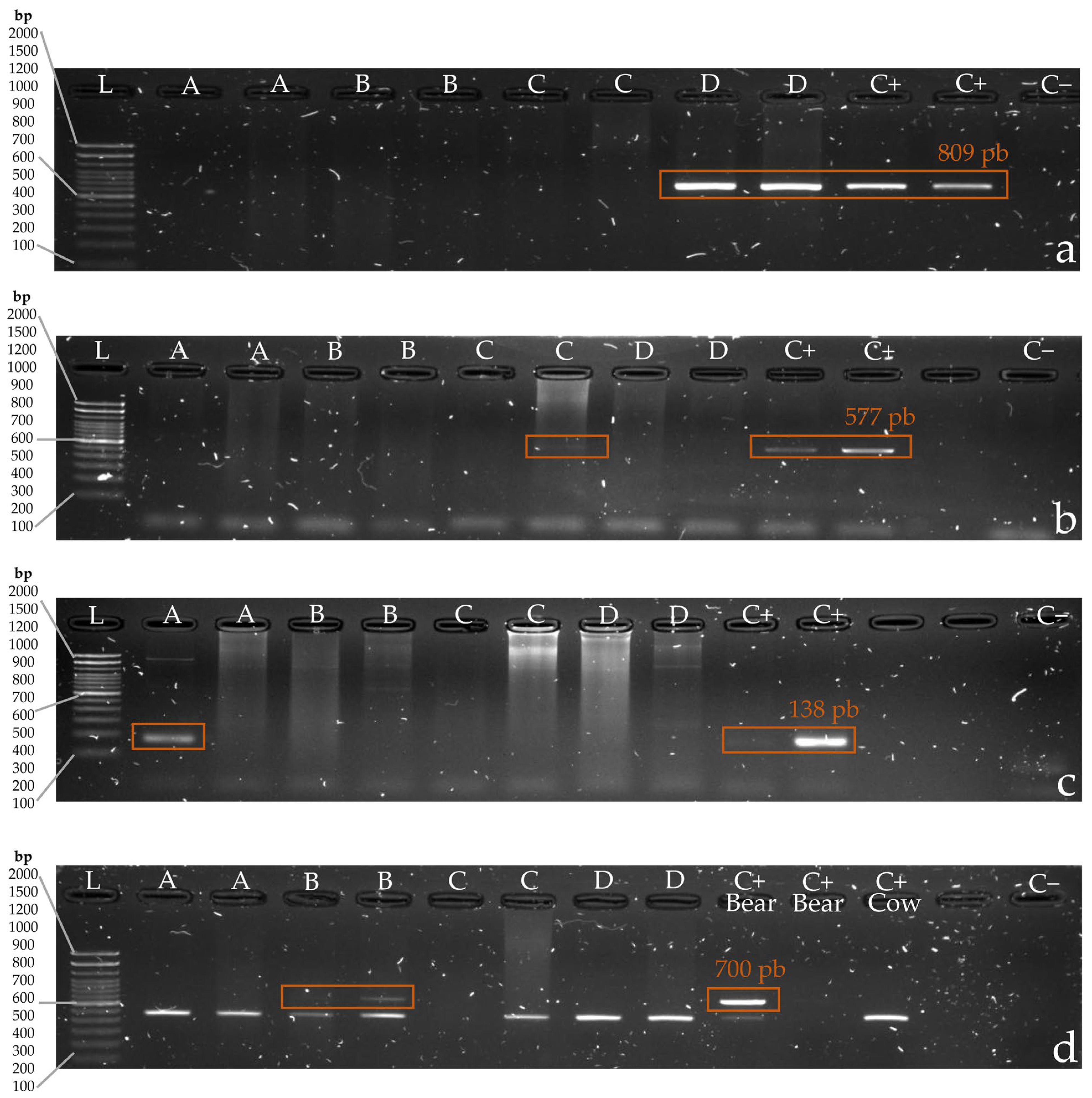
3. Results
3.1. Phase One: PCR Protocol Standardization
3.2. Phase Two: Testing the Standardized Protocols in Simulated Attacks
3.3. Phase Three: Use of Standardized Protocols to Analyze DNA from Bite Wounds Resulting from Attacks on Livestock in Rural Areas of Ecuador (Field Testing)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hughes, J.; Macdonald, D.W. A review of the interactions between free-roaming domestic dogs and wildlife. Biol. Conserv. 2013, 157, 341–351. [Google Scholar] [CrossRef]
- Headland, R.K. History of exotic terrestrial mammals in Antarctic regions. Polar Rec. 2012, 48, 123–144. [Google Scholar] [CrossRef]
- Gompper, M.E. The Dog-Human-Wildlife Interface: Assessing the Scope of the Problem. In Free-Ranging Dogs and Wildlife Conservation; Gompper, M.E., Ed.; Oxford University Press: Oxford, UK, 2014; pp. 9–54. [Google Scholar]
- Galibert, F.; Quignon, P.; Hitte, C.; André, C. Toward understanding dog evolutionary and domestication history. C. R. Biol. 2011, 334, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Doherty, T.S.; Dickman, C.R.; Glen, A.S.; Newsome, T.M.; Nimmo, D.G.; Ritchie, E.G.; Vanak, A.T.; Wirsing, A.J. The global impacts of domestic dogs on threatened vertebrates. Biol. Conserv. 2017, 210, 56–59. [Google Scholar] [CrossRef]
- Garde, E.; Acosta-Jamett, G.; Bronsvoort, B.M. Review of the risks of some canine zoonoses from free-roaming dogs in the post-disaster setting of Latin America. Animals 2013, 3, 855–865. [Google Scholar] [CrossRef]
- Canal, D.; Martín, B.; De Lucas, M.; Ferrer, M. Dogs are the main species involved in animal-vehicle collisions in southern Spain: Daily, seasonal and spatial analyses of collisions. PLoS ONE 2018, 13, e0203693. [Google Scholar] [CrossRef]
