
Molecular Characterization and Phylogenetic Analysis of Flightin Gene in Vespa basalis (Hymenoptera, Vespidae)



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Reagents
2.3. Instruments
2.4. RNA Extraction and cDNA Synthesis
2.5. PCR Amplification of the Flightin Gene
2.6. Online ExPASy Tool
3. Results
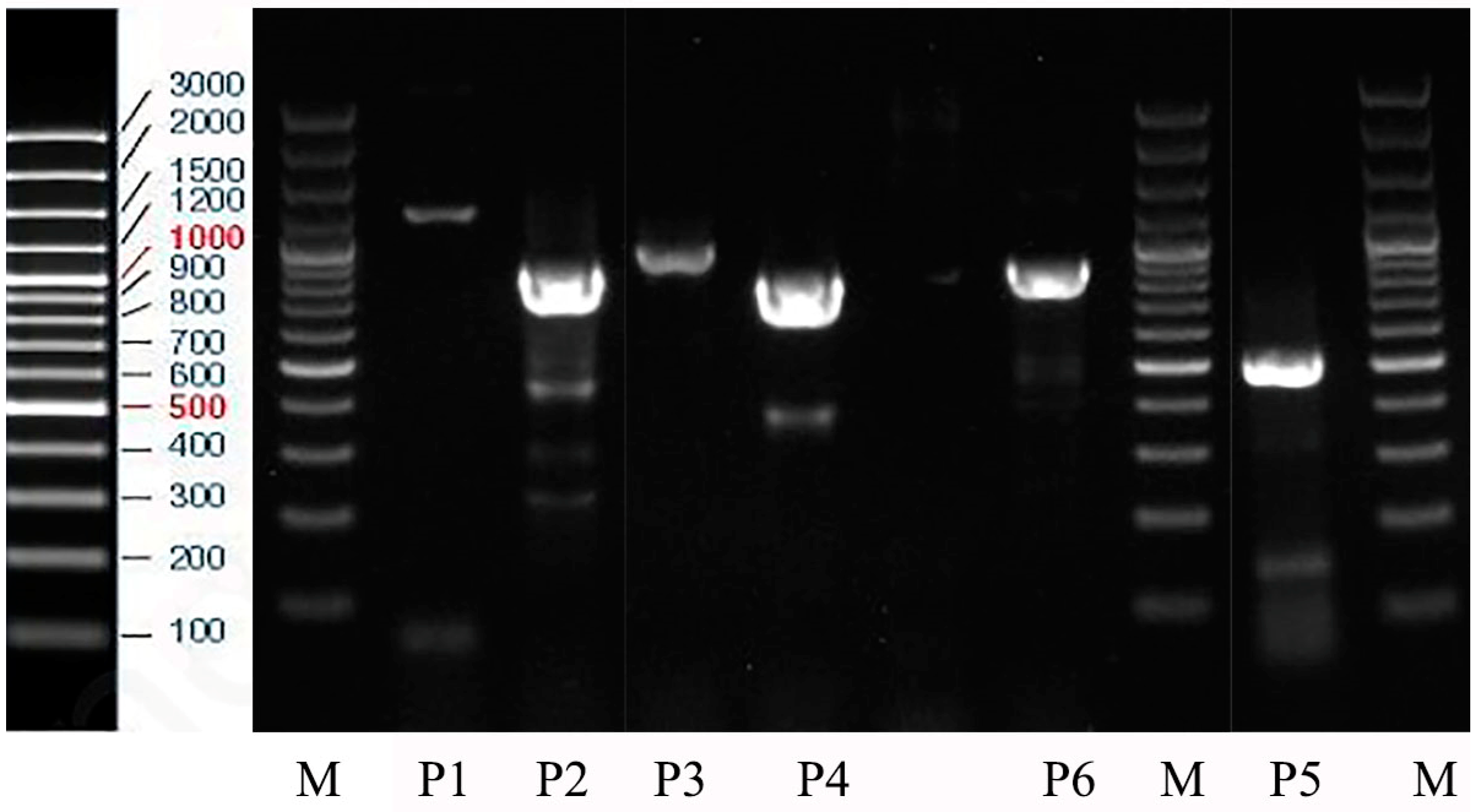
3.1. PCR Result
3.2. Molecular Weight Isoelectric Point Findings
3.3. Conservative Domains and Functional Sites
3.4. Prediction of Transmembrane Topology
3.5. Prediction of Secondary Structures
3.6. 3D Structure Prediction
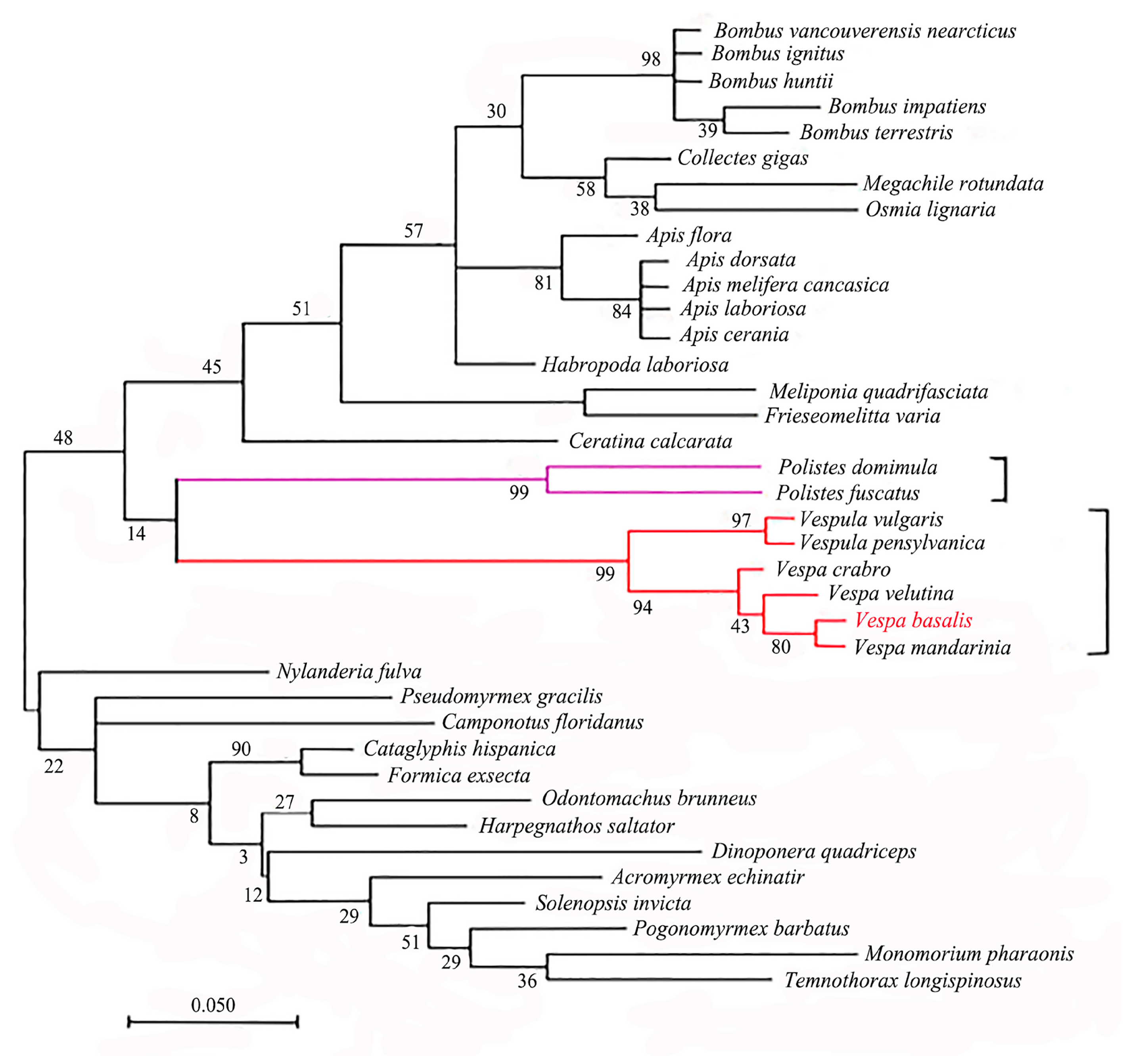
3.7. Protein Homology Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Martin, S.J. The Old World Versus New World Social Wasps: Similarities, Differences, and Threats. In Neotropical Social Wasps: Basic and Applied Aspects; Springer: Berlin/Heidelberg, Germany, 2021; pp. 317–326. [Google Scholar]
- Schmitz, S.; Schankin, C.J.; Prinz, H.; Curwen, R.S.; Ashton, P.D.; Caves, L.S.; Veigel, C. Molecular evolutionary convergence of the flight muscle protein arthritis in Diptera and Hemiptera. Mol. Biol. Evol. 2003, 20, 2019–2033. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.L.; Van Achterberg, C.; Chen, X.X. Potentially Lethal Social Wasps, Fauna of the Chinese Vespinae (Hymenoptera: Vespidae); Science Press: Beijing, China, 2015. [Google Scholar]
- Tan, J.L.; Xing, L.X. “Dengxian Shide Hufeng Mian. (Let Everyone Know Wasps Which Can Attack Humans)” Illustrated Introduction and Prevention of Hornets; Shaanxi Science and Technology Press: Xi’an, China, 2022. [Google Scholar]
- Josephson, R.K. Comparative physiology of insect flight muscle. In Nature’s Versatile Engine: Insect Flight Muscle Inside and Out; Springer: Berlin/Heidelberg, Germany, 2006; pp. 34–43. [Google Scholar]
- Tennekes, H. The Simple Science of Flight, Revised and Expanded Edition: From Insects to Jumbo Jets; MIT Press: Cambridge, MA, USA, 2009. [Google Scholar]
