
Health and Welfare Benefits of Computerized Cognitive Enrichment in California Sea Lions (Zalophus californianus) at the US Navy Marine Mammal Program

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
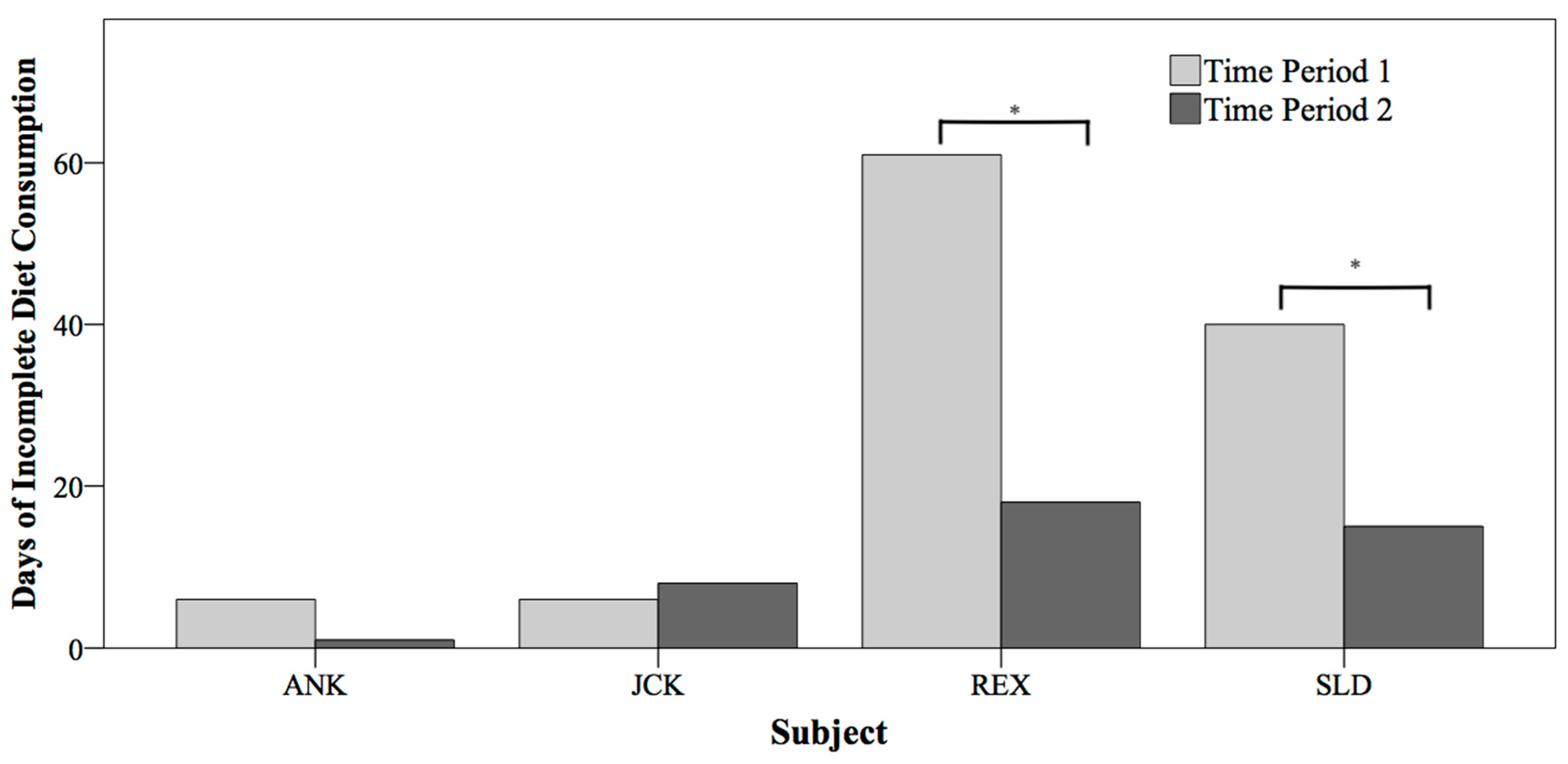
3.1. Incomplete Diet Consumption Day Analyses
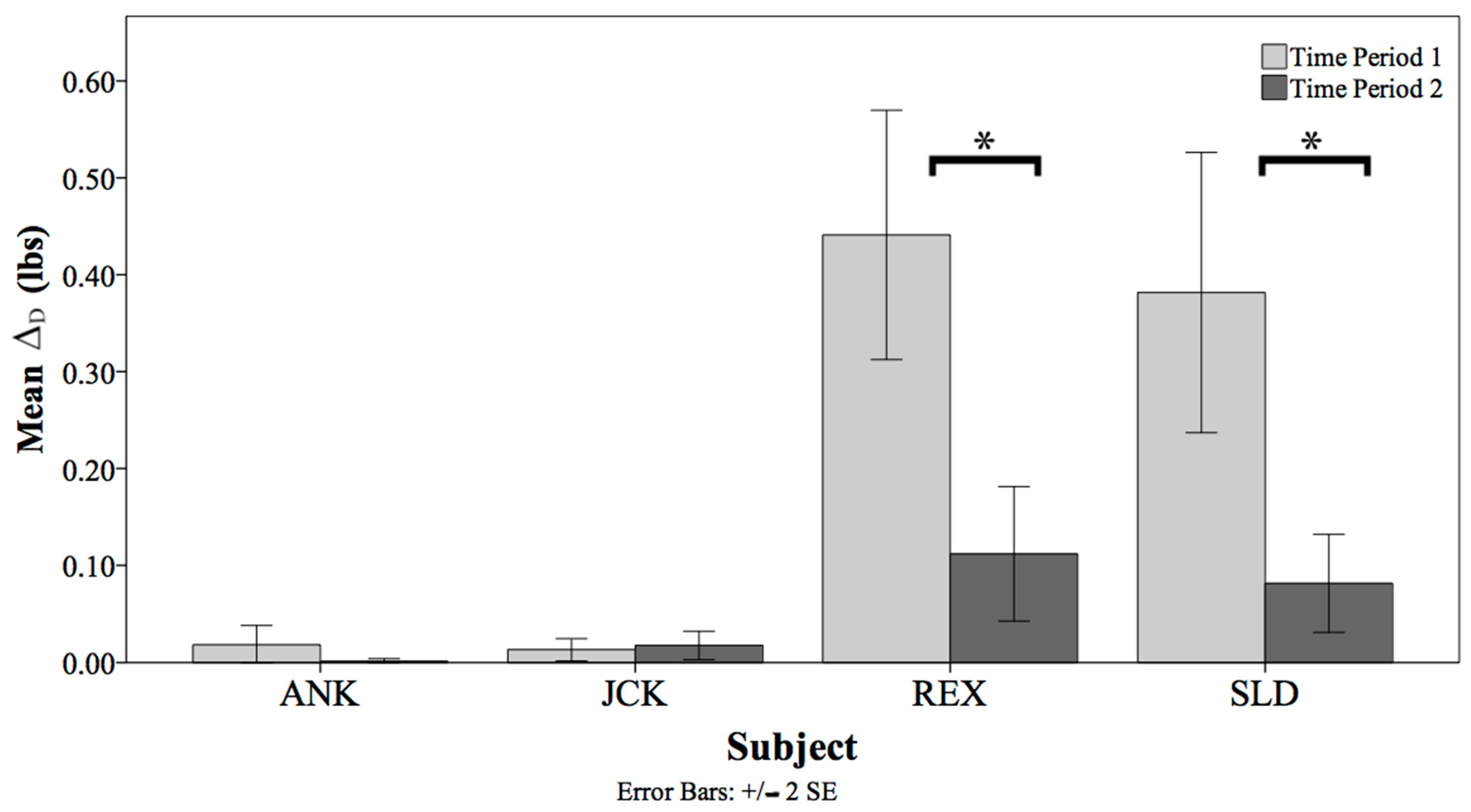
3.2. Diet Consumed Analyses
3.3. Number of Clinically Ill Days
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mellor, D.J.; Beausoleil, N.J.; Littlewood, K.E.; McLean, A.N.; McGreevy, P.D.; Jones, B.; Wilkins, C. The 2020 Five Domains Model: Including Human–Animal Interactions in Assessments of Animal Welfare. Animals 2020, 10, 1870. [Google Scholar] [CrossRef]
- Dawkins, M.S. A user’s guide to animal welfare science. Trends Ecol. Evol. 2006, 21, 77–82. [Google Scholar] [CrossRef]
- Miller, L.J. Visitor reaction to pacing behavior: Influence on the perception of animal care and interest in supporting zoological institutions. Zoo Biol. 2012, 31, 242–248. [Google Scholar] [CrossRef]
- Melfi, V. Is training zoo animals enriching? Appl. Anim. Behav. Sci. 2013, 147, 299–305. [Google Scholar] [CrossRef]
- Hacker, C.E.; Miller, L.J. Zoo visitor perceptions, attitudes, and conservation intent after viewing African elephants at the San Diego Zoo Safari Park. Zoo Biol. 2016, 35, 355–361. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, M.F.C.; Souto, A.; Caselli, B.; Sciel, N. People’s perception on animal welfare-why does it matter. Ethnobiol. Conserv. 2017, 6, 1–7. [Google Scholar] [CrossRef]
- Miller, L.J.; Luebke, J.F.; Matiasek, J. Viewing African and Asian elephants at accredited zoological institutions: Conservation intent and perceptions of animal welfare. Zoo Biol. 2018, 37, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Warsaw, D.a.S.J. The influence of animal welfare accreditation programmes on zoo visitor perceptions of the welfare of zoo animals. J. Zoo Aquar. Res. 2020, 8, 188–193. [Google Scholar]
- Veasey, J.S. Differing animal welfare conceptions and what they mean for the future of zoos and aquariums, insights from an animal welfare audit. Zoo Biol. 2022, 41, 292–307. [Google Scholar] [CrossRef]
- Ellis, T.; Yildiz, H.Y.; López-Olmeda, J.; Spedicato, M.T.; Tort, L.; Øverli, Ø.; Martins, C.I. Cortisol and finfish welfare. Fish Physiol. Biochem. 2012, 38, 163–188. [Google Scholar] [CrossRef]
- Tilbrook, A.J.; Ralph, C.R. Hormones, stress and the welfare of animals. Anim. Prod. Sci. 2018, 58, 408–415. [Google Scholar] [CrossRef]