- Dhillon, J.; Hoopes, J.; Epp, T. Scoping decades of dog evidence: A scoping review of dog bite-related sequelae. Can. J. Public Health 2019, 110, 364–375. [Google Scholar] [CrossRef] [PubMed]
- Home, C.; Pal, R.; Sharma, R.K.; Suryawanshi, K.R.; Bhatnagar, Y.V.; Vanak, A.T. Commensal in conflict: Livestock depredation patterns by free-ranging domestic dogs in the Upper Spiti Landscape, Himachal Pradesh, India. Ambio 2017, 46, 655–666. [Google Scholar] [CrossRef] [PubMed]
- Plumer, L.; Talvi, T.; Männil, P.; Saarma, U. Assessing the roles of wolves and dogs in livestock predation with suggestions for mitigating human–wildlife conflict and conservation of wolves. Conserv. Genet. 2018, 19, 665–672. [Google Scholar] [CrossRef]
- Montecino-Latorre, D.; San Martín, W. Evidence supporting that human-subsidized free-ranging dogs are the main cause of animal losses in small-scale farms in Chile. Ambio 2019, 48, 240–250. [Google Scholar] [CrossRef]
- Lowenstein, C.; Waters, W.F.; Roess, A.; Leibler, J.H.; Graham, J.P. Animal husbandry practices and perceptions of zoonotic infectious disease risks among livestock keepers in a rural parish of Quito, Ecuador. Am. J. Trop. Med. Hyg. 2016, 95, 1450. [Google Scholar] [CrossRef] [PubMed]
- Torres, B.; Andrade, V.; Heredia, R.M.; Toulkeridis, T.; Estupiñán, K.; Luna, M.; Bravo, C.; García, A. Productive Livestock Characterization and Recommendations for Good Practices Focused on the Achievement of the SDGs in the Ecuadorian Amazon. Sustainability 2022, 14, 10738. [Google Scholar] [CrossRef]
- UN General Assembly. Transforming Our World: The 2030 Agenda for Sustainable Development. 2015. Available online: https://sustainabledevelopment.un.org/content/documents/21252030%20Agenda%20for%20Sustainable%20Development%20web.pdf (accessed on 29 November 2023).
