
A Combined Approach for the Characterization of Small Ruminant Lentivirus Strains Circulating in the Islands and Mainland of Greece
 , , , and
, , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
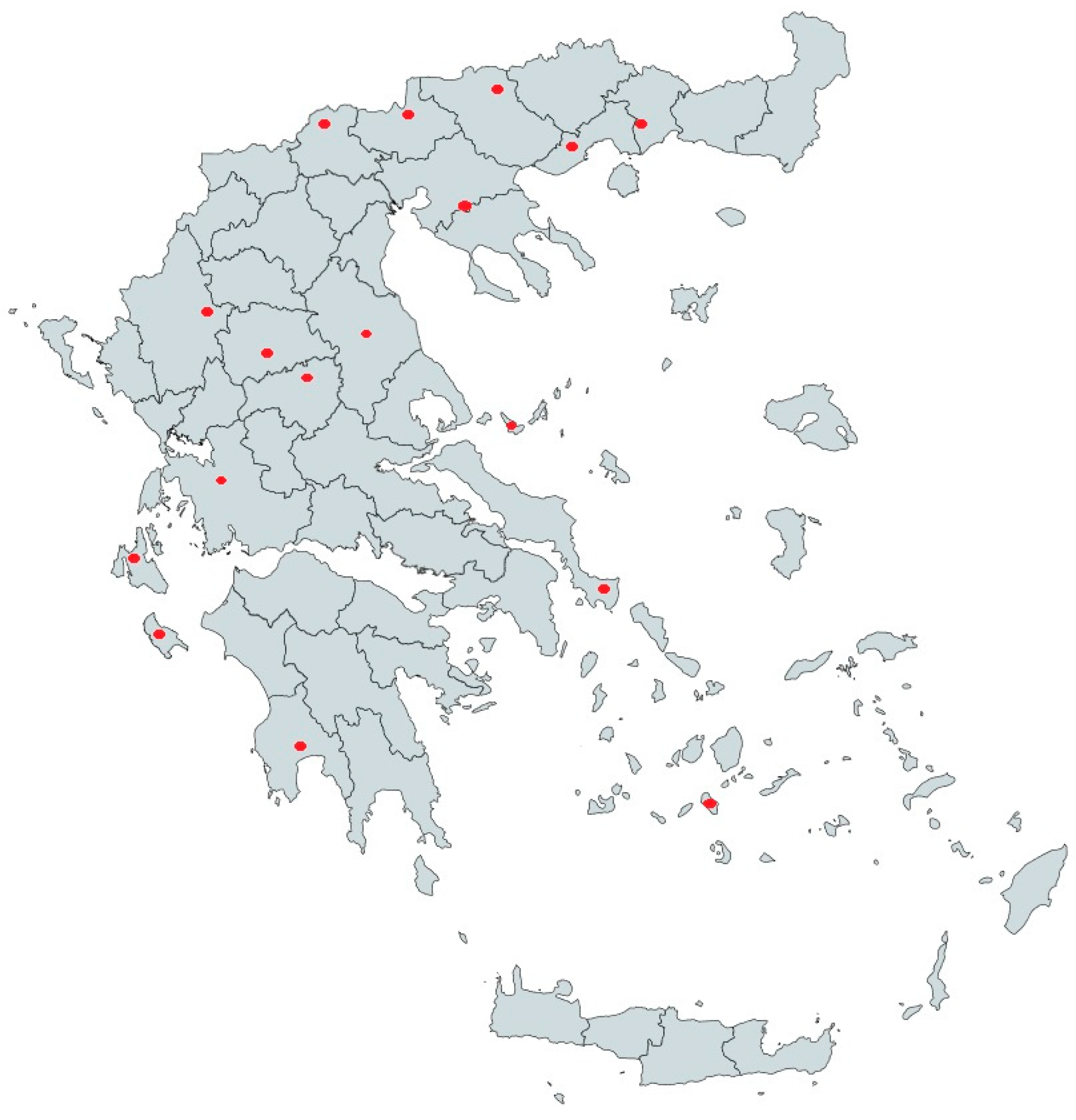
2.1. Sample Selection and Serological Analysis
2.2. DNA Extraction and SRLV Proviral Amplification
2.3. Amplicon Sequencing
2.4. Sequence Analysis
3. Results
3.1. Serology
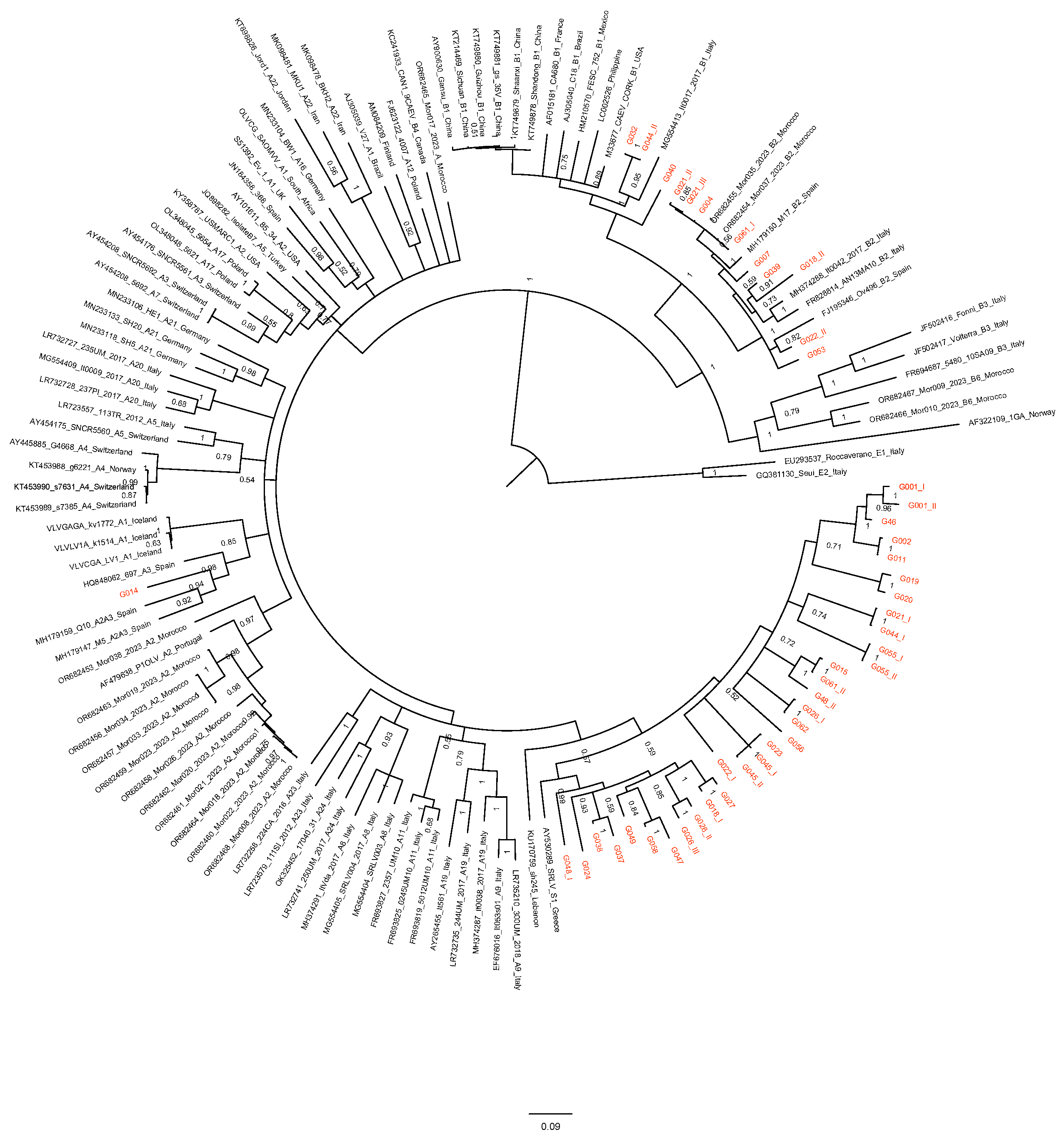
3.2. Sequence Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ramírez, H.; Reina, R.; Amorena, B.; Andrés, D.; Martínez, H. Small Ruminant Lentiviruses: Genetic Variability, Tropism and Diagnosis. Viruses 2013, 5, 1175–1207. [Google Scholar] [CrossRef] [PubMed]
- Leroux, C.; Minardi, C.J.; Mornex, J.-F. SRLVs: A Genetic Continuum of Lentiviral Species in Sheep and Goats with Cumulative Evidence of Cross Species Transmission. Curr. HIV Res. 2010, 8, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Shah, C.; Böni, J.; Huder, J.B.; Vogt, H.R.; Mühlherr, J.; Zanoni, R.; Miserez, R.; Lutz, H.; Schüpbach, J. Phylogenetic Analysis and Reclassification of Caprine and Ovine Lentiviruses Based on 104 New Isolates: Evidence for Regular Sheep-to-Goat Transmission and Worldwide Propagation through Livestock Trade. Virology 2004, 319, 12–26. [Google Scholar] [CrossRef] [PubMed]
- Leroux, C.; Chastang, J.; Greenland, T.; Mornex, J.F. Genomic Heterogeneity of Small Ruminant Lentiviruses: Existence of Heterogeneous Populations in Sheep and of the Samelentiviral Genotypes in Sheep and Goats. Arch. Virol. 1997, 142, 1125–1137. [Google Scholar] [CrossRef] [PubMed]
- de Miguel, R.; Arrieta, M.; Rodríguez-largo, A.; Echeverría, I.; Resendiz, R.; Pérez, E.; Ruiz, H.; Pérez, M.; de Andrés, D.; Reina, R.; et al. Worldwide Prevalence of Small Ruminant Lentiviruses in Sheep: A Systematic Review and Meta-Analysis. Animals 2021, 11, 784. [Google Scholar] [CrossRef] [PubMed]
- Angelopoulou, K.; Karanikolaou, K.; Papanastasopoulou, M.; Koumpati-Artopiou, M.; Vlemmas, I.; Papadopoulos, O.; Koptopoulos, G. First Partial Characterisation of Small Ruminant Lentiviruses from Greece. Vet. Microbiol. 2005, 109, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rossi, R. Briefing European Parliamentary Research Service. 2017. Available online: https://www.europarl.europa.eu/RegData/etudes/BRIE/2017/608663/EPRS_BRI(2017)608663_EN.pdf (accessed on 1 March 2024).
- Karanikolaou, K.; Angelopoulou, K.; Papanastasopoulou, M.; Koumpati-Artopiou, M.; Papadopoulos, O.; Koptopoulos, G. Detection of Small Ruminant Lentiviruses by PCR and Serology Tests in Field Samples of Animals from Greece. Small Rumin. Res. 2005, 58, 181–187. [Google Scholar] [CrossRef]
