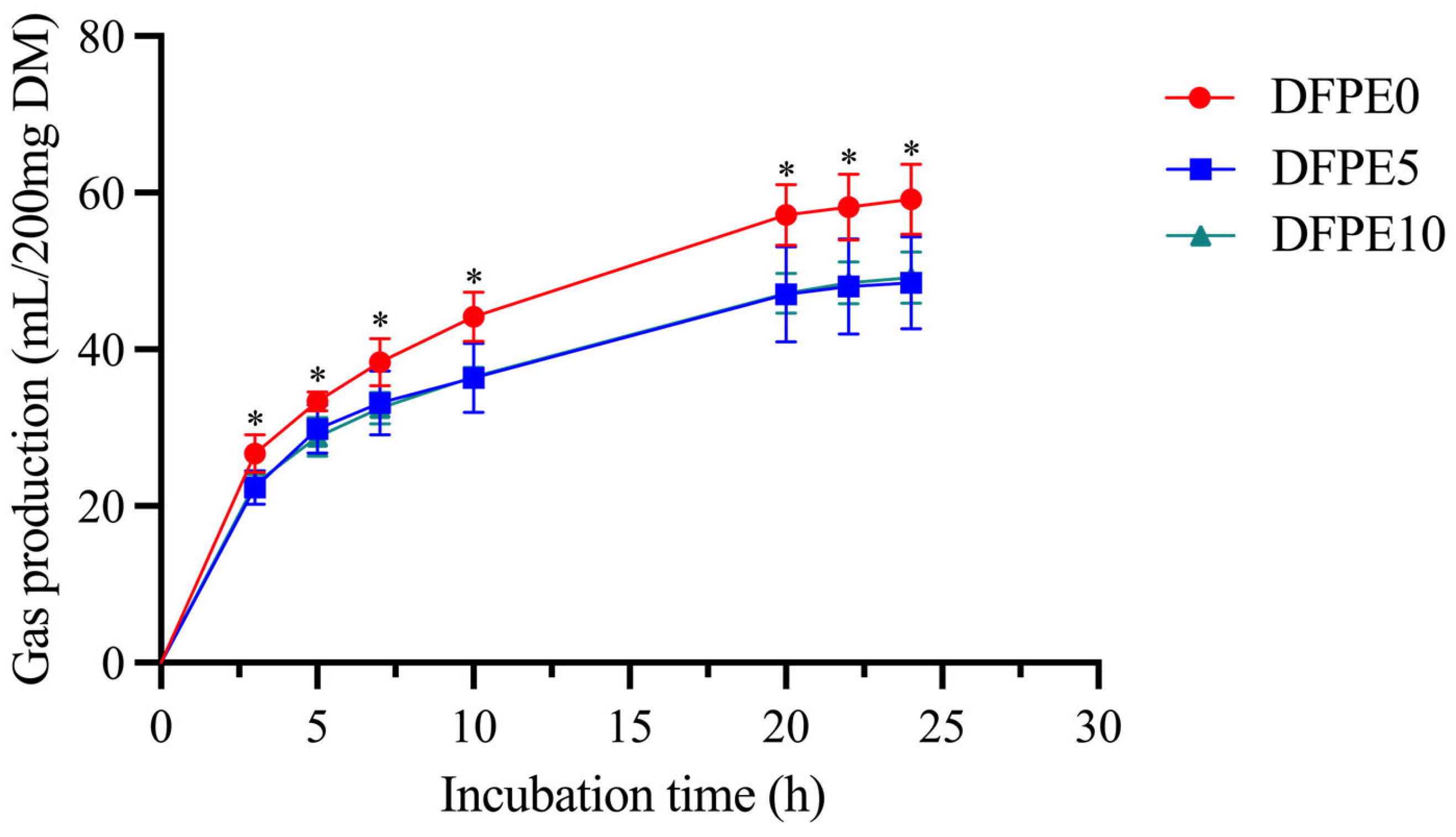
4.1. Effect of DFPE on Gas Production and Rumen Fermentation
GP serves as a crucial indicator of carbohydrate digestion, exhibiting a negative correlation with NDF content and a positive correlation with starch levels [
19]. An excess of GP can contribute to impaired ruminal function, particularly under highly concentrated feeding conditions, leading to the likelihood of cattle and sheep experiencing a subacute rumen bloat state [
20]. Polyphenols demonstrate potential in preventing rumen bloat [
21]. As anticipated, our finding suggests that DFPE has the ability to reduce GP in vitro. A previous study [
22] has reviewed that these compounds possess the capability to reduce GP and CH
4. Yu et al. [
23] reported that naringin could reduce CH
4 production in in vitro rumen fermentation. This reduction in GP may be attributed to the presence of phenolic compounds in DFPE, including naringin, caffeic acid, vanillic acid, and quercetin (
Table 2), which may contribute to this reduction in GP. Furthermore, we observed that supplement levels did not influence GP. One possible reason may be that some gas-producing bacteria (e.g., methanogens) have varying degrees of sensitivity to DFPE, which necessitates further investigation in the future.