- Bergman, D.L.; Breck, S.W.; Bender, S.C. Dogs Gone Wild: Feral Dog Damage in the United States. In Proceedings of the 13th Wildlife Damage Management Conference, Saratoga, NY, USA, 5 April 2009; Boulanger, J.R., Ed.; USDA National Wildlife Reasearch Center-Staff Publications: Fort Collins, CO, USA, 2009; pp. 177–183. [Google Scholar]
- Zapata-Ríos, G. Dogs are more than wet kisses and tail wags: Domestic dogs as invasive species. Anim. Conserv. 2018, 21, 287–288. [Google Scholar] [CrossRef]
- Sundqvist, A.K.; Ellegren, H.; Vila, C. Wolf or dog? Genetic identification of predators from saliva collected around bite wounds on prey. Conserv. Genet. 2008, 9, 1275–1279. [Google Scholar] [CrossRef]
- Caniglia, R.; Fabbri, E.; Mastrogiuseppe, L.; Randi, E. Who is who? Identification of livestock predators using forensic genetic approaches. Forensic Sci. Int. Genet. 2013, 7, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Suryawanshi, K.R.; Bhatnagar, Y.V.; Redpath, S.; Mishra, C. People, predators and perceptions: Patterns of livestock depredation by snow leopards and wolves. J. Appl. Ecol. 2013, 50, 550–560. [Google Scholar] [CrossRef]
- Rosas-Rosas, O.C.; Bender, L.C.; Valdez, R. Jaguar and puma predation on cattle calves in northeastern Sonora, Mexico. Rangel Ecol. Manag. 2008, 61, 554–560. [Google Scholar] [CrossRef]
- Palmeira, F.B.; Crawshaw, P.G., Jr.; Haddad, C.M.; Ferraz, K.M.P.; Verdade, L.M. Cattle depredation by puma (Puma concolor) and jaguar (Panthera onca) in central-western Brazil. Biol. Conserv. 2008, 141, 118–125. [Google Scholar] [CrossRef]
- Bergman, D.L.; Sparklin, W.; Carrillo, C.D.; Schmidt, J.A., III; Bender, S.; Breck, S.W. Depredation investigation: Using canine spread to identify the predator species. In Proceedings of the 24th Vertebrate Pest Conference, Sacramento, CA, USA, 22–25 February 2010; Timm, R.M., Fagerstone, K.A., Eds.; University of California: Davis, CA, USA, 2010; pp. 304–307. [Google Scholar] [CrossRef]
- Yravedra, J.; Maté-González, M.Á.; Courtenay, L.A.; González-Aguilera, D.; Fernández, M.F. The use of canid tooth marks on bone for the identification of livestock predation. Sci. Rep. 2019, 9, 16301. [Google Scholar] [CrossRef]
- Mumma, M.A.; Soulliere, C.E.; Mahoney, S.P.; Waits, L.P. Enhanced understanding of predator–prey relationships using molecular methods to identify predator species, individual and sex. Mol. Ecol. Resour. 2014, 14, 100–108. [Google Scholar] [CrossRef]
- Lino, S.; Lino, A.; Fernandes, J.M.; Ferreira, E.; Fonseca, C.; Aliácar, S.; Cadete, D.; Rosalino, L.M.; Carvalho, J. Environmental field conditions and sampling effort affect the molecular identification success of livestock predators. Mammal. Biol. 2023, 103, 339–345. [Google Scholar] [CrossRef]
- López-Bao, J.V.; Frank, J.; Svensson, L.; Åkesson, M.; Langefors, Å. Building public trust in compensation programs through accuracy assessments of damage verification protocols. Biol. Conserv. 2017, 213, 36–41. [Google Scholar] [CrossRef]
- Sugimoto, T.; Nagata, J.; Aramilev, V.V.; McCullough, D.R. Population size estimation of Amur tigers in Russian Far East using noninvasive genetic samples. J. Mammal. 2012, 93, 93–101. [Google Scholar] [CrossRef]