- Ploumi, K.; Christodoulou, V.; Vainas, E.; Lymberopoulos, A.; Xioufis, A.; Giouzeijiannis, A.; Paschaleri, E.; Ap Dewi, I. Effect of Maedi-Visna Virus Infection on Milk Production in Dairy Sheep in Greece. Vet. Rec. 2001, 149, 526–527. [Google Scholar] [CrossRef] [PubMed]
- Kominakis, A.; Tarsani, E.; Hager-Theodorides, A.L.; Mastranestasis, I.; Gkelia, D.; Hadjigeorgiou, I. Genetic Differentiation of Mainland-Island Sheep of Greece: Implications for Identifying Candidate Genes for Long-Term Local Adaptation. PLoS ONE 2021, 16, e0257461. [Google Scholar] [CrossRef]
- Reina, R.; Berriatua, E.; Luján, L.; Juste, R.; Sánchez, A.; de Andrés, D.; Amorena, B. Prevention Strategies against Small Ruminant Lentiviruses: An Update. Vet. J. 2009, 182, 31–37. [Google Scholar] [CrossRef]
- De Andrés, D.; Klein, D.; Watt, N.J.; Berriatua, E.; Torsteinsdottir, S.; Blacklaws, B.A.; Harkiss, G.D. Diagnostic Tests for Small Ruminant Lentiviruses. Vet. Microbiol. 2005, 107, 49–62. [Google Scholar] [CrossRef] [PubMed]
- de Andrés, X.; Ramírez, H.; Bertolotti, L.; San Román, B.; Glaria, I.; Crespo, H.; Jáuregui, P.; Minguijón, E.; Juste, R.; Leginagoikoa, I.; et al. An Insight into a Combination of ELISA Strategies to Diagnose Small Ruminant Lentivirus Infections. Vet. Immunol. Immunopathol. 2013, 152, 277–288. [Google Scholar] [CrossRef] [PubMed]
- WOAH. Terrestrial Manual Online Access World Organisation for Animal Health; WOAH—World Organisation for Animal Health: Paris, France, 2022; pp. 1–49. [Google Scholar]
- Olech, M. The Genetic Variability of Small-Ruminant Lentiviruses and Its Impact on Tropism, the Development of Diagnostic Tests and Vaccines and the Effectiveness of Control Programmes. J. Vet. Res. 2023, 67, 479–502. [Google Scholar] [CrossRef] [PubMed]
- Michiels, R.; Adjadj, N.R.; De Regge, N. Phylogenetic Analysis of Belgian Small Ruminant Lentiviruses Supports Cross Species Virus Transmission and Identifies New Subtype B5 Strains. Pathogens 2020, 9, 183. [Google Scholar] [CrossRef] [PubMed]
- Minguijón, E.; Reina, R.; Pérez, M.; Polledo, L.; Villoria, M.; Ramírez, H.; Leginagoikoa, I.; Badiola, J.J.; García-Marín, J.F.; de Andrés, D.; et al. Small Ruminant Lentivirus Infections and Diseases. Vet. Microbiol. 2015, 181, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Panneum, S.; Rukkwamsuk, T. Diagnosis of Caprine Arthritis Encephalitis Virus Infection in Dairy Goats by ELISA, PCR and Viral Culture. Pol. J. Vet. Sci. 2017, 20, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Herrmann-Hoesing, L.M. Diagnostic Assays Used to Control Small Ruminant Lentiviruses. J. Vet. Diagn. Investig. 2010, 22, 843–855. [Google Scholar] [CrossRef] [PubMed]
- Leroux, C.; Lerondelle, C.; Chastang, J.; Mornex, J.-F. RT-PCR Detection of Lentiviruses in Milk or Mammary Secretions of Sheep or Goats from Infected Flocks. Vet. Res. 1997, 28, 115–121. [Google Scholar]
