
Identification, Expression and Evolutional Analysis of Two cyp19-like Genes in Amphioxus

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. RNA Extraction and cDNA Synthesis
2.3. Cloning of the cyp19
2.4. Bioinformatics Analysis
2.5. Real-Time PCR
2.6. Paraffin Section In Situ Hybridization (ISH)
2.7. Hematoxylin and Eosin Staining
2.8. Protocol for Generating Mouse Polyclonal Antibodies
2.9. Western Blot
2.10. Immunohistochemical Staining
2.11. Subcelluar Localization
2.12. Statistical Analysis
3. Results
3.1. Cloning of the cyp19
3.2. Bioinformatic Analysis of the cyp19
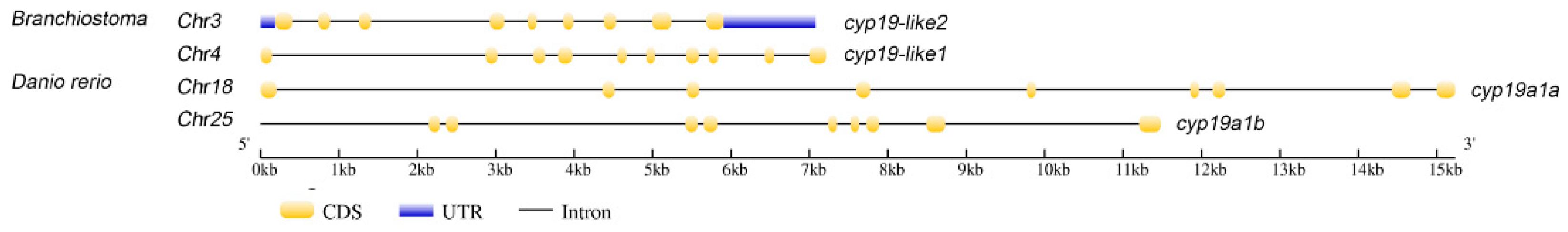
3.2.1. Comparison of cyp19 Structures between Zebrafish and Amphioxus
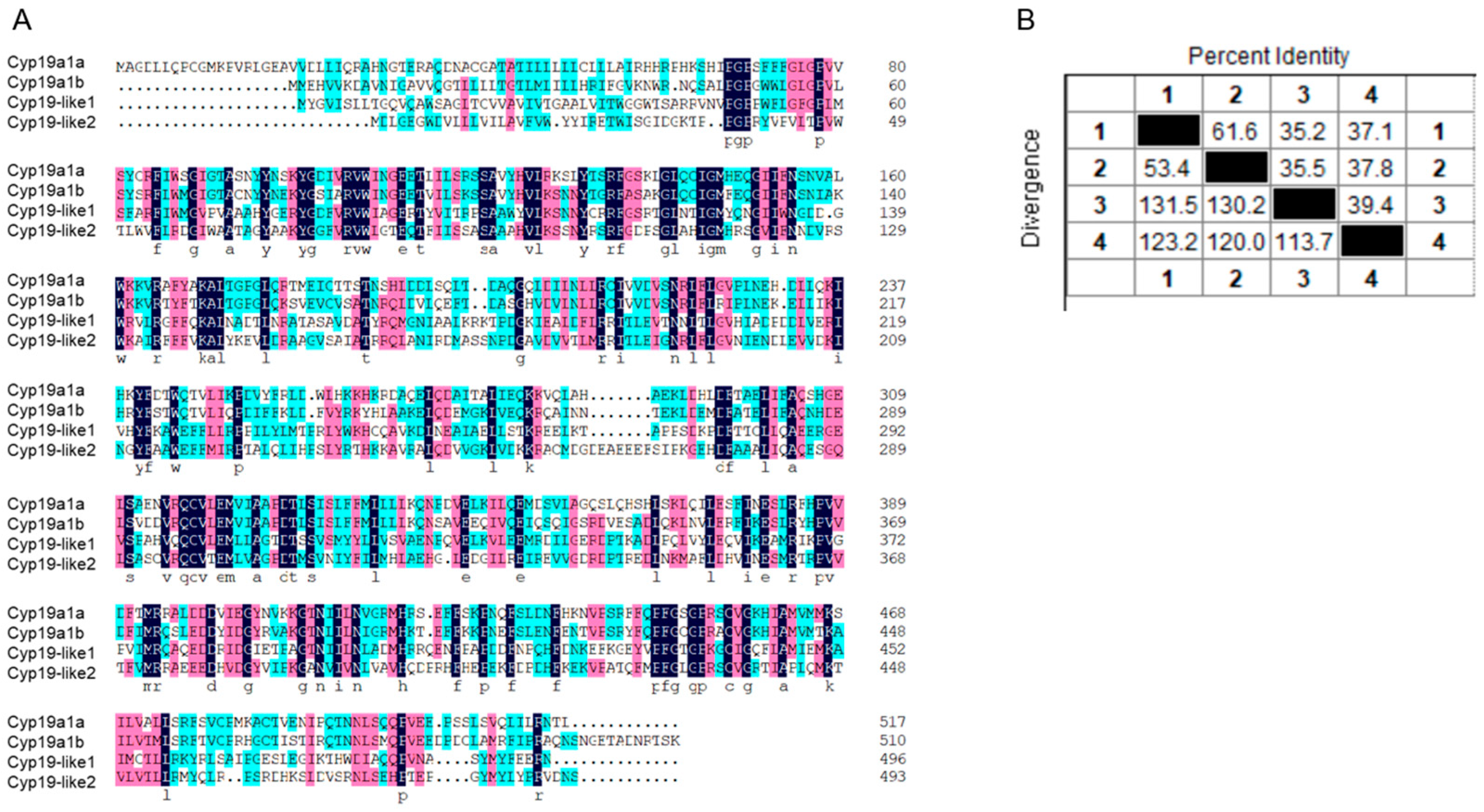
3.2.2. Comparison of Sequence Similarity between Zebrafish and Amphioxus Cyp19
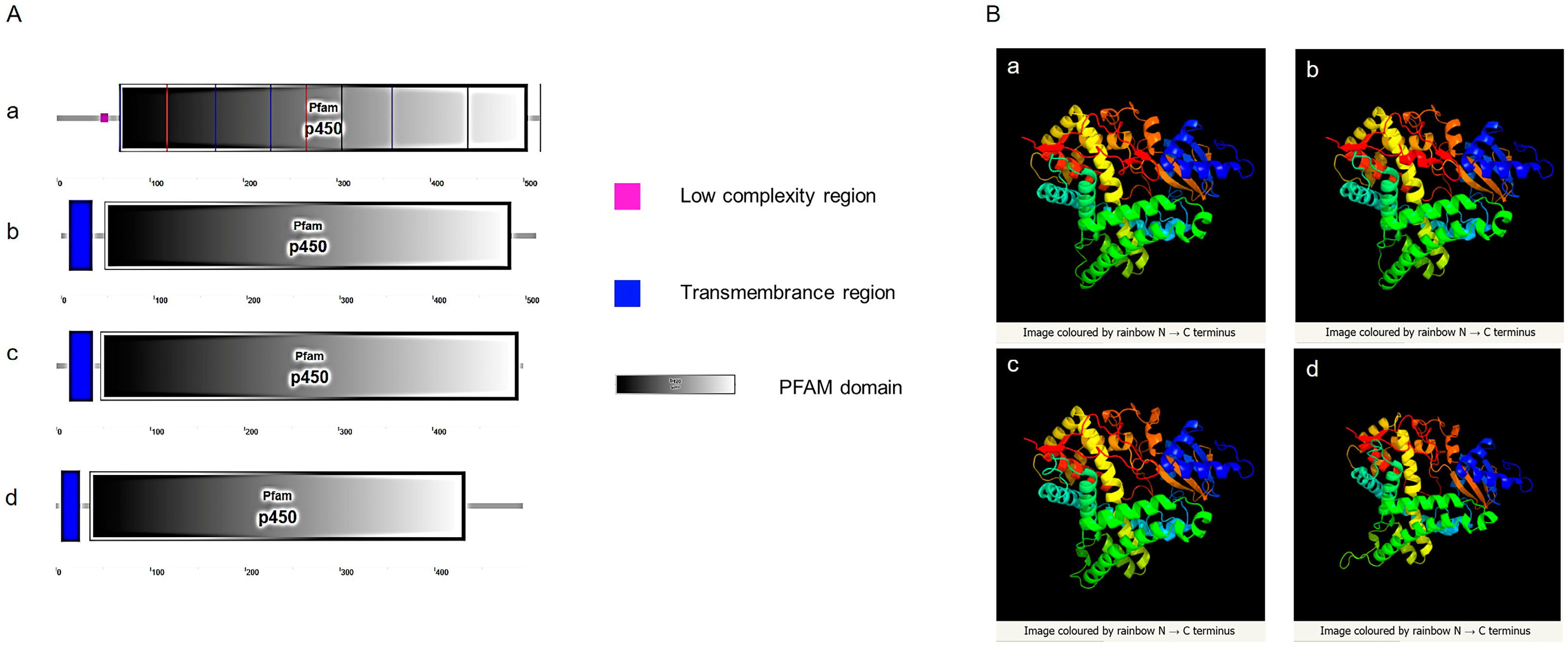
3.2.3. Prediction and Comparison of Cyp19 Structures between Zebrafish and Amphioxus
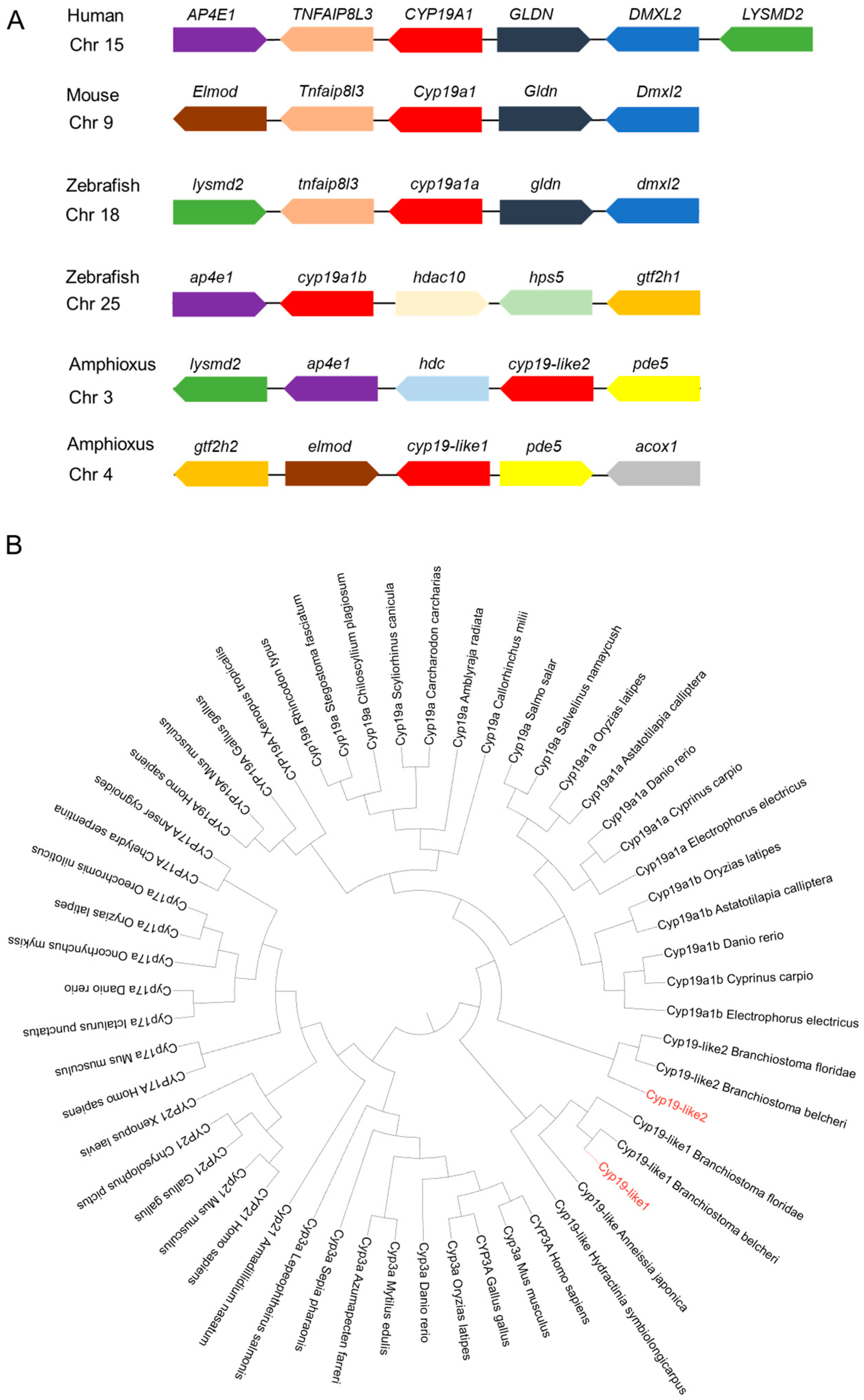
3.2.4. Synteny and Evolutionary Analysis of Cyp19
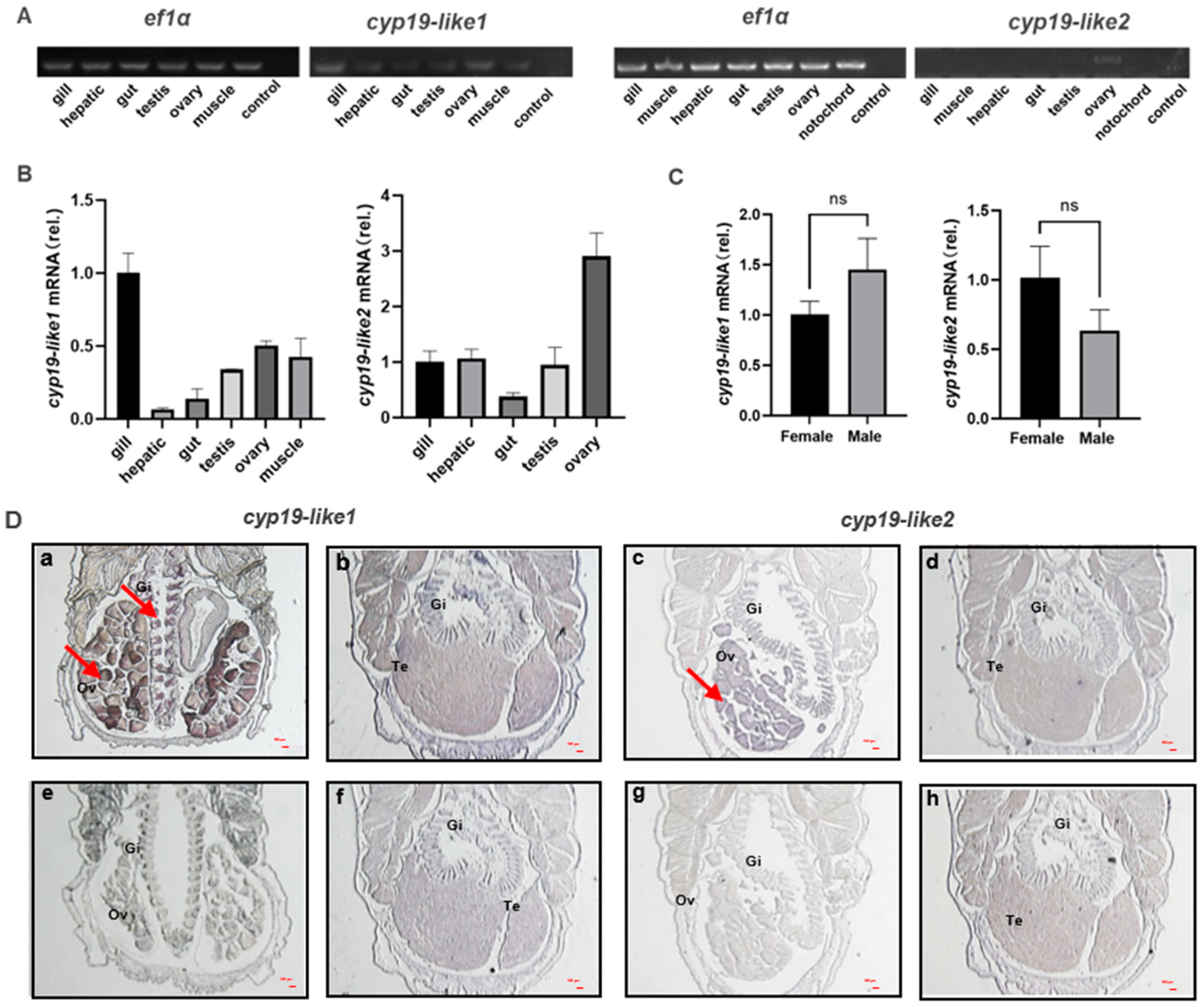
3.3. Expression Level of Two cyp19-like Genes in the Gonads of Male and Female Amphioxus
3.3.1. Differential Tissue Expression of cyp19
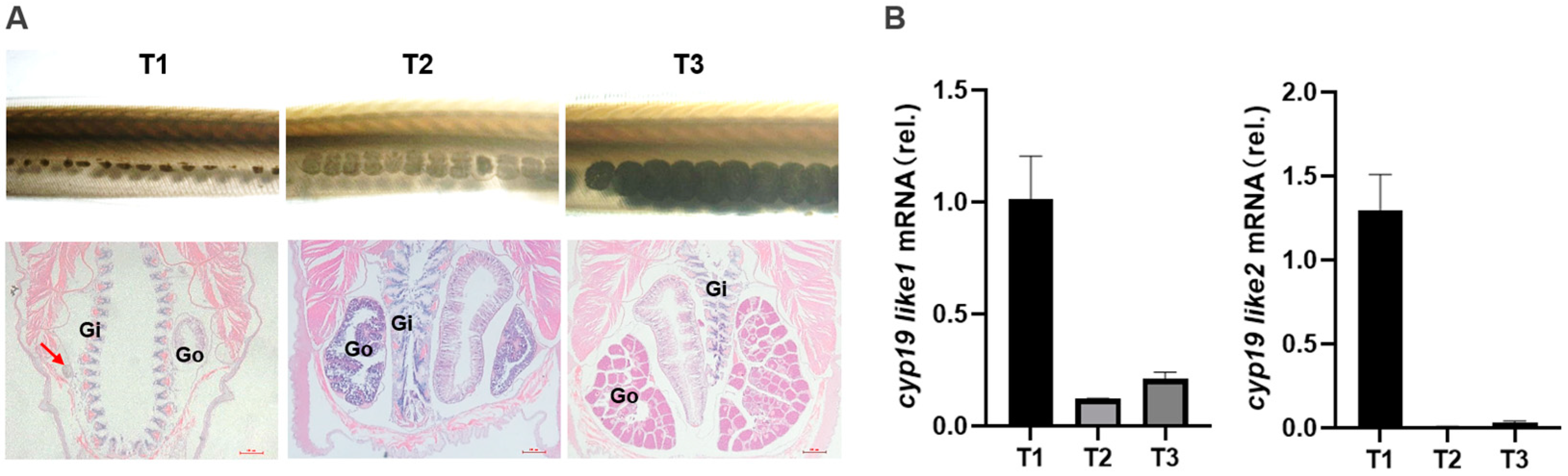
3.3.2. Expression of cyp19 at Different Stages
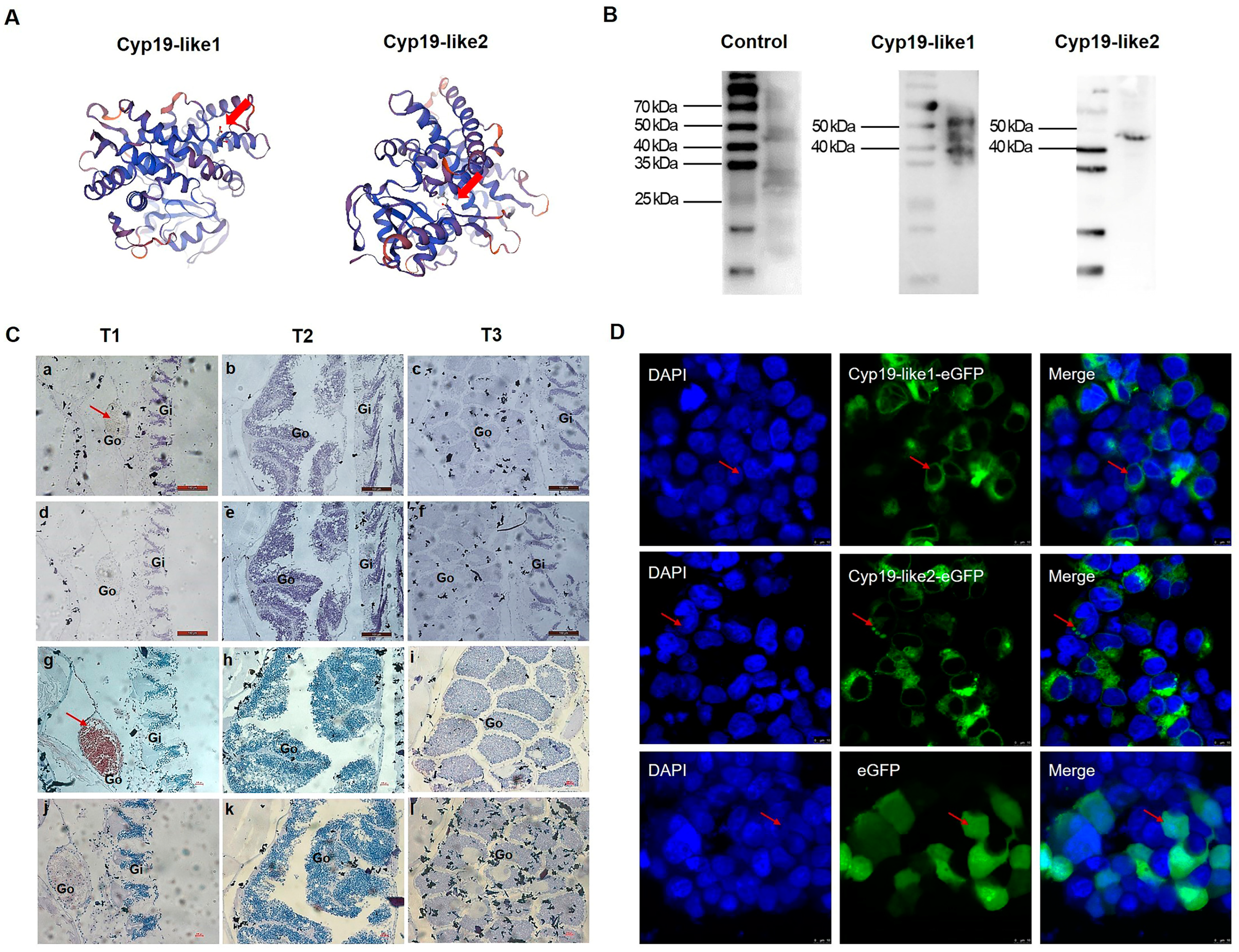
3.3.3. Temporal Expression and Cellular Localization of Cyp19
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Koopman, P.; Gubbay, J.; Vivian, N.; Goodfellow, P.; Lovell-Badge, R. Male development of chromosomally female mice transgenic for Sry. Nature 1991, 351, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Wright, E.; Hargrave, M.R.; Christiansen, J.; Cooper, L.; Kun, J.; Evans, T.; Gangadharan, U.; Greenfield, A.; Koopman, P. The Sry-related gene Sox9 is expressed during chondrogenesis in mouse embryos. Nat. Genet. 