On the other hand, phenolic compounds can induce the precipitation of soluble proteins by forming hydrogen bonds [
24]. The correlation of this phenomenon with alleviating tissue swelling has been substantiated [
25]. A previous study demonstrated that 10 g/kg of polyphenolic extract from
Eucommia ulmoides leaf reduced ruminal NH
3-N levels from 175.6 to 129.6 mg/L [
26]. Yu et al. [
23] reported that naringin (20 mg/kg of diet) had decreased NH
3-N from 13.05 mg/dL to 11.41 mg/dL in dairy cows. Conversely, the protein-binding property of phenolic compounds can mitigate ruminal fermentation, thereby enhancing nitrogen use efficiency [
27]. In the present study, our observations indicate that DFPE reduced NH
3-N levels, implying that DFPE has the ability to mitigate the degradation of dietary protein in the rumen and has the potential to enhance protein utilization.
Ruminal pH serves as a valuable indicator for assessing both ruminal health and optimal function. Consistent with the findings reported by Matra et al. [
7], no significant differences in pH were observed in this study, suggesting that DFPE has the capability to maintain an appropriate acid–base status without inducing rumen acidosis. In contrast, the pH observed in this experiment ranges from 6.12 to 6.17, which differs from the optimal pH range (6.2–7.2) for microbial activity. As stated by Jackson et al. [
28], this deviation may be attributed to constraints inherent in the in vitro fermentation system used in this experiment, wherein fermentation by-products such as VFA cannot be absorbed by the artificial rumen, leading to their accumulation and subsequent pH reduction. Nevertheless, the pH values remained within the normal range [
29].
VFA are end-products of carbohydrates and provide the main source of energy for ruminant metabolism. Polyphenols have the potential to increase VFA by modifying the rumen bacteria population and increasing bacterial richness and diversity [
30,
31]. Gao et al. [
32] reported that feeding beef bulls with polyphenol-rich red cabbage extract (120 g/d/head) resulted in a significant increase in total VFA from 83.06 mmol/L to 89.19 mmol/L. In the present study, we found that DFPE significantly increased total VFA. Similarly, previous studies [
33,
34] have reported that quercetin can increase ruminal VFA and decrease GP in vitro. In contrast, the present study revealed a comparatively lower total VFA content (10.78 to 11.95 mmol/L), while Sun et al. reported rumen total VFA ranges from 13.17 to 23.59 mmol/L (in vivo) [
35]. This may be related to the lower microbial concentration in vitro than in vivo [
36].
Butyric acid accounts for about 70% of the daily energy metabolism of ruminants [
37]. Higher butyrate concentrations also enhance ruminal structure and function, enhance nutrient absorption, promote gut health, and improve health benefits for ruminants [
38]. Remling et al. [
39] noted that butyric acid also stimulated the growth of the ruminal papilla, thereby increasing the absorptive surface area of ruminants. In the present study, we found that DFPE had no effect on acetic acid, propionic acid, or butyric acid. A similar result was found by Silva et al. [
40]. On the other hand, branched-chain VFA (BCVFA), such as isobutyric acid, isovaleric acid, and valeric acid, is utilized by ruminal microbes as a source of carbon skeleton for synthesizing branched-chain amino acids [
41]. In the present study, we found higher concentrations of isovaleric acid and valeric acid from DFPE treatments. This indicates that DFPE has the potential to augment the population of cellulolytic bacteria and improve fiber digestibility.