- Potârniche, A.V.; Czopowicz, M.; Szaluś-Jordanow, O.; Moroz-Fik, A.; Mickiewicz, M.; Biernacka, K.; Witkowski, L.; Markowska-Daniel, I.; Bagnicka, E.; Cerbu, C.; et al. Serological Testing of an Equal-Volume Milk Sample—A New Method to Estimate the Seroprevalence of Small Ruminant Lentivirus Infection? BMC Vet. Res. 2023, 19, 43. [Google Scholar] [CrossRef]
- Brinkhof, J.M.A.; Houwers, D.J.; Moll, L.; Dercksen, D.; van Maanen, C. Diagnostic Performance of ELISA and PCR in Identifying SRLV-Infected Sheep and Goats Using Serum, Plasma and Milk Samples and in Early Detection of Infection in Dairy Flocks through Bulk Milk Testing. Vet. Microbiol. 2010, 142, 193–198. [Google Scholar] [CrossRef]
- Reina, R.R.R.; Bertolotti, L.; Dei Giudici, S.; Puggioni, G.; Ponti, N.; Profiti, M.; Patta, C.; Rosati, S. Small Ruminant Lentivirus Genotype E Is Widespread in Sarda Goat. Vet. Microbiol. 2010, 144, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Adjadj, N.R.; Vicca, J.; Michiels, R.; De Regge, N. (Non-)Sense of Milk Testing in Small Ruminant Lentivirus Control Programs in Goats. Comparative Analysis of Antibody Detection and Molecular Diagnosis in Blood and Milk. Viruses 2019, 12, 3. [Google Scholar] [CrossRef] [PubMed]
- Colitti, B.; Coradduzza, E.; Puggioni, G.; Capucchio, M.T.M.T.; Reina, R.; Bertolotti, L.; Rosati, S. A New Approach for Small Ruminant Lentivirus Full Genome Characterization Revealed the Circulation of Divergent Strains. PLoS ONE 2019, 14, e0212585. [Google Scholar] [CrossRef] [PubMed]
- Colitti, B.; Daif, S.; Choukri, I.; Scalas, D.; Jerre, A.; El Berbri, I.; Fihri, O.F.; Rosati, S. Serological and Molecular Characterization of Small Ruminant Lentiviruses in Morocco. Animals 2024, 14, 550. [Google Scholar] [CrossRef] [PubMed]
- Olech, M.; Rachid, A.; Croisé, B.; Kuźmak, J.; Valas, S. Genetic and Antigenic Characterization of Small Ruminant Lentiviruses Circulating in Poland. Virus Res. 2012, 163, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Grego, E.; Bertolotti, L.; Quasso, A.; Profiti, M.; Lacerenza, D.; Muz, D.; Rosati, S. Genetic Characterization of Small Ruminant Lentivirus in Italian Mixed Flocks: Evidence for a Novel Genotype Circulating in a Local Goat Population. J. Gen. Virol. 2007, 88, 3423–3427. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S.; Krueger, F.; Seconds-Pichon, A.; Biggins, F.; Wingett, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Babraham Bioinformatics. Babraham Inst. 2015, 1, 1. [Google Scholar]
- Zerbino, D.R.; Birney, E. Velvet: Algorithms for de Novo Short Read Assembly Using de Bruijn Graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- De Azevedo, D.A.A.; dos Santos, V.W.S.; de Sousa, A.L.M.; Peixoto, R.M.; Pinheiro, R.R.; Andrioli, A.; das Teixeira, M.F. Small Ruminant Lentiviruses: Economic and Productive Losses, Consequences of the Disease. Arq. Inst. Biol. 2018, 84. [Google Scholar] [CrossRef]