1995, 9, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Swain, A.; Narvaez, V.; Burgoyne, P.; Camerino, G.; Lovell-Badge, R. Dax1 antagonizes Sry action in mammalian sex determination. Nature 1998, 391, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, M.; Shinomiya, A.; Kinoshita, M.; Suzuki, A.; Kobayashi, T.; Paul-Prasanth, B.; Lau, E.L.; Hamaguchi, S.; Sakaizumi, M.; Nagahama, Y. DMY gene induces male development in genetically female (XX) medaka fish. Proc. Natl. Acad. Sci. USA 2007, 104, 3865–3870. [Google Scholar] [CrossRef] [PubMed]
- Marín, I.; Baker, B.S. The evolutionary dynamics of sex determination. Science 1998, 281, 1990–1994. [Google Scholar] [CrossRef] [PubMed]
- Raymond, C.S.; Shamu, C.E.; Shen, M.M.; Seifert, K.J.; Hirsch, B.; Hodgkin, J.; Zarkower, D. Evidence for evolutionary conservation of sex-determining genes. Nature 1998, 391, 691–695. [Google Scholar] [CrossRef] [PubMed]
- Ottolenghi, C.; Pelosi, E.; Tran, J.; Colombino, M.; Douglass, E.; Nedorezov, T.; Cao, A.; Forabosco, A.; Schlessinger, D. Loss of Wnt4 and Foxl2 leads to female-to-male sex reversal extending to germ cells. Hum. Mol. Genet. 2007, 16, 2795–2804. [Google Scholar] [CrossRef] [PubMed]
- Dranow, D.B.; Hu, K.; Bird, A.M. Bmp15 Is an Oocyte-Produced Signal Required for Maintenance of the Adult Female Sexual Phenotype in Zebrafish. PLoS Genet. 2016, 12, e1006323. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, S.I.; Nakaki, F. ZGLP1 is a determinant for the oogenic fate in mice. Science 2020, 367, eaaw4115. [Google Scholar] [CrossRef]
- Sreenivasan, R.; Jiang, J.; Wang, X.; Bártfai, R.; Kwan, H.Y.; Christoffels, A.; Orbán, L. Gonad differentiation in zebrafish is regulated by the canonical Wnt signaling pathway. Biol. Reprod. 2014, 90, 45. [Google Scholar] [CrossRef]
- Picard, M.A.L.; Vicoso, B.; Bertrand, S.; Escriva, H. Diversity of Modes of Reproduction and Sex Determination Systems in Invertebrates, and the Putative Contribution of Genetic Conflict. Genes 2021, 12, 1136. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Kobayashi, K.; Watanabe, H.; Iguchi, T. Environmental sex determination in the branchiopod crustacean Daphnia magna: Deep conservation of a Doublesex gene in the sex-determining pathway. PLoS Genet. 2011, 7, e1001345. [Google Scholar] [CrossRef] [PubMed]