- Sugimoto, T.; Aramilev, V.V.; Kerley, L.L.; Nagata, J.; Miquelle, D.G.; McCullough, D.R. Noninvasive genetic analyses for estimating population size and genetic diversity of the remaining Far Eastern leopard (Panthera pardus orientalis) population. Conserv. Genet. 2014, 15, 521–532. [Google Scholar] [CrossRef]
- Harms, V.; Nowak, C.; Carl, S.; Muñoz-Fuentes, V. Experimental evaluation of genetic predator identification from saliva traces on wildlife kills. J. Mammal. 2015, 96, 138–143. [Google Scholar] [CrossRef]
- Ernest, H.B.; Boyce, W.M. DNA identification of mountain lions involved in livestock predation and public safety incidents and investigations. In Proceedings of the 19th Vertebrate Pest Conference, San Diego, CA, USA, 6–9 March 2000; Salmon, T.P., Crabb, A.C., Eds.; University of California: Davis, CA, USA, 2000; pp. 290–294. [Google Scholar] [CrossRef]
- Williams, C.L.; Blejwas, K.; Johnston, J.J.; Jaeger, M.M. A coyote in sheep’s clothing: Predator identification from saliva. Wildl. Soc. Bull. 2003, 31, 926–932. [Google Scholar]
- Blejwas, K.M.; Williams, C.L.; Shin, G.T.; McCullough, D.R.; Jaeger, M.M. Salivary DNA evidence convicts breeding male coyotes of killing sheep. J. Wildl. Manag. 2006, 70, 1087–1093. [Google Scholar] [CrossRef]
- Khorozyan, I.; Waltert, M. Variation and conservation implications of the effectiveness of anti-bear interventions. Sci. Rep. 2020, 10, 15341. [Google Scholar] [CrossRef]
- Guerisoli, M.D.L.M.; Luengos Vidal, E.; Caruso, N.; Giordano, A.J.; Lucherini, M. Puma–livestock conflicts in the Americas: A review of the evidence. Mammal Rev. 2021, 51, 228–246. [Google Scholar] [CrossRef]
- Palmeira, F.B.L.; Trinca, C.T.; Haddad, C.M. Livestock predation by puma (Puma concolor) in the highlands of a southeastern Brazilian Atlantic Forest. Environ. Manag. 2015, 56, 903–915. [Google Scholar] [CrossRef]
- Garrote, G.; Rodríguez-Castellanos, F.; Trujillo, F.; Mosquera-Guerra, F. Características de los ataques de jaguar (Panthera onca) sobre el ganado y evaluación económica de las pérdidas en fincas ganaderas de los Llanos Orientales (Vichada, Colombia). In Conflicto Entre Felinos y Humanos en América Latina; Lasso, C., Ed.; Serie editorial Fauna silvestre neotropical; Instituto de Investagación de Recursos Biológicos Alexander von Humboldt (IAvH): Bogotá, Colombia, 2017; pp. 89–102. [Google Scholar]
- Guerisoli, M.D.L.M.; Luengos Vidal, E.; Fanchini, M.; Caruso, N.; Casanave, E.B.; Lucherini, M. Characterization of puma–livestock conflicts in rangelands of central Argentina. R. Soc. Open Sci. 2017, 4, 170852. [Google Scholar] [CrossRef]
- Jędrzejewski, W.; Carreño, R.; Sánchez-Mercado, A.; Schmidt, K.; Abarca, M.; Robinson, H.S.; Boede, E.O.; Hoogesteijn, R.; Viloria, A.L.; Cerda, H.; et al. Human-jaguar conflicts and the relative importance of retaliatory killing and hunting for jaguar (Panthera onca) populations in Venezuela. Biol. Conserv. 2017, 209, 524–532. [Google Scholar] [CrossRef]
- Rodriguez, V.; Poo-Muñoz, D.A.; Escobar, L.E.; Astorga, F.; Medina-Vogel, G. Carnivore–livestock conflicts in Chile: Evidence and methods for mitigation. Hum. Wildl. Interact. 2019, 13, 50–62. [Google Scholar] [CrossRef]
- Gallardo, G.; Pacheco, L.F.; Rios, R.S.; Jiménez, J.E. Predation of livestock by puma (Puma concolor) and culpeo fox (Lycalopex culpaeus): Numeric and economic perspectives. Therya 2020, 11, 359–373. [Google Scholar] [CrossRef]