- Michiels, R.; Van Mael, E.; Quinet, C.; Welby, S.; Cay, A.B.; De Regge, N. Seroprevalence and Risk Factors Related to Small Ruminant Lentivirus Infections in Belgian Sheep and Goats. Prev. Vet. Med. 2018, 151, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Pérez, M.; Biescas, E.; de Andrés, X.; Leginagoikoa, I.; Salazar, E.; Berriatua, E.; Reina, R.; Bolea, R.; de Andrés, D.; Juste, R.A.; et al. Visna/Maedi Virus Serology in Sheep: Survey, Risk Factors and Implementation of a Successful Control Programme in Aragón (Spain). Vet. J. 2010, 186, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Giammarioli, M.; Bazzucchi, M.; Puggioni, G.; Brajon, G.; Dei Giudici, S.; Taccori, F.; Feliziani, F.; De Mia, G.M. Phylogenetic Analysis of Small Ruminant Lentivirus (SRLV) in Italian Flocks Reveals the Existence of Novel Genetic Subtypes. Virus Genes 2011, 43, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Bazzucchi, M.; Pierini, I.; Gobbi, P.; Pirani, S.; Torresi, C.; Iscaro, C.; Feliziani, F.; Giammarioli, M. Genomic Epidemiology and Heterogeneity of SRLV in Italy from 1998 to 2019. Viruses 2021, 13, 2338. [Google Scholar] [CrossRef] [PubMed]
- Molaee, V.; Bazzucchi, M.; De Mia, G.M.; Otarod, V.; Abdollahi, D.; Rosati, S.; Lühken, G. Phylogenetic Analysis of Small Ruminant Lentiviruses in Germany and Iran Suggests Their Expansion with Domestic Sheep. Sci. Rep. 2020, 10, 2243. [Google Scholar] [CrossRef] [PubMed]
- Arcangeli, C.; Torricelli, M.; Sebastiani, C.; Lucarelli, D.; Ciullo, M.; Passamonti, F.; Giammarioli, M.; Biagetti, M. Genetic Characterization of Small Ruminant Lentiviruses (SRLVs) Circulating in Naturally Infected Sheep in Central Italy. Viruses 2022, 14, 686. [Google Scholar] [CrossRef]
- Lianou, D.T.; Michael, C.K.; Fthenakis, G.C. Data on Mapping 444 Dairy Small Ruminant Farms during a Countrywide Investigation Performed in Greece. Animals 2023, 13, 2044. [Google Scholar] [CrossRef]
- Kalogianni, A.I.; Bouzalas, I.; Bossis, I.; Gelasakis, A.I. A Longitudinal Cohort Study of Risk Factors Associated with Small Ruminant Lentivirus Seropositivity in Intensively Reared Dairy Ewes in Greece. Pathogens 2023, 12, 1200. [Google Scholar] [CrossRef]
- Nobrega, D.B.; French, J.E.; Kelton, D.F. A Scoping Review of the Testing of Bulk Tank Milk to Detect Nonbacterial Pathogens or Herd Exposure to Nonbacterial Pathogens in Dairy Cattle. J. Dairy Sci. 2023, 106, 5636–5658. [Google Scholar] [CrossRef] [PubMed]
- Nobrega, D.B.; French, J.E.; Kelton, D.F. A Scoping Review of the Testing of Bulk Milk to Detect Infectious Diseases of Dairy Cattle: Diseases Caused by Bacteria. J. Dairy Sci. 2023, 106, 1986–2006. [Google Scholar] [CrossRef]