- Holland, L.Z.; Laudet, V.; Schubert, M. The chordate amphioxus: An emerging model organism for developmental biology. Cell. Mol. Life Sci. CMLS 2004, 61, 2290–2308. [Google Scholar] [CrossRef] [PubMed]
- Bourlat, S.J.; Juliusdottir, T.; Lowe, C.J.; Freeman, R.; Aronowicz, J.; Kirschner, M.; Lander, E.S.; Thorndyke, M.; Nakano, H.; Kohn, A.B.; et al. Deuterostome phylogeny reveals monophyletic chordates and the new phylum Xenoturbellida. Nature 2006, 444, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Delsuc, F.; Brinkmann, H.; Chourrout, D.; Philippe, H. Tunicates and not cephalochordates are the closest living relatives of vertebrates. Nature 2006, 439, 965–968. [Google Scholar] [CrossRef] [PubMed]
- Putnam, N.H.; Butts, T.; Ferrier, D.E.; Furlong, R.F.; Hellsten, U.; Kawashima, T.; Robinson-Rechavi, M.; Shoguchi, E.; Terry, A.; Yu, J.K.; et al. The amphioxus genome and the evolution of the chordate karyotype. Nature 2008, 453, 1064–1071. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, S.; Escriva, H. Evolutionary crossroads in developmental biology: Amphioxus. Development 2011, 138, 4819–4830. [Google Scholar] [CrossRef] [PubMed]
- Holland, P. The dawn of amphioxus molecular biology—A personal perspective. Int. J. Dev. Biol. 2017, 61, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Henmi, Y. Biology of the amphioxus, Branchiostoma belcheri in the Ariake Sea, Japan II. Reproduction. Zool. Sci. 2003, 20, 907–918. [Google Scholar] [CrossRef]
- Kubokawa, K.; Mizuta, T.; Morisawa, M.; Azuma, N. Gonadal state of wild amphioxus populations and spawning success in captive conditions during the breeding period in Japan. Zool. Sci. 2003, 20, 889–895. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, S.; Chu, J. G-banding patterns of the chromosomes of amphioxus Branchiostoma belcheri tsingtauense. Hereditas 2004, 141, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Guraya, S.S. Recent advances in the morphology, histochemistry, and biochemistry of steroid-synthesizing cellular sites in the nonmammalian vertebrate ovary. Int. Rev. Cytol. 1976, 44, 365–409. [Google Scholar] [CrossRef] [PubMed]