- Heimbürge, S.; Kanitz, E.; Otten, W. The use of hair cortisol for the assessment of stress in animals. Gen. Comp. Endocrinol. 2019, 270, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Palme, R. Monitoring stress hormone metabolites as a useful, non-invasive tool for welfare assessment in farm animals. Anim. Welf. 2023, 21, 331–337. [Google Scholar] [CrossRef]
- Corwin, A.L. Training fish and aquatic invertebrates for husbandry and medical behaviors. Veter Clin. N. Am. Exot. Anim. Pr. 2012, 15, 455–467. [Google Scholar] [CrossRef] [PubMed]
- Forthman, D.L.; Ogden, J.J. The role of applied behavior analysis in zoo management-today and tomorrow. J. Appl. Behav. Anal. 1992, 25, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Laule, G.; Desmond, T. Positive reinforcement training as an enrichment strategy. In Second Nature: Environmental Enrichment for Captive Animals; M.J.D. Shepherdson, D.J., Hutchins, M., Eds.; Smithsonian Institution Press: London, UK, 1998; pp. 302–313. [Google Scholar]
- Lukas, K.E.; Marr, M.J.; Maple, T.L. Teaching Operant Conditioning at the Zoo. Teach. Psychol. 2016, 25, 112–116. [Google Scholar] [CrossRef]
- Fernandez, E.J.; Martin, A.L. Animal Training, Environmental Enrichment, and Animal Welfare: A History of Behavior Analysis in Zoos. J. Zool. Bot. Gard. 2021, 2, 531–543. [Google Scholar] [CrossRef]
- Fernandez, E.J. Training as enrichment: A critical review. Anim. Welf. 2023, 31, 1–12. [Google Scholar] [CrossRef]
- Ward, S.J.; Melfi, V. The implications of husbandry training on zoo animal response rates. Appl. Anim. Behav. Sci. 2013, 147, 179–185. [Google Scholar] [CrossRef]
- Mattison, S. Training birds and small mammals for medical behaviors. Veter Clin. N. Am. Exot. Anim. Pr. 2012, 15, 487–499. [Google Scholar] [CrossRef]
- Brando, S.I.C.A. Advances in Husbandry Training in Marine Mammal Care Programs. Int. J. Comp. Psychol. 2010, 23, 777–791. [Google Scholar] [CrossRef]
- Kuczaj, S.A., 2nd; Xitco, M.J., Jr. It Takes More Than Fish: The Psychology of Marine Mammal Training. Int. J. Comp. Psychol. 2002, 15, 186–200. [Google Scholar] [CrossRef]
- Lauderdale, L.K.; Walsh, M.T.; Mellen, J.D.; Granger, D.A.; Miller, L.J. Environmental enrichment, training, and habitat characteristics of common bottlenose dolphins (Tursiops truncatus) and Indo-Pacific bottlenose dolphins (Tursiops aduncus). PLoS ONE 2021, 16, e0253688. [Google Scholar] [CrossRef] [PubMed]
- Bassett, L.; Buchanan-Smith, H.M.; McKinley, J.; Smith, T.E. Effects of training on stress-related behavior of the common marmoset (Callithrix jacchus) in relation to coping with routine husbandry procedures. J. Appl. Anim. Welf Sci. 2003, 6, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Carroll, S.L.; Sykes, B.W.; Mills, P.C. Moving toward Fear-Free Husbandry and Veterinary Care for Horses. Animals 2022, 12, 2907. [Google Scholar] [CrossRef] [PubMed]