- Brito, B.; Hick, P. Milk as a Diagnostic Fluid to Monitor Viral Diseases in Dairy Cattle. Aust. Vet. J. 2024, 102, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Mazzei, M.; Carrozza, M.L.; Bandecchi, P.; Mazzanti, G.; Mannelli, A.; Tolari, F. Evaluation of an ELISA to Detect Antibodies to Maedi-Visna Virus in Individual and Pooled Samples of Milk from Sheep. Vet. Rec. 2005, 157, 552–555. [Google Scholar] [CrossRef] [PubMed]
- Barquero, N.; Arjona, A.; Domenech, A.; Toural, C.; De Las Heras, A.; Fernández-Garayzabal, J.F.; Ruiz-Santa Quiteria, J.A.; Gomez-Lucia, E. Diagnostic Performance of PCR and ELISA on Blood and Milk Samples and Serological Survey for Small Ruminant Lentiviruses in Central Spain. Vet. Rec. 2011, 168, 20–22. [Google Scholar] [CrossRef] [PubMed]
- Acevedo Jiménez, G.E.; Tórtora Pérez, J.L.; Rodríguez Murillo, C.; Arellano Reynoso, B.; Ramírez Álvarez, H. Serotyping versus Genotyping in Infected Sheep and Goats with Small Ruminant Lentiviruses. Vet. Microbiol. 2021, 252, 108931. [Google Scholar] [CrossRef]
- Nardelli, S.; Bettini, A.; Capello, K.; Bertoni, G.; Tavella, A. Eradication of Caprine Arthritis Encephalitis Virus in the Goat Population of South Tyrol, Italy: Analysis of the Tailing Phenomenon during the 2016–2017 Campaign. J. Vet. Diagn. Invest. 2020, 32, 589. [Google Scholar] [CrossRef] [PubMed]
- Cardinaux, L.; Zahno, M.-L.; Deubelbeiss, M.; Zanoni, R.; Vogt, H.-R.; Bertoni, G. Virological and Phylogenetic Characterization of Attenuated Small Ruminant Lentivirus Isolates Eluding Efficient Serological Detection. Vet. Microbiol. 2013, 162, 572–581. [Google Scholar] [CrossRef]
- Glaria, I.; Reina, R.; Crespo, H.; de Andrés, X.; Ramírez, H.; Biescas, E.; Pérez, M.M.; Badiola, J.; Luján, L.; Amorena, B.; et al. Phylogenetic Analysis of SRLV Sequences from an Arthritic Sheep Outbreak Demonstrates the Introduction of CAEV-like Viruses among Spanish Sheep. Vet. Microbiol. 2009, 138, 156–162. [Google Scholar] [CrossRef]
- Pérez, M.; Biescas, E.; Reina, R.; Glaria, I.; Marín, B.; Marquina, A.; Salazar, E.; Álvarez, N.; de Andrés, D.; Fantova, E.; et al. Small Ruminant Lentivirus–Induced Arthritis: Clinicopathologic Findings in Sheep Infected by a Highly Replicative SRLV B2 Genotype. Vet. Pathol. 2015, 52, 132–139. [Google Scholar] [CrossRef]
- Germain, K.; Valas, S. Distribution and Heterogeneity of Small Ruminant Lentivirus Envelope Subtypes in Naturally Infected French Sheep. Virus Res. 2006, 120, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Bertolotti, L.; Mazzei, M.; Puggioni, G.; Carrozza, M.L.; dei Giudici, S.; Muz, D.; Juganaru, M.; Patta, C.; Tolari, F.; Rosati, S. Characterization of New Small Ruminant Lentivirus Subtype B3 Suggests Animal Trade within the Mediterranean Basin. J. Gen. Virol. 2011, 92, 1923–1929. [Google Scholar] [CrossRef] [PubMed]