- Pinto, R.A.R.V.; Butrón, R.; Martel, C. Reports of feeding incidents of cattle by Andean bear (Tremarctos ornatus) in Central Peru. Rev. Mex. Mastozool. 2020, 10, 25–32. [Google Scholar] [CrossRef]
- Kleemann, J.; Koo, H.; Hensen, I.; Mendieta-Leiva, G.; Kahnt, B.; Kurze, C.; Inclan, D.J.; Cuenca, P.; Noh, J.K.; Horrmann, M.H.; et al. Priorities of action and research for the protection of biodiversity and ecosystem services in continental Ecuador. Biol. Conserv. 2022, 265, 109404. [Google Scholar] [CrossRef]
- Tirira, D.G. Lista Roja de los mamíferos del Ecuador. In Libro Rojo de los Mamíferos del Ecuador; Tirira, D.G., Ed.; Editorial Murciélago Blanco: Quito, Ecuador, 2021; pp. 3–82. [Google Scholar]
- Espinosa, S.; Álvarez-Solas, S.; Pacheco, J.; Salvador, J.; Utreras, V.; Nivelo-Villavicencio, C.; Zapata-Ríos, G.; Tirira, D.G. Jaguar (Panthera onca). In Libro Rojo de los Mamíferos del Ecuador; Tirira, D.G., Ed.; Editorial Murciélago Blanco: Quito, Ecuador, 2021; p. 60. [Google Scholar]
- Espinosa, S.; Salvador, J.; Álvarez-Solas, S.; Tirira, D.G. Puma (Puma concolor). In Libro Rojo de los Mamíferos del Ecuador; Tirira, D.G., Ed.; Editorial Murciélago Blanco: Quito, Ecuador, 2021; p. 60. [Google Scholar]
- Urgilés-Verdugo, C.A.; Cisneros-Vidal, R.; García, S.; Molina-Proaño, S.; Potaufeu, Y.; Sánchez-Karste, F.; Zapata-Ríos, G.; Tirira, D.G. Oso andino (Tremarctos ornatus). In Libro Rojo de los Mamíferos del Ecuador; Tirira, D.G., Ed.; Editorial Murciélago Blanco: Quito, Ecuador, 2021; p. 61. [Google Scholar]
- Zapata-Ríos, G.; Branch, L.C. Altered activity patterns and reduced abundance of native mammals in sites with feral dogs in the high Andes. Biol. Conserv. 2016, 193, 9–16. [Google Scholar] [CrossRef]
- Zapata-Ríos, G.; Branch, L.C. Mammalian carnivore occupancy is inversely related to presence of domestic dogs in the high Andes of Ecuador. PLoS ONE 2018, 13, e0192346. [Google Scholar] [CrossRef] [PubMed]
- Díaz, E.A.; Donoso, G.; Sáenz, C.; Dueñas, I.; Cabrera, F. Clinical and pathological findings in a Dwarf Red Brocket Mazama rufina (Mammalia: Cetartiodactyla: Cervidae) attacked by dogs. J. Threat. Taxa 2020, 12, 16885–16890. [Google Scholar] [CrossRef]
- Díaz, E.A.; Sáenz, C.; Segnini, G.; Villagómez, A.; Díaz, R.F.; Zug, R. Dystocia and cesarean section in a free-ranging ocelot (Leopardus pardalis) after traumatic spinal cord injury resulting from dog (Canis familiaris) attack. Open Vet. J. 2021, 11, 422–430. [Google Scholar] [CrossRef]
- Díaz, E.A.; Sáenz, C.; Vega, Y.; Rubio, E.; González, G.; Zug, R.; Zapata-Ríos, G. Dog and cat-related attacks on wildlife in the Metropolitan District of Quito, Ecuador: An integrative approach to reduce the impact. Ecosyst. People 2023, 19, 2191735. [Google Scholar] [CrossRef]
- Narváez, V.; Zapata-Ríos, G. Guía para la Identificación de Ataques a Animales Domésticos Causados por Carnívoros Grandes; Ministerio del Ambiente y Wildlife Conservation Society—Ecuador: Quito, Ecuador, 2016; 14p, Available online: https://global.wcs.org/Resources/Publications/Publications-Search-II/ctl/view/mid/13340/pubid/DMX3368600000.aspx (accessed on 29 November 2023).