- Mizuta, T.; Kubokawa, K. Presence of Sex Steroids and Cytochrome P450 Genes in Amphioxus. Endocrinology 2007, 148, 3554–3565. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.E. Co-evolution of steroidogenic and steroid-inactivating enzymes and adrenal and sex steroid receptors. Mol. Cell. Endocrinol. 2004, 215, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Kubokawa, K.; Tando, Y.; Roy, S. Evolution of the reproductive endocrine system in chordates. Integr. Comp. Biol. 2010, 50, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhang, L.; Li, H.; Wang, Y.; Zhang, S.; Liu, Z. Characterization of GRP as a functional neuropeptide in basal chordate amphioxus. Int. J. Biol. Macromol. 2020, 142, 384–394. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, M.; Zhang, L.; Zhong, S.; Jiang, W.; Wang, Z.; Sun, C.; Zhang, S.; Liu, Z. Functional characterization of an orexin neuropeptide in amphioxus reveals an ancient origin of orexin/orexin receptor system in chordate. Sci. China. Life Sci. 2019, 62, 1655–1669. [Google Scholar] [CrossRef]
- Wang, P.; Wang, M.; Ji, G.; Yang, S.; Zhang, S.; Liu, Z. Demonstration of a Functional Kisspeptin/Kisspeptin Receptor System in Amphioxus With Implications for Origin of Neuroendocrine Regulation. Endocrinology 2017, 158, 1461–1473. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Wu, X. A ZZ/ZW Sex Chromosome System in Cephalochordate Amphioxus. Genetics 2020, 214, 617–622. [Google Scholar] [CrossRef]
- Huang, Z.; Xu, L.; Cai, C. Three amphioxus reference genomes reveal gene and chromosome evolution of chordates. Proc. Natl. Acad. Sci. USA 2023, 120, e2201504120. [Google Scholar] [CrossRef]
- Manikandan, P.; Nagini, S. Cytochrome P450 Structure, Function and Clinical Significance: A Review. Curr. Drug Targets 2018, 19, 38–54. [Google Scholar] [CrossRef] [PubMed]
- Simpson, E.R.; Mahendroo, M.S.; Means, G.D.; Kilgore, M.W.; Hinshelwood, M.M.; Graham-Lorence, S.; Amarneh, B.; Ito, Y.; Fisher, C.R.; Michael, M.D.; et al. Aromatase cytochrome P450, the enzyme responsible for estrogen biosynthesis. Endocr. Rev. 1994, 15, 342–355. [Google Scholar] [CrossRef] [PubMed]