- Tabet, E.; Tlaige, R.; El Hage, J.; Abi-Rizk, A. The Occurrence of Maedi-Visna Virus in Lebanon. Rev. Sci. Et Tech. De L’oie 2017, 36, 899–903. [Google Scholar] [CrossRef] [PubMed]
- Muz, D.; Oǧuzoǧlu, T.Ç.; Rosati, S.; Reina, R.; Bertolotti, L.; Burgu, I. First Molecular Characterization of Visna/Maedi Viruses from Naturally Infected Sheep in Turkey. Arch. Virol. 2013, 158, 559–570. [Google Scholar] [CrossRef]
- Carrozza, M.L.; Niewiadomska, A.M.; Mazzei, M.; Abi-Said, M.R.; Hué, S.; Hughes, J.; Gatseva, A.; Gifford, R.J. Emergence and Pandemic Spread of Small Ruminant Lentiviruses. Virus Evol. 2023, 9, vead005. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Origin of Samples | Region | Farms Tested | Species | Nr of Animals |
|---|---|---|---|---|
| Mainland | Aitolokarnania | 2 | Goats | 190 |
| 3 | Sheep | 202 | ||
| Mainland | Edessa | 2 | Goats | 500 |
| Mainland | Chalkidiki | 6 | Goats | 3050 |
| 2 | Sheep | 240 | ||
| Island | Ios | 10 | Goats | 706 |
| 3 | Sheep | 175 | ||
| 1 | Mixed | 80 | ||
| Island | Karistos | 1` | Sheep | 70 |
| 1 | Mixed | 354 | ||
| Island | Kefalonia | 4 | Goats | 450 |
| 2 | Sheep | 230 | ||
| Mainland | Kilkis | 1 | Goats | 800 |
| 2 | Sheep | 280 | ||
| Mainland | Larisa | 2 | Goats | 600 |
| 1 | Sheep | 400 | ||
| 1 | Mixed | 310 | ||
| Mainland | Grevena | 3 | Mixed | 1700 |
| Mainland | Serres | 1 | Sheep | 230 |
| Island | Skopelos | 5 | Goats | 510 |
| Mainland | Thessaloniki | 1 | Goats | 640 |
| Mainland | Trikala | 1 | Sheep | 250 |
| Mainland | Xanthi | 1 | Sheep | 200 |
| Island | Zante | 4 | Sheep | 410 |
| Sample ID | Location | Species | ELISA Screening | ELISA Genotyping | PCR | Subtype |
|---|---|---|---|---|---|---|
| G001_I | Karistos | Sheep | Positive | Negative | Positive | A |
| G001_II | “ | “ | “ | “ | “ | A |
| G002 | Karistos | Mixed | Positive | B | Positive | A |
| G004 | Ios | Sheep | Positive | B | Positive | B2 |
| G006 | Ios | Goat | Positive | Negative | Negative | - |
| G007 | Ios | Goat | Positive | B | Positive | B2 |
| G011 | Ios | Goat | Positive | AB | Negative | A |
| G014 | Ios | Goat | Positive | AB | Positive | A3 |
| G015 | Ios | Sheep | Positive | Indeterminate | Positive | A |
| G018_I | Zante | Sheep | Positive | A | Positive | A |
| G018_II | “ | “ | “ | “ | “ | B2 |
| G019 | Zante | Sheep | Positive | Indeterminate | Positive | A |
| G020 | Zante | Sheep | Positive | Indeterminate | Positive | A |
| G021_I | Grevena | Mixed | Positive | A | Positive | A |
| G021_II | “ | “ | “ | “ | “ | B2 |
| G021_III | “ | “ | “ | “ | “ | B2 |
| G022_I | Grevena | Mixed | Positive | A | Positive | A |
| G022_II | “ | “ | “ | “ | “ | B2 |
| G023 | Grevena | Mixed | Positive | A | Positive | A |
| G024 | Chalkidiki | Goat | Positive | Indeterminate | Positive | A |
| G025 | Chalkidiki | Goat | Positive | Negative | Negative | - |