- Abbasi Yeganeh, F.; Rastegarpouyani, H.; Li, J.; Taylor, K.A. Structure of the Drosophila melanogaster Flight Muscle Myosin Filament at 4.7 Å Resolution Reveals New Details of Non-Myosin Proteins. Int. J. Mol. Sci. 2023, 24, 14936. [Google Scholar] [CrossRef] [PubMed]
- Bullard, B.; Pastore, A. Regulating the contraction of insect flight muscle. J. Muscle Res. Cell Motil. 2011, 32, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Holmes, K.C. Steric blocking mechanism explains stretch activation in insect flight muscle. Proc. Natl. Acad. Sci. USA 2011, 108, 7–8. [Google Scholar] [CrossRef]
- Ayme-Southgate, A.J.; Southgate, R.J.; Philipp, R.A.; Sotka, E.E.; Kramp, C. The myofibrillar protein, projectin, is highly conserved across insect evolution except for its PEVK domain. J. Mol. Evol. 2008, 67, 653–669. [Google Scholar] [CrossRef]
- Ghazi, A.; Anant, S.; Vijay Raghavan, K. Apterous mediates the development of direct flight muscles autonomously and indirect flight muscles through epidermal cues. Development 2000, 127, 5309–5318. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, M.Q.; Wang, X.Q.; Guo, J.S.; Li, D.T.; Xue, J.; Zhang, C.X. The flightin gene is necessary for the emission of vibrational signals in the rice brown planthopper (Nilaparvata lugens Stǻl). J. Insect Physiol. 2019, 112, 101–108. [Google Scholar] [CrossRef]
- Vigoreaux, J.O.; Saide, J.D.; Valgeirsdottir, K.; Pardue, M.L. Flightin, a novel myofibrillar protein of Drosophila stretch-activated muscles. J. Cell Biol. 1993, 121, 587–598. [Google Scholar] [CrossRef]
- Contompasis, J.L.; Nyland, L.R.; Maughan, D.W.; Vigoreaux, J.O. Flightin is necessary for length determination, structural integrity, and large bending stiffness of insect flight muscle thick filaments. J. Mol. Biol. 2010, 359, 340–348. [Google Scholar] [CrossRef]