- Crews, D.; Bergeron, J.M. Role of reductase and aromatase in sex determination in the red-eared slider (Trachemys scripta), a turtle with temperature-dependent sex determination. J. Endocrinol. 1994, 143, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Sakata, N.; Tamori, Y.; Wakahara, M. P450 aromatase expression in the temperature-sensitive sexual differentiation of salamander (Hynobius retardatus) gonads. Int. J. Dev. Biol. 2005, 49, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Kitano, T.; Takamune, K.; Nagahama, Y.; Abe, S.I. Aromatase inhibitor and 17alpha-methyltestosterone cause sex-reversal from genetical females to phenotypic males and suppression of P450 aromatase gene expression in Japanese flounder (Paralichthys olivaceus). Mol. Reprod. Dev. 2000, 56, 1–5. [Google Scholar] [CrossRef]
- Elbrecht, A.; Smith, R.G. Aromatase enzyme activity and sex determination in chickens. Science 1992, 255, 467–470. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Zou, Y.; Jiao, S.; Tan, X.; Wu, Z.; Liang, D.; Zhang, P.; You, F. Significant association of cyp19a promoter methylation with environmental factors and gonadal differentiation in olive flounder Paralichthys olivaceus. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2017, 208, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.W.; Zhang, Z.; Qin, M.; Ge, W. Knockout of Zebrafish Ovarian Aromatase Gene (cyp19a1a) by TALEN and CRISPR/Cas9 Leads to All-male Offspring Due to Failed Ovarian Differentiation. Sci. Rep. 2016, 6, 37357. [Google Scholar] [CrossRef] [PubMed]
- Nakamoto, M.; Shibata, Y.; Ohno, K.; Usami, T.; Kamei, Y.; Taniguchi, Y.; Todo, T.; Sakamoto, T.; Young, G.; Swanson, P.; et al. Ovarian aromatase loss-of-function mutant medaka undergo ovary degeneration and partial female-to-male sex reversal after puberty. Mol. Cell. Endocrinol. 2018, 460, 104–122. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, S.; Liu, Z.; Zhang, L.; Zhang, W. Epigenetic modifications during sex change repress gonadotropin stimulation of cyp19a1a in a teleost ricefield eel (Monopterus albus). Endocrinology 2013, 154, 2881–2890. [Google Scholar] [CrossRef]