4.2. Effect of DFPE on Bacteria Communities In Vitro
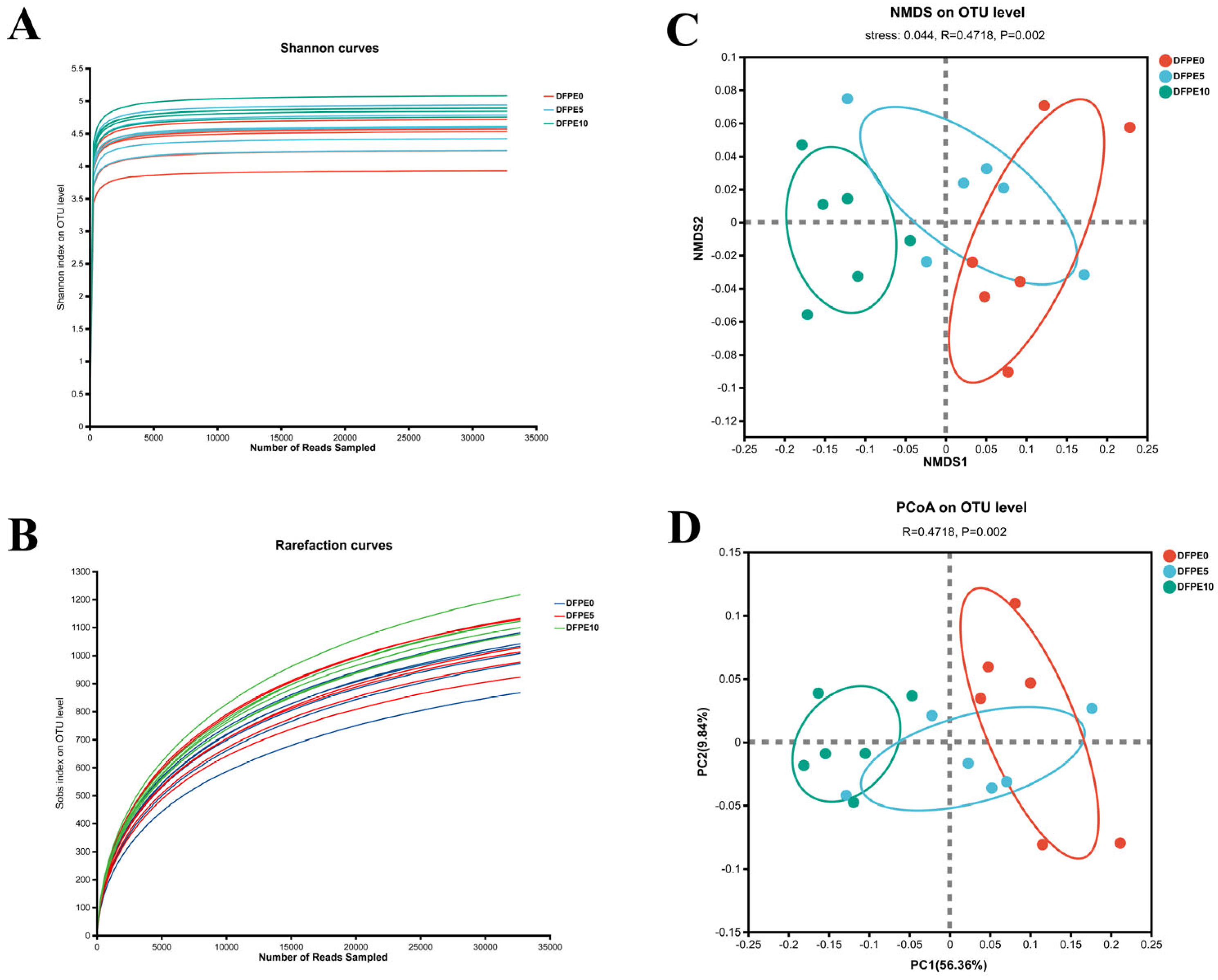
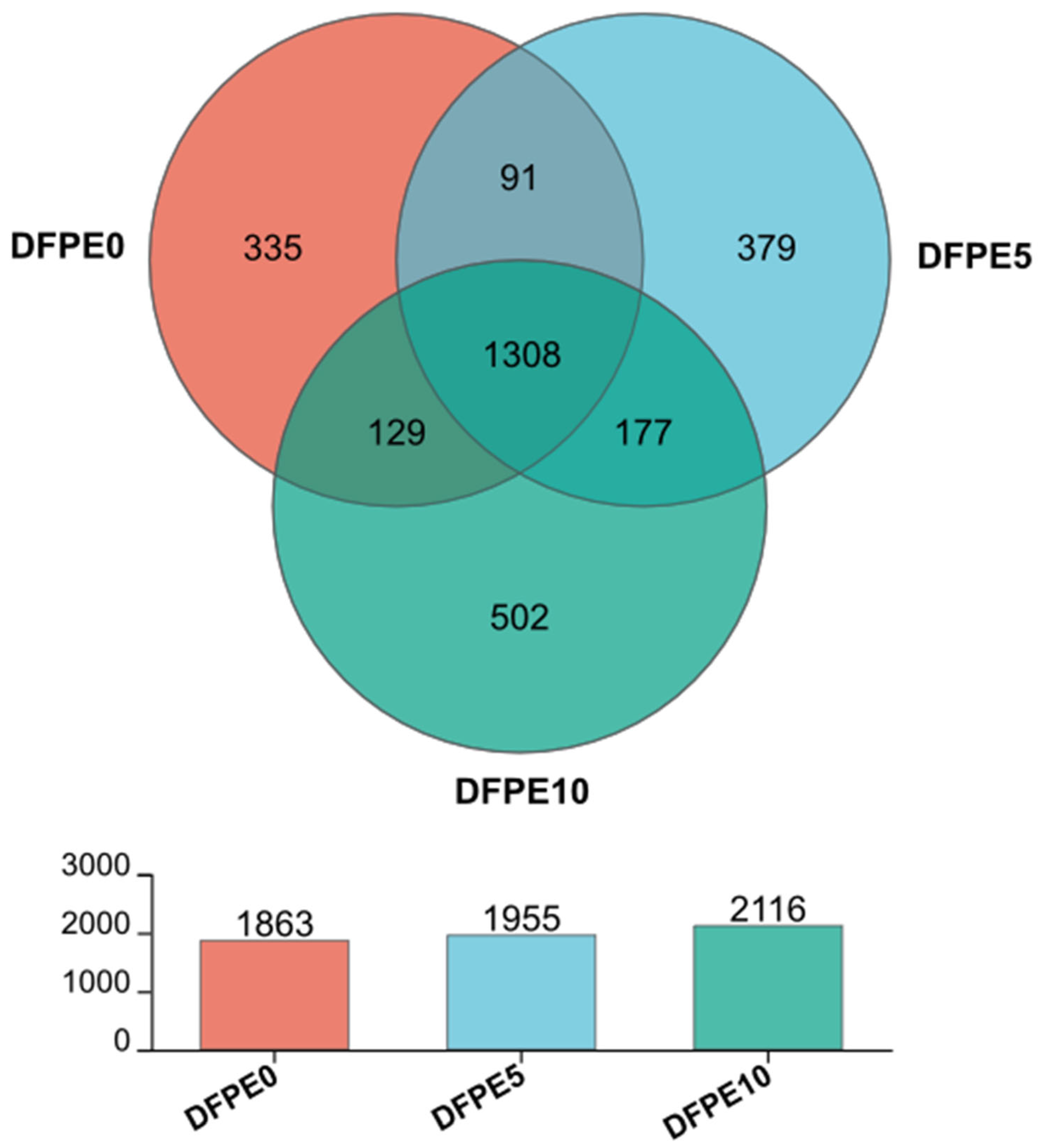
The well-being of the rumen is, in part, contingent upon the appropriate richness, diversity, and stability exhibited by the ruminal microbiome. The alpha diversity indexes of Sobs, Shannon, and Ace were increased by DFPE supplementation. In the present study, we examined the rumen microbiota using 16S rRNA sequencing analysis. As previously elucidated and documented in Sika deer [