- Colahan, H.; Breder, C. Primate training at Disney’s Animal Kingdom. J. Appl. Anim. Welf. Sci. 2003, 6, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.; King, C.E. Husbandry training, using positive reinforcement techniques, for Marabou stork Leptoptilos crumeniferus at Edinburgh Zoo. Int. Zoo Yearb. 2013, 47, 171–180. [Google Scholar] [CrossRef]
- Lima, D.S.; Vergara-Parente, J.E.; Young, R.J.; Paszkiewicz, E. Training of Antillean manatee Trichechus manatus manatus Linnaeus, 1758 as a management technique for individual welfare. Lat. Am. J. Aquat. Mamm. 2005, 4, 61–68. [Google Scholar] [CrossRef]
- Martelli, P.; Krishnasamy, K. The Role of Preventative Medicine Programs in Animal Welfare and Wellbeing in Zoological Institutions. Animals 2023, 13, 2299. [Google Scholar] [CrossRef]
- Clegg, I.; Borger-Turner, J.; Eskelinen, H. C-Well: The development of a welfare assessment index for captive bottlenose dolphins (Tursiops truncatus). Anim. Welf. 2015, 24, 267–282. [Google Scholar] [CrossRef]
- Lauderdale, L.K.; Mellen, J.D.; Walsh, M.T.; Granger, D.A.; Miller, L.J. Towards understanding the welfare of cetaceans in accredited zoos and aquariums. PLoS ONE 2021, 16, e0255506. [Google Scholar] [CrossRef] [PubMed]
- Lauderdale, L.K.; Shorter, K.A.; Zhang, D.; Gabaldon, J.; Mellen, J.D.; Walsh, M.T.; Granger, D.A.; Miller, L.J. Bottlenose dolphin habitat and management factors related to activity and distance traveled in zoos and aquariums. PLoS ONE 2021, 16, e0250687. [Google Scholar] [CrossRef] [PubMed]
- Lauderdale, L.K.; Shorter, K.A.; Zhang, D.; Gabaldon, J.; Mellen, J.D.; Walsh, M.T.; Granger, D.A.; Miller, L.J. Habitat characteristics and animal management factors associated with habitat use by bottlenose dolphins in zoological environments. PLoS ONE 2021, 16, e0252010. [Google Scholar] [CrossRef] [PubMed]
- Lauderdale, L.K.; Walsh, M.T.; Mitchell, K.A.; Granger, D.A.; Mellen, J.D.; Miller, L.J. Health reference intervals and values for common bottlenose dolphins (Tursiops truncatus), Indo-Pacific bottlenose dolphins (Tursiops aduncus), Pacific white-sided dolphins (Lagenorhynchus obliquidens), and beluga whales (Delphinapterus leucas). PLoS ONE 2021, 16, e0250332. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.J.; Lauderdale, L.K.; Bryant, J.L.; Mellen, J.D.; Walsh, M.T.; Granger, D.A. Behavioral diversity as a potential positive indicator of animal welfare in bottlenose dolphins. PLoS ONE 2021, 16, e0253113. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.J.; Lauderdale, L.K.; Mellen, J.D.; Walsh, M.T.; Granger, D.A. Relationships between animal management and habitat characteristics with two potential indicators of welfare for bottlenose dolphins under professional care. PLoS ONE 2021, 16, e0252861. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.J.; Lauderdale, L.K.; Mellen, J.D.; Walsh, M.T.; Granger, D.A. Assessment of animal management and habitat characteristics associated with social behavior in bottlenose dolphins across zoological facilities. PLoS ONE 2021, 16, e0253732. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.J.; Lauderdale, L.K.; Walsh, M.T.; Bryant, J.L.; Mitchell, K.A.; Granger, D.A.; Mellen, J.D. Reference intervals and values for fecal cortisol, aldosterone, and the ratio of cortisol to dehydroepiandrosterone metabolites in four species of cetaceans. PLoS ONE 2021, 16, e0250331. [Google Scholar] [CrossRef] [PubMed]
- Brando, S.; Broom, D.M.; Acasuso-Rivero, C.; Clark, F. Optimal marine mammal welfare under human care: Current efforts and future directions. Behav. Process. 2018, 156, 16–36. [Google Scholar] [CrossRef]
- Mench, J. Environmental enrichment and the importance of exploratory behavior. In Second Nature: Environmental Enrichment for Captive Animals; Shepherdson, J., Mellen, J., Hutchins, M., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1998; pp. 30–46. [Google Scholar]
- Wemelsfelder, F. Boredom and laboratory animal welfare. Exp. Anim. Biomed. Res. 1990, 243–272. [Google Scholar]
- Newberry, R.C. Environmental enrichment- Increasing the biological relevance of captive environments. Appl. Anim. Behav. Sci. 1995, 44, 229–243. [Google Scholar] [CrossRef]
- Boissy, A.; Manteuffel, G.; Jensen, M.B.; Moe, R.O.; Spruijt, B.; Keeling, L.J.; Winckler, C.; Forkman, B.; Dimitrov, I.; Langbein, J.; et al. Assessment of positive emotions in animals to improve their welfare. Physiol. Behav. 2007, 92, 375–397. [Google Scholar] [CrossRef] [PubMed]
- Clark, F.E. Marine mammal cognition and captive care-a proposal for cognitive enrichment in zoos and aquariums. J. Zoo Aquar. Res. 2013, 1, 1–6. [Google Scholar]
- Kuczaj, S.; Lacinak, T.; Fad, O.; Trone, M.; Solangi, M.; Ramos, J. Keeping Environmental Enrichment Enriching. Int. J. Comp. Psychol. 2002; 15, 127–137. [Google Scholar]
- Hirskyj-Douglas, I.; Kankaanpää, V. Exploring How White-Faced Sakis Control Digital Visual Enrichment Systems. Animals 2021, 11, 557. [Google Scholar] [CrossRef] [PubMed]