| G026_I | Chalkidiki | Goat | Positive | A | Positive | A |
| G026_II | “ | “ | “ | “ | “ | A |
| G026_III | “ | “ | “ | “ | “ | A |
| G027 | Chalkidiki | Goat | Positive | A | Positive | A |
| G032 | Kefalonia | Goat | Positive | B | Positive | B1 |
| G037 | Kilkis | Sheep | Positive | Indeterminate | Positive | A |
| G038 | Kilkis | Sheep | Positive | Indeterminate | Positive | A |
| G039 | Kilkis | Goat | Positive | AB | Positive | B2 |
| G040 | Xanthi | Sheep | Positive | AB | Positive | B1 |
| G041 | Edessa | Goat | Positive | Negative | Negative | - |
| G043 | Chalkidiki | Sheep | Positive | A | Positive | - |
| G044_I | Skopelos | Goat | Negative | Negative | Doubt | A |
| G044_II | “ | “ | “ | “ | “ | B1 |
| G045_I | Skopelos | Goat | Negative | Negative | Doubt | A |
| G045_II | “ | “ | “ | “ | “ | A |
| G046 | Skopelos | Goat | Negative | Negative | Doubt | A |
| G047 | Edessa | Goat | Doubt | Indeterminate | Positive | A |
| G048_I | Serres | Goat | Doubt | Negative | Positive | A |
| G048_II | “ | “ | “ | “ | “ | A |
| G049 | Larisa | Mixed | Positive | Positive | A | |
| G051 | Aitolokarnania | Goat | Doubt | Negative | Negative | - |
| G052 | Larisa | Goat | Positive | A | - | - |
| G053 | Larisa | Sheep | Positive | A | Positive | B2 |
| G055_I | Aitolokarnania | Sheep | Positive | Negative | Positive | A |
| G055_II | “ | “ | “ | “ | “ | A |
| G056 | Aitolokarnania | Sheep | Positive | A | Positive | A |
| G058 | Trikala | Sheep | Positive | Indeterminate | Doubt | A |
| G061_I | Kilkis | Sheep swab | Not applicable | Not applicable | Positive | B2 |
| G061_II | “ | “ | “ | “ | “ | A |
| G062 | Messinia | Sheep swab | Not applicable | Not applicable | Positive | A |
| Strain | Subtype | Sequence |
|---|---|---|
| VLVLV1A_k1514_Iceland | A1 | KLNEEAERWVRQNPPGP |
| AY101611_85-34_USA | A2 | ................. |
| AY530289_SRLV-S1_Greece | ................. | |
| G001_I | ................. | |
| G001_II | ................. | |
| G011 | ................. | |
| G018_I | ................. | |
| G022_I | ................. | |
| G026_II | ................. | |
| G026_III | ................. | |
| G045_I | ................. | |
| G045_II | ................. | |
| G046 | ................. | |
| G048_I | ................. | |
| G055 | ................. | |
| G058 | ................. | |
| G061_II | ................. | |
| G015 | ................. | |
| G019 | ................. | |
| G020 | ................. | |
| G023 | ................. | |
| G027 | ................. | |
| G037 | ................. | |
| G038 | ................. | |
| MN233104_IIW1_Germany | A16 | .........I....... |
| MG554409_It0009_2017_Italy | A20 | .........I....... |
| AY530290_SRLV-S2_Greece | A | .........I....... |
| AF479638_P1OLV_Portugal | A2 | ..............GPN |