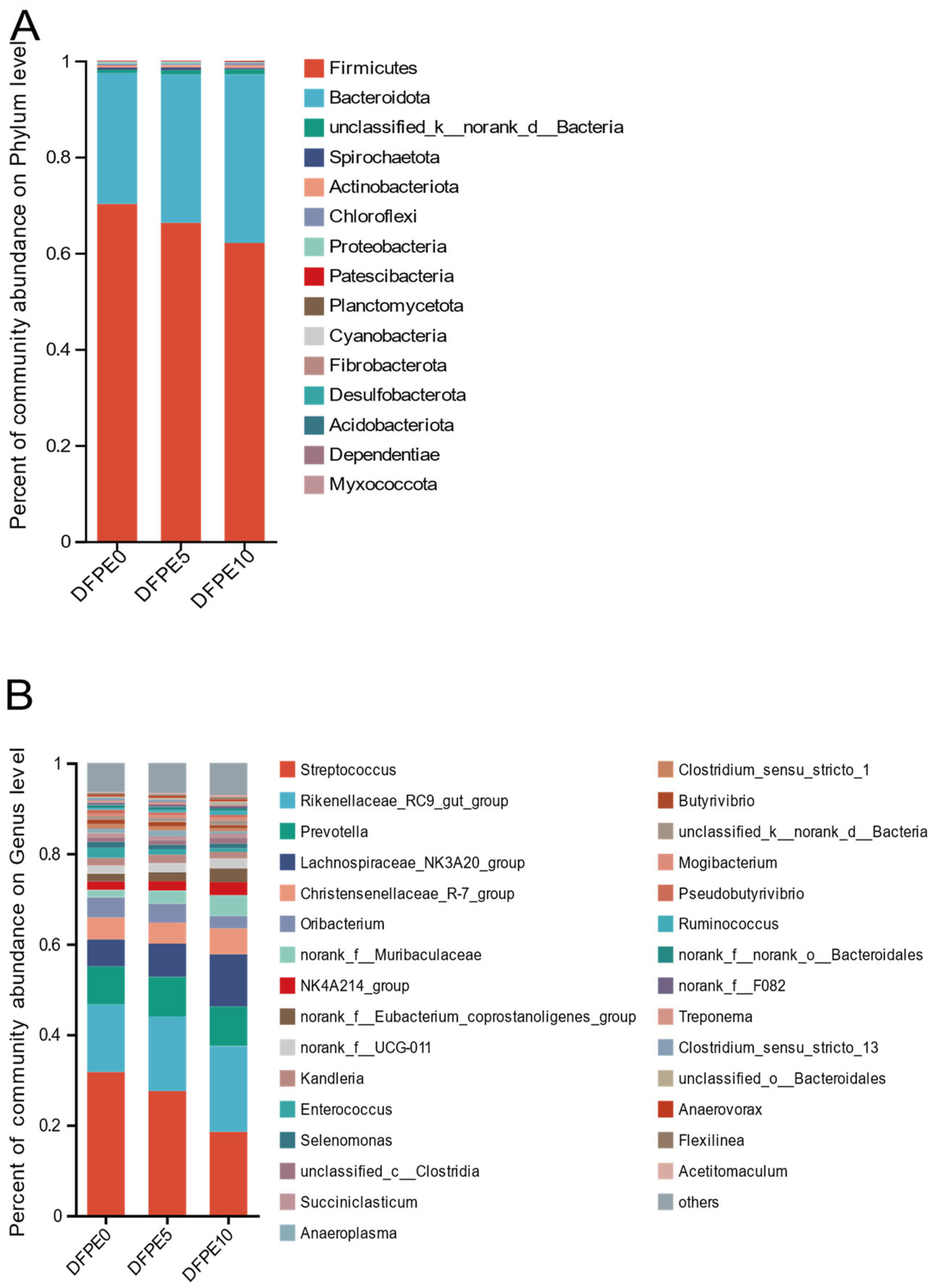
42], we observed that the predominant bacteria belonged to the phyla
Firmicutes and
Bacteroidota, with a high relative ratio of
Firmicutes to
Bacteroidetes in this study. This occurrence may be attributed to the fact that Sika deer typically inhabit cold and high-altitude regions [
43], where a high ratio of
Firmicutes to
Bacteroidetes is advantageous for fat deposition, helping animals adapt to severe cold climates [
44], thereby enhancing their resilience to these challenging conditions. Polyphenols are regarded as candidate compounds for prebiotics due to their interactions with gut microbiota [
45]. The observations from the present study revealed that the relative abundance of
Bacteroidetes increased with the inclusion of DFPE supplementation, while
Firmicutes exhibited a significant decrease. In agreement with our study, Sutoyo et al. [
46] reported that dietary intake of phenolic compounds is associated with a reduced proportion of
Firmicutes and an increased proportion of
Bacteroidetes.