- Hoy, J.M.; Murray, P.J.; Tribe, A. Thirty years later: Enrichment practices for captive mammals. Zoo Biol. 2010, 29, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Gao, L.; Zhang, X. Environmental enrichment increases aquatic animal welfare: A systematic review and meta-analysis. Rev. Aquac. 2021, 14, 1120–1135. [Google Scholar] [CrossRef]
- Delfour, F.; Faulkner, C.; Carter, T. Object Manipulation and Play Behaviour in Bottlenose Dolphins (Tursiops truncatus) under Human Care. Int. J. Comp. Psychol. 2017, 30. [Google Scholar] [CrossRef]
- Herrelko, E.S.; Vick, S.-J.; Buchanan-Smith, H.M. Cognitive research in zoo-housed chimpanzees: Influence of personality and impact on welfare. Am. J. Primatol. 2012, 74, 828–840. [Google Scholar] [CrossRef]
- Line, S.W.; Morgan, K.N. The effects of two novel objects on the behavior of singly caged adult rhesus macaques. Lab. Anim. Sci. 1991, 41, 365–369. [Google Scholar]
- EEskelinen, H.; Winship, K.; Borger-Turner, J. Sex, Age, and Individual Differences in Bottlenose Dolphins (Tursiops truncatus) in Response to Environmental Enrichment. Anim. Behav. Cogn. 2015, 2, 241–253. [Google Scholar] [CrossRef]
- Winship, K.A.; Eskelinen, H.C. Behavioral responses of two species of dolphins to novel video footage: An exploration of sex differences. Zoo Biol. 2018, 37, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Clay, A.W.; Perdue, B.M.; Gaalema, D.E.; Dolins, F.L.; Bloomsmith, M.A. The use of technology to enhance zoological parks. Zoo Biol. 2011, 30, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Markowitz, H. Applied behavioral analysis and engineering in the zoo, 1988.
- Jenny, S.; Schmid, H. Effect of feeding boxes on the behavior of stereotyping amur tigers (Panthera tigris altaica) in the Zurich Zoo, Zurich, Switzerland. Zoo Biol. 2002, 21, 573–584. [Google Scholar] [CrossRef]
- Kim-McCormack, N.N.; Smith, C.L.; Behie, A.M. Is interactive technology a relevant and effective enrichment for captive great apes? Appl. Anim. Behav. Sci. 2016, 185, 1–8. [Google Scholar] [CrossRef]
- Perdue, B.M.; Evans, T.A.; Washburn, D.A.; Rumbaugh, D.M.; Beran, M.J. Do monkeys choose to choose? Learn. Behav. 2014, 42, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Perdue, B.M.; Beran, M.J.; Washburn, D.A. A computerized testing system for primates: Cognition, welfare, and the Rumbaughx. Behav. Process. 2018, 156, 37–50. [Google Scholar] [CrossRef]