| AY530292_SRLV-S4_Greece | A | R................ |
| G002 | R................ | |
| G021_I | R................ | |
| G026_I | R................ | |
| G044_I | R................ | |
| G062 | R................ | |
| G024 | R................ | |
| G049 | R................ | |
| AM084209_Finland | ...D............. | |
| G48_II | ...D............. | |
| G056 | ...D............. | |
| AY445885_G4668_Switzerland | A4 | ..............A.. |
| G047 | ..............Q.. | |
| AJ305039_V27_Brazil | A1 | ................Q |
| OR666874_RO46_Romania | A | ................Q |
| G014 | ................Q | |
| KC241933_CAN1_9CAEV_Canada | B4 | ................R |
| AF015181_CA680_France | B1 | ......D..R.N....G |
| M33677_CAEV_CORK_USA | B1 | .........R.N...P. |
| G044_II | .........R.N...P. | |
| G032 | .........R.N...P. | |
| G040 | .........R.N...P. | |
| LC002526_Philippine | B | ......D..R.N...P. |
| FJ195346_Ov496_Spain | B2 | .........R.N...PQ |
| G021_II | .........R.N...PQ | |
| G021_III | .........R.N...PQ | |
| G022_II | .........R.N...PQ | |
| G061_I | .........R.N...PQ | |
| G039 | .........R.N...PQ | |
| G053 | .........R.N...PQ | |
| G07 | .........R.N...PQ | |
| JF502416_Fonni_Italy | B3 | .........R.N...PA |
| JF502417_Volterra_Italy | B3 | .........R.N...PA |
| G018_II | .........KKN...PQ | |
| G04 | .......G.R.N...PQ | |
| AF322109_1GA_Norway | C | .........R....QPA |
| EU293537_Roccaverano_Italy | E1 | ...K...T.M....QP. |
| GQ381130_Seui_Italy | E2 | ...K...T.M....QP. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouzalas, I.; Apostolidi, E.D.; Scalas, D.; Davidopoulou, E.; Chassalevris, T.; Rosati, S.; Colitti, B. A Combined Approach for the Characterization of Small Ruminant Lentivirus Strains Circulating in the Islands and Mainland of Greece. Animals 2024, 14, 1119. https://doi.org/10.3390/ani14071119
Bouzalas I, Apostolidi ED, Scalas D, Davidopoulou E, Chassalevris T, Rosati S, Colitti B. A Combined Approach for the Characterization of Small Ruminant Lentivirus Strains Circulating in the Islands and Mainland of Greece. Animals. 2024; 14(7):1119. https://doi.org/10.3390/ani14071119
Chicago/Turabian StyleBouzalas, Ilias, Evangelia D. Apostolidi, Daniela Scalas, Evangelia Davidopoulou, Taxiarchis Chassalevris, Sergio Rosati, and Barbara Colitti. 2024. "A Combined Approach for the Characterization of Small Ruminant Lentivirus Strains Circulating in the Islands and Mainland of Greece" Animals 14, no. 7: 1119. https://doi.org/10.3390/ani14071119
APA StyleBouzalas, I., Apostolidi, E. D., Scalas, D., Davidopoulou, E., Chassalevris, T., Rosati, S., & Colitti, B. (2024). A Combined Approach for the Characterization of Small Ruminant Lentivirus Strains Circulating in the Islands and Mainland of Greece. Animals, 14(7), 1119. https://doi.org/10.3390/ani14071119