- Yin, Y.; Tang, H.; Liu, Y.; Chen, Y.; Li, G.; Liu, X.; Lin, H. Targeted Disruption of Aromatase Reveals Dual Functions of cyp19a1a During Sex Differentiation in Zebrafish. Endocrinology 2017, 158, 3030–3041. [Google Scholar] [CrossRef] [PubMed]
- Böhne, A.; Heule, C.; Boileau, N.; Salzburger, W. Expression and sequence evolution of aromatase cyp19a1 and other sexual development genes in East African cichlid fishes. Mol. Biol. Evol. 2013, 30, 2268–2285. [Google Scholar] [CrossRef] [PubMed]
- Ramallo, M.R.; Morandini, L.; Birba, A.; Somoza, G.M.; Pandolfi, M. From molecule to behavior: Brain aromatase (cyp19a1b) characterization, expression analysis and its relation with social status and male agonistic behavior in a Neotropical cichlid fish. Horm. Behav. 2017, 89, 176–188. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, S.; Lu, H.; Zhang, L.; Zhang, W. Genes encoding aromatases in teleosts: Evolution and expression regulation. Gen. Comp. Endocrinol. 2014, 205, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Castro, L.F.; Santos, M.M.; Reis-Henriques, M.A. The genomic environment around the Aromatase gene: Evolutionary insights. BMC Evol. Biol. 2005, 5, 43. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, S. EF1α is a useful internal reference for studies of gene expression regulation in amphioxus Branchiostoma japonicum. Fish Shellfish. Immunol. 2012, 32, 1068–1073. [Google Scholar] [CrossRef] [PubMed]
- Tchoudakova, A.; Kishida, M.; Wood, E.; Callard, G.V. Promoter characteristics of two cyp19 genes differentially expressed in the brain and ovary of teleost fish. J. Steroid Biochem. Mol. Biol. 2001, 78, 427–439. [Google Scholar] [CrossRef] [PubMed]
- Kishida, M.; McLellan, M.; Miranda, J.A.; Callard, G.V. Estrogen and xenoestrogens upregulate the brain aromatase isoform (P450aromB) and perturb markers of early development in zebrafish (Danio rerio). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2001, 129, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Mak, P.; Tchoudakova, A.; Callard, G.; Chen, S. Different catalytic properties and inhibitor responses of the goldfish brain and ovary aromatase isozymes. Gen. Comp. Endocrinol. 2001, 123, 180–191. [Google Scholar] [CrossRef]