- Washburn, D.A.; Rumbaugh, D.M. Rhesus monkey complex learning skills reassessed. Int. J. Primatol. 1991, 12, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Washburn, D. The Four Cs of Psychological Wellbeing: Lessons from Three Decades of Computer-based Environmental Enrichment. Anim. Behav. Cogn. 2015, 2, 218–232. [Google Scholar] [CrossRef]
- Coe, J.; Hoy, J. Choice, Control and Computers: Empowering Wildlife in Human Care. Multimodal Technol. Interact. 2020, 4, 92. [Google Scholar] [CrossRef]
- Brent; Stone, A.M. M. Long-term use of televisions, balls, and mirrors as enrichment for paired and singly caged chimpanzees. Am. J. Primatol. 1996, 39, 139–145. [Google Scholar] [CrossRef]
- Hanna, P.; Brown, B.; Winship, K.; Cameron, D.; Hill, H.M.; Ii, S.K. A Killer Whale’s (Orcinus orca) Response to Visual Media. Int. J. Comp. Psychol. 2017, 30. [Google Scholar] [CrossRef]
- Waples, K.A.; Gales, N.J. Evaluating and minimising social stress in the care of captive bottlenose dolphins (Tursiops aduncus). Zoo Biol. 2002, 21, 5–26. [Google Scholar] [CrossRef]
- Hart, B.L.; Hart, L.A. Sickness Behavior in Animals: Implications for Health and Wellness. In Encyclopedia of Animal Behavior; Elsevier: Amsterdam, The Netherlands, 2019; pp. 171–175. [Google Scholar]
- Johnson, R.W. The concept of sickness behavior: A brief chronological account of four key discoveries. Vet. Immunol. Immunopathol. 2002, 87, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Clegg, I.L.; Rödel, H.G.; Mercera, B.; Van der Heul, S.; Schrijvers, T.; De Laender, P.; Gojceta, R.; Zimmitti, M.; Verhoeven, E.; Burger, J.; et al. Dolphins’ Willingness to Participate (WtP) in Positive Reinforcement Training as a Potential Welfare Indicator, Where WtP Predicts Early Changes in Health Status. Front. Psychol. 2019, 10, 2112. [Google Scholar] [CrossRef] [PubMed]
- Kastelein, R.; Schooneman, N.; Vaughan, N.; Wiepkema, P. Food consumption and growth of California Sea Lion (Zalophus californianus californianus). Zoo Biol. 2000, 19, 143–159. [Google Scholar] [CrossRef]
- Smith, B.P.; Litchfield, C.A. An empirical case study examining effectiveness of environmental enrichment in two captive Australian Sea Lions (Neophoca cinerea). J. Appl. Anim. Welf. Sci. 2010, 13, 103–122. [Google Scholar] [CrossRef] [PubMed]
- Donald, K.; Benedetti, A.; Goulart, V.D.L.R.; Deming, A.; Nollens, H.; Stafford, G.; Brando, S. Environmental Enrichment Devices Are Safe and Effective at Reducing Undesirable Behaviors in California Sea Lions and Northern Elephant Seals during Rehabilitation. Animals 2023, 13, 1222. [Google Scholar] [CrossRef] [PubMed]
