

Fish Meal Substitution Effects with the Combined Animal Proteins in the Feeds of Olive Flounder (Paralichthys olivaceus) on Growth Performance, Feed Availability, and Disease Resistance against Streptococcus iniae

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of the Experimental Feeds
2.2. Rearing Conditions
2.3. Measurements of the Biological Indices of Fish
2.4. Plasma Analysis of Fish
2.5. Serum Analysis of Fish
2.6. Biochemical Composition of the Samples
2.7. Challenge Test of Fish
2.8. Statistical Analysis
3. Results
3.1. AA and FA Profiles of the Experimental Feeds
3.2. Survival, Weight Gain and SGR of Olive Flounder
3.3. Feed Availability and Biological Indices of Fish
3.4. Plasma Parameters of Fish
3.5. Serum Lysozyme Activity and SOD of Fish
3.6. Biochemical Composition of the Whole-Body Fish
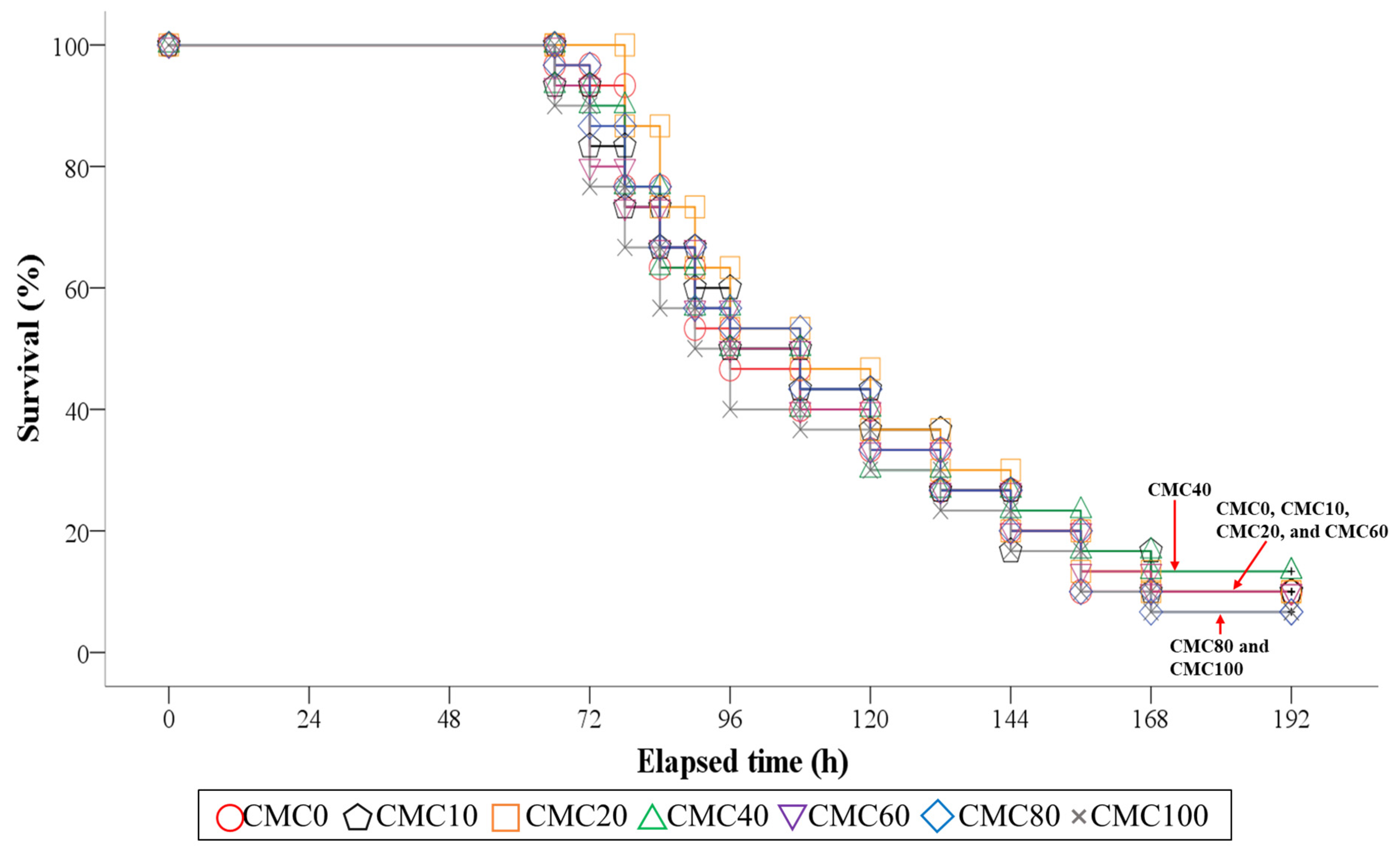
3.7. Survival of Fish in the 8-Day Post-Observation after S. iniae Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stieglitz, J.D.; Hoenig, R.H.; Baggett, J.K.; Tudela, C.E.; Mathur, S.K.; Benetti, D.D. Advancing production of marine fish in the United States: Olive flounder, Paralichthys olivaceus, aquaculture. J. World Aquacult. Soc. 2021, 52, 566–581. [Google Scholar] [CrossRef]
- Hamidoghli, A.; Won, S.; Lee, S.; Lee, S.; Farris, N.W.; Bai, S. Nutrition and feeding of olive flounder Paralichthys olivaceus: A Review. Rev. Fish. Sci. Aquac. 2020, 28, 340–357. [Google Scholar] [CrossRef]
- KOSIS. Korean Statistical Information Service. 2023. Available online: https://kosis.kr/statHtml/statHtml.do?orgId=101&tblId=DT_1EZ0008&conn_path=I2 (accessed on 15 June 2023).