- Goode, M.R.; Cheong, S.Y.; Li, N.; Ray, W.C.; Bartlett, C.W. Collection and extraction of saliva DNA for next generation sequencing. JoVE 2014, 90, e51697. [Google Scholar] [CrossRef]
- Shahzadi, I.; Janjua, S. A universal primer set to amplify the cytochrome c oxidase subunit I gene in bears. Ursus 2014, 25, 73–77. [Google Scholar] [CrossRef]
- Luo, S.J.; Kim, J.H.; Johnson, W.E.; van der Walt, J.; Martenson, J.; Yuhki, N.; Miquelle, D.G.; Uphyrkina, O.; Goodrich, J.M.; Quigley, H.B.; et al. Phylogeography and Genetic Ancestry of Tigers (Panthera tigris). PLoS Biol. 2004, 2, e442. [Google Scholar] [CrossRef]
- Lang, L.; Tessier, N.; Gauthier, M.; Wissink, R.; Jolicoeur, H.; Lapointe, F.J. Genetic confirmation of cougars (Puma concolor) in eastern Canada. Northeast Nat. 2013, 20, 383–396. [Google Scholar] [CrossRef]
- Abdel-Rahman, S.; El-Saadani, M.; Ashry, K.; Haggag, A. Detection of Adulteration and Identification of Cat’s, Dog’s, Donkey’s and Horse’s Meat Using Species-Specific PCR and PCR-RFLP Techniques. Aust. J. Basic Appl. Sci. 2009, 3, 1716–1719. [Google Scholar]
- Hossain, M.A.M.; Ali, M.E.; Hamid, S.B.A.; Mustafa, S.; Desa, M.N.M.; Zaidul, I.S.M. Targeting double genes in multiplex PCR for discriminating bovine, buffalo and porcine materials in food chain. Food Control 2017, 73, 175–184. [Google Scholar] [CrossRef]
- Goldstein, I.; Paisley, S.; Wallace, R.; Jorgenson, J.P.; Cuesta, F.; Castellanos, A. Andean bear–livestock conflicts: A review. Ursus 2006, 17, 8–15. [Google Scholar] [CrossRef]
- Álvarez, H.G.; Zapata-Ríos, G. Do social factors influence perceptions of the jaguar Panthera onca in Ecuador? Oryx 2022, 56, 308–315. [Google Scholar] [CrossRef]
- Ganz, T.R.; DeVivo, M.T.; Reese, E.M.; Prugh, L.R. Wildlife whodunnit: Forensic identification of predators to inform wildlife management and conservation. Wildl. Soc. Bull. 2023, 47, e1386. [Google Scholar] [CrossRef]
- Chinchilla, S.; Berghe, E.V.D.; Polisar, J.; Arévalo, C.; Bonacic, C. Livestock–Carnivore Coexistence: Moving beyond Preventive Killing. Animals 2022, 12, 479. [Google Scholar] [CrossRef]
- Rojas-VeraPinto, R.; Bautista, C.; Selva, N. Living high and at risk: Predicting Andean bear occurrence and conflicts with humans in southeastern Peru. Glob. Ecol. Conserv. 2022, 36, e02112. [Google Scholar] [CrossRef]
- Kleemann, J.; Zamora, C.; Villacis-Chiluisa, A.B.; Cuenca, P.; Koo, H.; Noh, J.K.; Fürst, C.; Thiel, M. Deforestation in Continental Ecuador with a Focus on Protected Areas. Land 2022, 11, 268. [Google Scholar] [CrossRef]
- Zukowski, B.; Ormsby, A. Andean bear livestock depredation and community perceptions in northern Ecuador. Hum. Dimens. Wildl. 2016, 21, 111–126. [Google Scholar] [CrossRef]
- Treves, A.; Krofel, M.; McManus, J. Predator control should not be a shot in the dark. Front. Ecol. Environ. 2016, 14, 380–388. [Google Scholar] [CrossRef]
- Moreira-Arce, D.; Ugarte, C.S.; Zorondo-Rodríguez, F.; Simonetti, J.A. Management tools to reduce carnivore-livestock conflicts: Current gap and future challenges. Rangel Ecol. Manag. 2018, 71, 389–394. [Google Scholar] [CrossRef]
- Spencer, K.; Sambrook, M.; Bremner-Harrison, S.; Cilliers, D.; Yarnell, R.W.; Brummer, R.; Whitehouse-Tedd, K. Livestock guarding dogs enable human-carnivore coexistence: First evidence of equivalent carnivore occupancy on guarded and unguarded farms. Biol. Conserv. 2020, 241, 108256. [Google Scholar] [CrossRef]