- Iwamoto, H. Structure, function and evolution of insect flight muscle. Biophysics 2011, 7, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Reedy, M.C.; Bullard, B.; Vigoreaux, J.O. Flightin is essential for thick filament assembly and sarcomere stability in Drosophila flight muscles. J. Cell Biol. 2000, 151, 1483–1500. [Google Scholar] [CrossRef] [PubMed]
- Schöck, F.; González-Morales, N. The insect perspective on Z-disc structure and biology. J. Cell Sci. 2022, 135, jcs260179. [Google Scholar] [CrossRef] [PubMed]
- Qiu, F.; Brendel, S.; Cunha, P.M.; Astola, N.; Song, B.; Furlong, E.E.; Bullard, B. Myofilin, a protein in the thick filaments of insect muscle. J. Cell Sci. 2005, 118, 1527–1536. [Google Scholar] [CrossRef]
- Ayme-Southgate, A.; Feldman, S.; Fulmer, D. Myofilament proteins in the synchronous flight muscles of Manduca sexta show both similarities and differences to Drosophila melanogaster. Insect Biochem. Mol. Biol. 2015, 62, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Henkin, J.A.; Maughan, D.W.; Vigoreaux, J. Mutations that affect flightin expression in Drosophila alter the viscoelastic properties of flight muscle fibers. Am. J. Physiol.-Cell Physiol. 2004, 286, C65–C72. [Google Scholar] [CrossRef] [PubMed]
- Ayer, G.; Vigoreaux, J.O. Flightin is a myosin rod binding protein. Cell Biochem. Biophys. 2003, 38, 41–54. [Google Scholar] [CrossRef]
- Auguin, D.; Moussaoui, D.; Robblee, J.P.; Kikuti, C.; Sirkia, M.E.; Canon, L.; Houdusse, A.M. Small molecules modulating force production: A new perspective against myosin-associated diseases. Biophys. J. 2023, 122, 260a. [Google Scholar] [CrossRef]
- Lemas, D.; Lekkas, P.; Ballif, B.A.; Vigoreaux, J.O. Intrinsic disorder and multiple phosphorylations constrain the evolution of the flightin N-terminal region. J. Proteom. 2016, 135, 191–200. [Google Scholar] [CrossRef]
- Marden, J.H. Variability in the size, composition, and function of insect flight muscles. Annu. Rev. Physiol. 2000, 62, 157–178. [Google Scholar] [CrossRef]