- Hocking, D.P.; Salverson, M.; Evans, A.R. Foraging-Based Enrichment Promotes More Varied Behaviour in Captive Australian Fur Seals (Arctocephalus pusillus doriferus). PLoS ONE 2015, 10, e0124615. [Google Scholar] [CrossRef] [PubMed]
- Kastelein, R.A.; Wiepkema, P.R. The significance of training for the behavior of Steller sea lions (Eumetopias jubata) in human care. Aquat. Mamm. 1988, 14, 39–41. [Google Scholar]
- Samuelson, M.M.; Lauderdale, L.K.; Pulis, K.; Solangi, M.; Hoffland, T.; Lyn, H. Olfactory Enrichment in California Sea Lions (Zalophus californianus): An Effective Tool for Captive Welfare? J. Appl. Anim. Welf. Sci. 2017, 20, 75–85. [Google Scholar] [CrossRef]
- Cook, P.; Reichmuth, C.; Hanke, F.D. The Mind of a Sea Lion, in Ethology and Behavioral Ecology of Otariids and the Odobenid. In Ethology and Behavioral Ecology of Otariids and the Odobenid; Springer: Berlin/Heidelberg, Germany, 2021; pp. 323–345. [Google Scholar]
- Clark, F.E.; Davies, S.L.; Madigan, A.W.; Warner, A.J.; Kuczaj, S.A. Cognitive enrichment for bottlenose Dolphins (Tursiops truncatus): Evaluation of a novel underwater maze device. Zoo Biol. 2013, 32, 608–619. [Google Scholar] [CrossRef] [PubMed]
- Kuczaj, S.A.; Winship, K.A.; Eskelinen, H.C. Can bottlenose dolphins (Tursiops truncatus) cooperate when solving a novel task? Anim. Cogn. 2015, 18, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Winship, K.A.; Ramos, A.; Xitco, M.J., Jr. The Introduction of a Novel Computerized Apparatus to California Sea Lions (Zalophus californianus). Aquat. Mamm. 2023, 49, 73–86. [Google Scholar] [CrossRef]
- Roberts, D.L.; Eskelinen, H.C.; Winship, K.A.; Ramos, A.M.; Xitco, M.J. Effects of Failure on California Sea Lion (Zalophus californianus) Gameplay Strategies and Interest in a Cognitive Task: Implications for Cognitive Enrichment in Pinnipeds. J. Zool. Bot. Gard. 2023, 4, 240–255. [Google Scholar] [CrossRef]
- Lukas, K.E. A review of nutritional and motivational factors contributing to the performance of regurgitation and reingestion in captive lowland gorillas (Gorilla gorilla gorilla). Appl. Anim. Behav. Sci. 1999, 63, 237–249. [Google Scholar] [CrossRef]
- Struck, K.; Videan, E.N.; Fritz, J.; Murphy, J. Attempting to reduce regurgitation and reingestion in a captive chimpanzee through increased feeding opportunities—A case study. Lab. Anim. 2007, 36, 35–38. [Google Scholar] [CrossRef]
- Tidière, M.; Colchero, F.; Staerk, J.; Adkesson, M.J.; Andersen, D.H.; Bland, L.; Böye, M.; Brando, S.; Clegg, I.; Cubaynes, S.; et al. Survival improvements of marine mammals in zoological institutions mirror historical advances in human longevity. Proc. R. Soc. B Biol. Sci. 2023, 290, 20231895. [Google Scholar] [CrossRef]