- Urbigkit, C.D. Livestock guardian dogs and cattle protection: Opportunities, challenges, and methods. Hum. Wildl. Interact. 2019, 13, 9. [Google Scholar] [CrossRef]
- Smith, B.R.; Yarnell, R.W.; Uzal, A.; Whitehouse-Tedd, K. The ecological effects of livestock guarding dogs (LGDs) on target and non-target wildlife. J. Vertebr. Biol. 2020, 69, 20103-1. [Google Scholar] [CrossRef]
- Whitehouse-Tedd, K.; Wilkes, R.; Stannard, C.; Wettlaufer, D.; Cilliers, D. Reported livestock guarding dog-wildlife interactions: Implications for conservation and animal welfare. Biol. Conserv. 2020, 241, 108249. [Google Scholar] [CrossRef]
- Ellwanger, J.H.; Chies, J.A.B. The triad “dogs, conservation and zoonotic diseases”—An old and still neglected problem in Brazil. Perspect. Ecol. Conserv. 2019, 17, 157–161. [Google Scholar] [CrossRef]
- Ortiz-Prado, E.; Ponce-Zea, J.; Ramirez, D.; Stewart-Ibarra, A.M.; Armijos, L.; Yockteng, J.; Cárdenas, W.B. Rabies epidemiology and control in Ecuador. Glob. J. Health Sci. 2016, 8, 113. [Google Scholar] [CrossRef]
- Capistros, F.B.; Roman, J.R.; Andrade, P.G.; Aluisa, E.M.; Ortiz, W.B.; Berkvens, D.; Saegerman, C. Dogs as potential mechanical vector of transmission of Brucella spp., in dairy cattle in the sierra of Ecuador. In Joint Colloquium on Zoonoses and Neglected Infectious Diseases of Africa; Berkvens, D., Saegerman, C., Eds.; IMT/DVTD: Johannesburg, South Africa, 2011; p. 41. [Google Scholar]
- Calvopina, M.; Cabezas-Moreno, M.; Cisneros-Vásquez, E.; Paredes-Betancourt, I.; Bastidas-Caldes, C. Diversity and prevalence of gastrointestinal helminths of free-roaming dogs on coastal beaches in Ecuador: Potential for zoonotic transmission. Vet. Parasitol. Reg. Stud. Rep. 2023, 40, 100859. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.R. Mapping attack hotspots to mitigate human–carnivore conflict: Approaches and applications of spatial predation risk modeling. Biodivers. Conserv. 2015, 24, 2887–2911. [Google Scholar] [CrossRef]
- Mestanza-Ramón, C.; Henkanaththegedara, S.M.; Vásconez, P.; Vargas, Y.; Sánchez, M.; Constante, D.; Jimenez, M.; Charco, M.; Mestanza, P. In-Situ and ex-situ biodiversity conservation in Ecuador: A review of policies, actions and challenges. Diversity 2020, 12, 315. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | DNA Region | Reference Literature | Standardized Protocol |
|---|---|---|---|
| Andean bear (Tremarctos ornatus) | COI | [54] | A 25 μL reaction system containing 1X PCR buffer, 1.5 mM MgCl2, 0.2 mM dNTPs, 0.2 mM of each primer, and 1.5 U Platinum® Taq DNA polymerase. The amplification protocol was an initial denaturation step at 94 °C for 5 min, followed by 35 cycles of 94 °C denaturation for 45 s, 58 °C annealing for 1 min, 72 °C extension for 1 min, with a final extension step at 72 °C for 10 min. |
| Jaguar (Panthera onca) | 12S | [55] | A 15 μL reaction system containing 1X PCR buffer, 1 mM MgCl2, 0.2 mM dNTPs, 0.5 μM of each primer, and 1U of Platinum® Taq DNA polymerase. The amplification protocol was an initial denaturation step at 95 °C for 10 min, followed by 35 cycles of 95 °C denaturation for 30 s, 54 °C annealing for 30 s, 72 °C extension for 45 s, with a final extension step at 72 °C for 10 min. |