- Vigoreaux, J.O. Alterations in flightin phosphorylation in Drosophila flight muscles are associated with myofibrillar defects engendered by actin and myosin heavy-chain mutant alleles. Biochem. Genet. 1994, 32, 301–314. [Google Scholar] [CrossRef]
- Sun, W.; Margam, V.M.; Sun, L.; Buczkowski, G.; Bennett, G.W.; Schermerhorn, B.; Pittendrigh, B.R. Genome-wide analysis of phenobarbital-inducible genes in Drosophila melanogaster. Insect Mol. Biol. 2006, 15, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Yue, G.H.; Orban, L.A. Simple and affordable method for high-throughput DNA extraction from animal tissues for polymerase chain reaction. Electrophoresis 2005, 26, 3081–3083. [Google Scholar] [CrossRef] [PubMed]
- Li, J. Cryo-EM structure of the flight muscle thick filament from the bumble bee, Bombus ignitus, with novel non-myosin densities revealed. Biophys. J. 2023, 122, 260a. [Google Scholar] [CrossRef]
- Menard, L.M.; Wood, N.B.; Vigoreaux, J.O. Contiguity and Structural Impacts of a Non-Myosin Protein within the Thick Filament Myosin Layer. Biology 2021, 10, 613. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Zhang, X.Q.; Xu, H.J. Molecular characterization of the flightin gene in the wing-dimorphic planthopper Nilaparvata lugens, and its evolution in Pancrustacea. Insect Biochem. Mol. Biol. 2013, 43, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Xu, B.; Huang, Y. The complete mitochondrial genome of a medically important wasp, Vespa magnifica (Hymenoptera, Vespidae). Mitochondrial DNA B Resour. 2022, 7, 93–95. [Google Scholar] [CrossRef]
- Fu, Y.; Wu, T.; Yu, H.; Xu, J.; Zhang, J.Z.; Fu, D.Y.; Ye, H. The Transcription of Flight Energy Metabolism Enzymes Declined with Aging While Enzyme Activity Increased in the Long-Distance Migratory Moth, Spodoptera frugiperda. Insects 2022, 13, 936. [Google Scholar] [CrossRef]