Bacteroidetes are primarily involved with the degradation of carbohydrates, fats, and proteins. At the
Bacteroidetes phylum,
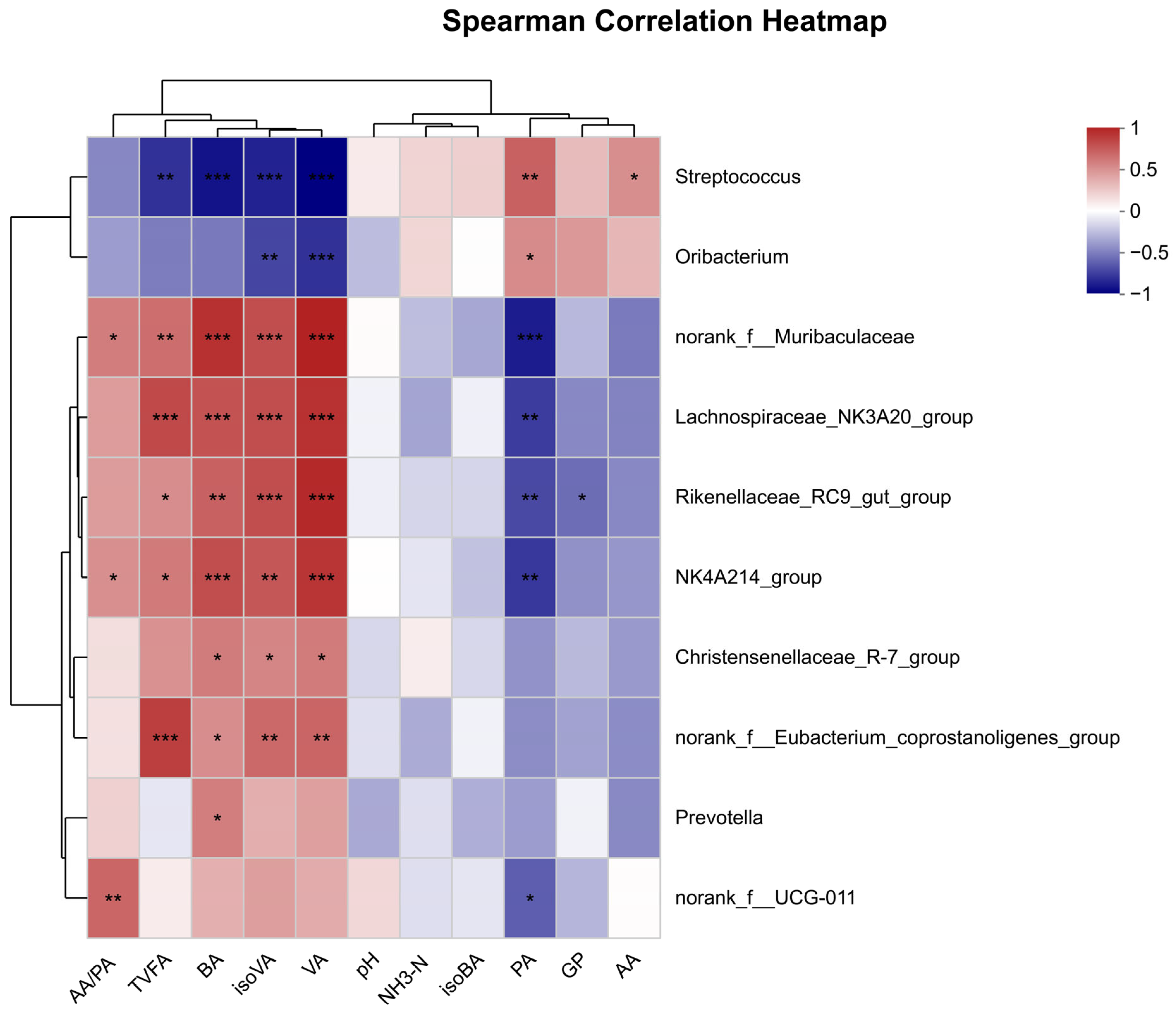
Rikenellaceae_RC9_gut_group,
Prevotella, and
norank_f_Muribaculaceae emerged as the predominant bacterial genera in our study, consistent with findings from a previous study by Si et al. [
47]. The family
Rikenellaceae participates in the degradation of structural carbohydrates, produces succinate and propionate as fermentation end products, and promotes lipid metabolism [
48], while the family
Muribaculaceae has been demonstrated to exhibit probiotic effects and is associated with the innate immune system and the absorption and utilization of fat [
49]. A previous report indicated that the relative abundance of
Muribaculaceae is negatively correlated with fat deposition and is affected by the digestion of carbohydrates [
50]. In the present study, we observed an increase in the relative abundance of
Rikenellaceae_RC9_gut_group and
norank_
f_Muribaculaceae with DFPE supplementation. Meanwhile, these bacterium abundances were positively correlated with butyric acid, isovaleric acid, and valeric acid. Similarly, a previous study showed that feeding Hu sheep with the polyphenol-rich fruit
Nitraria tangutorum could increase the abundance of
Rikenellaceae_RC9_gut_group and
Christensenellaceae_R-7_group [
51]. In this study, a positive correlation was identified between
Rikenellaceae_RC9_gut_group and butyric acid content. Loubet Filho et al. [
52] observed that cyanidin-3-glucoside led to an augmentation in the abundance of
Muribaculaceae, suggesting that the bacteria of the
Muribaculaceae family may be the target of cyanidin-3-glucoside, potentially contributing to the positive effects of cyanidin-3-glucoside on metabolism and inflammation.