| Puma (Puma concolor) | 16S | [56] | A 15 μL reaction system containing 1X PCR buffer, 2 mM MgCl2, 0.2 mM dNTPs, 0.5 μM of each primer, and 1U of Platinum® Taq DNA polymerase. The amplification protocol was an initial denaturation step at 95 °C for 3 min, followed by 40 cycles of 94 °C denaturation for 30 s, 53 °C annealing for 45 s, 72 °C extension for 1 min, with a final extension step at 72 °C for 5 min. |
| Domestic dog (Canis familiaris) | CytB | [57] | A 15 μL reaction system containing 1X PCR buffer, 1.5 mM MgCl2, 0.2 mM dNTPs, 0.5 μM of each primer, and 1.5 U of Platinum® Taq DNA polymerase. The amplification protocol was an initial denaturation step at 95 °C for 5 min, followed by 35 cycles of 94 °C denaturation for 30 s, 60 °C annealing for 30 s, 72 °C extension for 30 s, with a final extension step at 72 °C for 10 min. |
| Species Identification | ||||
|---|---|---|---|---|
| Blind Assay Number | Andean Bear | Jaguar | Puma | Domestic Dog |
| 1 | X | X | X | |
| 2 | X | X | ||
| 3 | X | X | X | X |
| 4 | X | |||
| 5 | X | X | ||
| Attack # | Site of the Attack | Approximate Date of Attack | Date of Sample Collection | Animal and Part of the Body from Which Sample Was Taken | Detected Animal Using Species-Specific DNA Markers |
|---|---|---|---|---|---|
| 1 | Nono, Pichincha | No information | 27 December 2021 | Cow—neck | Domestic dog, puma |
| Cow—inferior extremity | None | ||||
| 2 | Yanahurco grande, Cotopaxi | 20 February 2022 (20:00) | 21 February 2022 | Sheep 1—unknown | Domestic dog |
| Sheep 2—unknown | Domestic dog | ||||
| Sheep 3—unknown | Domestic dog | ||||
| Sheep 4—unknown | Domestic dog | ||||
| 3 | Paluguillo, Pichincha | On or before 25 December 2022 | 27 December 2022 | Sheep-Face | None |
| 4 | Nono bajo, Pichincha | On or before 23 April 2023 | 26 April 2023 | Cow—face | None |
| Cow—thorax | None | ||||
| Cow—leg | None | ||||
| 5 | Chilla, El Oro | 22 December 2023 | 22 December 2023 | Sheep—neck | Domestic dog |
| 6 | Las Villegas, Santo Domingo | 14 January 2024 | 14 January 2024 | Mule—mouth, nose, head, hip (individual sample origin was not identified) | Domestic dog |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz, E.A.; Pozo, M.J.; Alarcón, P.; Pozo, G.; Zug, R.; Sáenz, C.; Torres, M.d.L. A Genetic Tool to Identify Predators Responsible for Livestock Attacks in South America and Recommendations for Human–Wildlife Conflict Mitigation. Animals 2024, 14, 838. https://doi.org/10.3390/ani14060838
Díaz EA, Pozo MJ, Alarcón P, Pozo G, Zug R, Sáenz C, Torres MdL. A Genetic Tool to Identify Predators Responsible for Livestock Attacks in South America and Recommendations for Human–Wildlife Conflict Mitigation. Animals. 2024; 14(6):838. https://doi.org/10.3390/ani14060838
Chicago/Turabian StyleDíaz, Eduardo A., María José Pozo, Pablo Alarcón, Gabriela Pozo, Rebecca Zug, Carolina Sáenz, and Maria de Lourdes Torres. 2024. "A Genetic Tool to Identify Predators Responsible for Livestock Attacks in South America and Recommendations for Human–Wildlife Conflict Mitigation" Animals 14, no. 6: 838. https://doi.org/10.3390/ani14060838
APA StyleDíaz, E. A., Pozo, M. J., Alarcón, P., Pozo, G., Zug, R., Sáenz, C., & Torres, M. d. L. (2024). A Genetic Tool to Identify Predators Responsible for Livestock Attacks in South America and Recommendations for Human–Wildlife Conflict Mitigation. Animals, 14(6), 838. https://doi.org/10.3390/ani14060838