- Tong, S.K.; Chiang, E.F.; Hsiao, P.H.; Chung, B. Phylogeny, expression and enzyme activity of zebrafish cyp19 (P450 aromatase) genes. J. Steroid Biochem. Mol. Biol. 2001, 79, 299–303. [Google Scholar] [CrossRef]
- Kishida, M.; Callard, G.V. Distinct cytochrome P450 aromatase isoforms in zebrafish (Danio rerio) brain and ovary are differentially programmed and estrogen regulated during early development. Endocrinology 2001, 142, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Gelinas, D.; Pitoc, G.A.; Callard, G.V. Isolation of a goldfish brain cytochrome P450 aromatase cDNA: mRNA expression during the seasonal cycle and after steroid treatment. Mol. Cell. Endocrinol. 1998, 138, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Tchoudakova, A.; Callard, G.V. Identification of multiple CYP19 genes encoding different cytochrome P450 aromatase isozymes in brain and ovary. Endocrinology 1998, 139, 2179–2189. [Google Scholar] [CrossRef] [PubMed]
- Conley, A.J.; Corbin, C.J.; Hughes, A.L. Adaptive evolution of mammalian aromatases: Lessons from Suiformes. J. Exp. Zool. Part A Ecol. Genet. Physiol. 2009, 311, 346–357. [Google Scholar] [CrossRef] [PubMed]
- Corbin, C.J.; Hughes, A.L.; Heffelfinger, J.R.; Berger, T.; Waltzek, T.B.; Roser, J.F.; Santos, T.C.; Miglino, M.A.; Oliveira, M.F.; Braga, F.C.; et al. Evolution of suiform aromatases: Ancestral duplication with conservation of tissue-specific expression in the collared peccary (Pecari tayassu). J. Mol. Evol. 2007, 65, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Amarneh, B.; Corbin, C.J.; Peterson, J.A.; Simpson, E.R.; Graham-Lorence, S. Functional domains of human aromatase cytochrome P450 characterized by linear alignment and site-directed mutagenesis. Mol. Endocrinol. 2007, 65, 1617. [Google Scholar]