- Wang, H.; Wen, Q.; Wang, T.; Ran, F.; Wang, M.; Fan, X.; Li, Z.; Fan, X.; Tan, J. Next-Generation Sequencing of Four Mitochondrial Genomes of Dolichovespula (Hymenoptera: Vespidae) with a Phylogenetic Analysis and Divergence Time Estimation of Vespidae. Animals 2022, 12, 3004. [Google Scholar] [CrossRef]
- Hahn, C.; Bachmann, L.; Chevreux, B. Reconstructing mitochondrial genomes directly from genomic next-generation sequencing reads—A baiting and iterative mapping approach. Nucleic Acids Res. 2013, 41, e129. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Functional Site | Flightin | |
|---|---|---|
| Site | Sequence Signature | |
| Casein kinase II phosphorylation site | 14–17 | TapE |
| N-myristoylation site | 24–29 | GAkeGG |
| 28–33 | GGaeGA | |
| 29–34 | GAegAA | |
| 32–37 | GAapGE | |
| Protein kinase C phosphorylation site | 102–104 | TmR |
| 114–116 | StK | |
| 115–117 | TkR | |
| 130–132 | TpR | |
| 143–145 | SlK | |
| Tyrosine kinase phosphorylation site 2 | 116–123 | KrsaDmkY |
| 4 different patterns found | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ullah, H.; Huang, X.; Zhou, T.; Tang, Y.; Zhu, D.; Xu, H.; Tan, J. Molecular Characterization and Phylogenetic Analysis of Flightin Gene in Vespa basalis (Hymenoptera, Vespidae). Animals 2024, 14, 978. https://doi.org/10.3390/ani14060978
Ullah H, Huang X, Zhou T, Tang Y, Zhu D, Xu H, Tan J. Molecular Characterization and Phylogenetic Analysis of Flightin Gene in Vespa basalis (Hymenoptera, Vespidae). Animals. 2024; 14(6):978. https://doi.org/10.3390/ani14060978
Chicago/Turabian StyleUllah, Hasin, Xiaojuan Huang, Tong Zhou, Yan Tang, Danyang Zhu, Hongli Xu, and Jiangli Tan. 2024. "Molecular Characterization and Phylogenetic Analysis of Flightin Gene in Vespa basalis (Hymenoptera, Vespidae)" Animals 14, no. 6: 978. https://doi.org/10.3390/ani14060978
APA StyleUllah, H., Huang, X., Zhou, T., Tang, Y., Zhu, D., Xu, H., & Tan, J. (2024). Molecular Characterization and Phylogenetic Analysis of Flightin Gene in Vespa basalis (Hymenoptera, Vespidae). Animals, 14(6), 978. https://doi.org/10.3390/ani14060978