- Frame, G. Differences in anticipatory behavior and session performance of California sea lions (Zalophus californianus) based on session type. In Rosenstiel School of Marine and Atmospheric Science; University of Miami: Miami, FL, USA, 2021; p. 42. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Animal | Age at Start of Time Period 1 | Classification | Time Period 1 Dates | Time Period 2 Dates | EVE Sessions in Time Period 2 |
|---|---|---|---|---|---|
| ANK | 9 | EVE–Control | 22 July 2018–22 July 2020 | 23 July 2020–23 July 2022 | 93 |
| JCK | 12 | Non-EVE–Control | 20 March 2018–20 March 2020 | 21 March 2020–21 March 2022 * | 0 |
| REX | 15 | EVE–Focal | 8 September 2018–8 September 2020 | 9 September 2020–9 September 2022 | 183 |
| SLD | 16 | EVE–Focal | 2 August 2018–2 August 2020 | 3 August 2020–3 August 2022 | 113 |
| Subject | Time Period | Days Removed Due to Illness | Number of Days Analyzed | Days of ICOD | Average Diet (lbs) | Mean % of Diet Consumed |
|---|---|---|---|---|---|---|
| ANK | 1 | 0 | 747 | 6 | 12.19 | 99.83 |
| 2 | 0 | 743 | 1 | 13.04 | 99.93 | |
| JCK | 1 | 0 | 728 | 6 | 11.58 | 99.89 |
| 2 | 19 | 701 | 8 | 12.47 | 99.86 | |
| REX | 1 | 5 | 717 | 61 | 15.19 | 97.36 |
| 2 | 0 | 717 | 18 | 16.30 | 99.27 | |
| SLD | 1 | 25 | 701 | 40 | 15.16 | 98.06 |
| 2 | 1 | 721 | 15 | 13.23 | 99.37 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Winship, K.; McClain, A.; Ramos, A.; Dunham, J.; Xitco, M., Jr. Health and Welfare Benefits of Computerized Cognitive Enrichment in California Sea Lions (Zalophus californianus) at the US Navy Marine Mammal Program. Animals 2024, 14, 1120. https://doi.org/10.3390/ani14071120
Winship K, McClain A, Ramos A, Dunham J, Xitco M Jr. Health and Welfare Benefits of Computerized Cognitive Enrichment in California Sea Lions (Zalophus californianus) at the US Navy Marine Mammal Program. Animals. 2024; 14(7):1120. https://doi.org/10.3390/ani14071120
Chicago/Turabian StyleWinship, Kelley, Abby McClain, Amber Ramos, Jennifer Dunham, and Mark Xitco, Jr. 2024. "Health and Welfare Benefits of Computerized Cognitive Enrichment in California Sea Lions (Zalophus californianus) at the US Navy Marine Mammal Program" Animals 14, no. 7: 1120. https://doi.org/10.3390/ani14071120
APA StyleWinship, K., McClain, A., Ramos, A., Dunham, J., & Xitco, M., Jr. (2024). Health and Welfare Benefits of Computerized Cognitive Enrichment in California Sea Lions (Zalophus californianus) at the US Navy Marine Mammal Program. Animals, 14(7), 1120. https://doi.org/10.3390/ani14071120