- Shimozawa, O.; Sakaguchi, M.; Ogawa, H.; Harada, N.; Mihara, K.; Omura, T. Core glycosylation of cytochrome P-450(arom). Evidence for localization of N terminus of microsomal cytochrome P-450 in the lumen. J. Biol. Chem. 1993, 268, 21399–21402. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Description | Sequences (5′–3′) |
|---|---|---|
| PCR | P1(cyp19-like1 F) | ATGTACGGAGTGATCTCTCTCCTTA |
| P2(cyp19-like1 R) | CTAGTTTCTCTCTTCAAAGTACATG | |
| P3(cyp19-like2 F) | ATGGACCTAGGCGAAGGCTGGGACG | |
| P4(cyp19-like2 R) | TCAGCTGTTGTCCACCCTTGGGTAC | |
| Real-time PCR | P5(ef1α F) | TGCTGATTGTGGCTGCTGGTACTG |
| P6(ef1α R) | GGTGTAGGCCAGCAGGGCGTG | |
| P7(cyp19-like1 F) | GCTCAGGAGGACGACAGGATTG | |
| P8(cyp19-like1 R) | GCAGCAGCGTACACATGATGG | |
| P9(cyp19-like2 F) | TTCGCCGCTGCTCTCATCCA | |
| P10(cyp19-like2 R) | CGGTCTCCGACGACTTCTCTGA | |
| In situ hybridization | P11(cyp19-like1 F) | GCGTGGTCGCCGTTGTCGTT |
| P12(cyp19-like1 R) | CGCCGCAAGAAATCCAGAGCT | |
| P13(cyp19-like2 F) | GTGTATCCGCCATTGCTACC | |
| P14(cyp19-like2 R) | TCTCCGACGACTTCTCTGATT | |
| Polyclonal antibody | Cyp19-like1 | N′-CREELKTAPPSDKPD-C′ |
| preparation (mouse) | Cyp19-like2 | N′-CPSRDHKSLDVSRNL-C′ |
| Subcellular Localization | P15(cyp19-like1 F) | gcacagtggcggccgctcgagATGTACGGAGTGATCTCTCTCCTTACC |
| P16(cyp19-like1 R) | gctcaccattctagactcgagGTTTCTCTCTTCAAAGTACATGTAACTAGC | |
| P17(cyp19-like2 F) | gcacagtggcggccgctcgagATGGACCTAGGCGAAGGCTG | |
| P18(cyp19-like2 R) | gctcaccattctagactcgagGCTGTTGTCCACCCTTGGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Lin, J.; Li, W.; Ji, G.; Liu, Z. Identification, Expression and Evolutional Analysis of Two cyp19-like Genes in Amphioxus. Animals 2024, 14, 1140. https://doi.org/10.3390/ani14081140
Wang Y, Lin J, Li W, Ji G, Liu Z. Identification, Expression and Evolutional Analysis of Two cyp19-like Genes in Amphioxus. Animals. 2024; 14(8):1140. https://doi.org/10.3390/ani14081140
Chicago/Turabian StyleWang, Yajun, Jingyuan Lin, Wenjin Li, Guangdong Ji, and Zhenhui Liu. 2024. "Identification, Expression and Evolutional Analysis of Two cyp19-like Genes in Amphioxus" Animals 14, no. 8: 1140. https://doi.org/10.3390/ani14081140