Firmicutes exhibit the capability to break down fibers into short-chain fatty acids. Within the Firmicutes phylum,
Christensenellaceae_R-7_group is commonly found in the intestinal tract and mucosa of both humans and animals, and it plays a crucial role in preserving the health of the host [
53]. Previous studies indicated that
Christensenellaceae_R-7_group has the potential to stimulate the development of the rumen, improving the absorption and digestion of nutrients. They have also exhibited a positive correlation with the breakdown of dietary protein, a negative correlation with diseases, such as inflammation [
54,
55,
56], and a positive correlation with nipple width, epithelial thickness, and stratum corneum thickness [
57]. Certainly, it is worth noting that microorganisms with different taxonomic features may serve the same function. Conversely, microorganisms with the same taxonomic characteristics may fulfill distinct functions. For example,
NK4A214_group (
Oscillospiraceae family) and
Christensenellaceae_R-7_group (
Christensenellaceae family), despite not belonging to the same family, may contribute individually or cooperatively to the promotion of ruminal biohydrogenation [
58].
Lachnospiraceae_NK3A20_group (
Lachnospiraceae family) plays an important role in rumen development and the promotion of rumen fermentation by increasing butyrate production [
57].
Oribacterium belongs to the
Lachnospiraceae family and has been recognized as one of the predominant bacteria in the rumen of cows that are fed forage-based diets [
59], but its role in the rumen has not been reported yet. In the present study, we found that polyphenols from DFPE increased the abundance of
Lachnospiraceae_NK3A20_group,
Christensenellaceae_R-7_group, and
NK4A214_group, indicating that polyphenols from DFPE can regulate rumen fermentation and potentially promote ruminal epithelial development, which may explain the rise in butyrate. A similar result was found in a report by Xu et al. [
60]. Polyphenols could potentially play a crucial role in safeguarding microorganisms from oxidative stress [
16]. This seems to explain the increased abundance of multiple bacteria caused by DFPE.
Streptococcus serves as the primary lactic acid fermenter in the rumen, converting starch and lactic acid into acetic acid and propionic acid. Since lactic acid is a stronger acid than VFA, the accumulation of lactate frequently causes a decrease in ruminal pH (5.0 or less) [
61].
Streptococcus displays a notable tolerance to acidity, enabling them to thrive and multiply at low pH levels [
62]. This characteristic may hinder the growth of other bacteria during such conditions [
62]. On the other hand, the
Streptococcus genus is recognized as one of the primary pathogens responsible for inducing mastitis [
63]. In the present study, we reveal that DFPE had the ability to reduce the relative abundance of
Streptococcus. The reason may be that polyphenols have the capability to engage with bacterial proteins, resulting in the inhibition of bacterial nucleic acid synthesis, the modulation of cell membrane function and fluidity, adjustments to cell wall integrity and synthesis, an impact on cell metabolism, and the prevention of biofilm formation [
64]; in addition, polyphenols have the ability to increase the production of short-chain fatty acids and inhibit the growth of pathogenic bacteria [
65]. Similarly, we also found
Streptococcus had a negative correlation with the butyric acid content.
Enterococcus is considered a probiotic, which contributes to the maintenance of lactic acid-utilizing bacteria activity, stimulates the growth of rumen microorganisms, and enhances the supply of gluco-propionic acid-producing energy for the host ruminants [
66]. In the present study, we found that
Enterococcus was decreased by DFPE. The mechanism for this is still unclear and requires future studies. A potential reason might be the increased relative abundance of certain microbes indirectly influencing the growth of other microbes [
16].










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}