- FAO. Fisheries & Aquaculture-Global Aquaculture Production Quantity. 2023. Available online: https://www.fao.org/fishery/statistics-query/en/aquaculture/aquaculture_quantity (accessed on 16 June 2023).
- Gasco, L.; Gai, F.; Maricchiolo, G.; Genovese, L.; Ragonese, S.; Bottari, T.; Caruso, G. Fishmeal alternative protein sources for aquaculture feeds. In Feeds for the Aquaculture Sector: Current Situation and Alternative Sources; Gasco, L., Gai, F., Maricchiolo, G., Genovese, L., Ragonese, S., Bottari, T., Caruso, G., Eds.; Springer International Publishing: Berlin, Germany, 2018; pp. 1–28. [Google Scholar] [CrossRef]
- Galkanda-Arachchige, H.S.; Wilson, A.E.; Davis, D.A. Success of fishmeal replacement through poultry by-product meal in aquaculture feed formulations: A meta-analysis. Rev. Aquac. 2020, 12, 1624–1636. [Google Scholar] [CrossRef]
- Tacon, A.G.; Metian, M. Feed matters: Satisfying the feed demand of aquaculture. Rev. Fish. Sci. Aquac. 2015, 23, 1–10. [Google Scholar] [CrossRef]
- Oncul, F.O.; Aya, F.A.; Hamidoghli, A.; Won, S.; Lee, G.; Han, K.R.; Bai, S. Effects of the dietary fermented tuna by-product meal on growth, blood parameters, nonspecific immune response, and disease resistance in juvenile olive flounder, Paralichthys olivaceus. J. World Aquacult. Soc. 2019, 50, 65–77. [Google Scholar] [CrossRef]
- Seong, M.; Lee, S.; Lee, S.; Song, Y.; Bae, J.; Chang, K.; Bai, S.C. The effects of different levels of dietary fermented plant-based protein concentrate on growth, hematology and non-specific immune responses in juvenile olive flounder, Paralichthys olivaceus. Aquaculture 2018, 483, 196–202. [Google Scholar] [CrossRef]
- Kim, K.W.; Wang, X.J.; Bai, S.C. Optimum dietary protein level for maximum growth of juvenile olive flounder Paralichthys olivaceus (Temminck et Schlegel). Aquac. Res. 2002, 33, 673–679. [Google Scholar] [CrossRef]
- Lee, S.; Cho, S.H.; Kim, K. Effects of dietary protein and energy levels on growth and body composition of juvenile flounder Paralichthys olivaceus. J. World Aquacult. Soc. 2000, 31, 306–315. [Google Scholar] [CrossRef]
- Hua, K.; Cobcroft, J.M.; Cole, A.; Condon, K.; Jerry, D.R.; Mangott, A.; Praeger, C.; Vucko, M.J.; Zeng, C.; Zenger, K.; et al. The future of aquatic protein: Implications for protein sources in aquaculture diets. One Earth 2019, 1, 316–329. [Google Scholar] [CrossRef]
- Carvalho, M.; Torrecillas, S.; Montero, D.; Sanmartín, A.; Fontanillas, R.; Farías, A.; Moutou, K.; Velasquez, J.H.; Izquierdo, M. Insect and single-cell protein meals as replacers of fish meal in low fish meal and fish oil diets for gilthead sea bream (Sparus aurata) juveniles. Aquaculture 2023, 566, 739215. [Google Scholar] [CrossRef]
- Zaman, M.F.U.; Li, R.; Cho, S.H. Evaluation of meat meal as a replacer for fish meal in diet on growth performance, feed utilization, chemical composition, hematology, and innate immune responses of olive flounder (Paralichthys olivaceus). Fishes 2022, 7, 343. [Google Scholar] [CrossRef]
- Ye, H.; Zhou, Y.; Su, N.; Wang, A.; Tan, X.; Sun, Z.; Zou, C.; Liu, Q.; Ye, C. Effects of replacing fish meal with rendered animal protein blend on growth performance, hepatic steatosis and immune status in hybrid grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂). Aquaculture 2019, 511, 734203. [Google Scholar] [CrossRef]
- Lee, M.J.; Kim, J.; Baek, S.I.; Cho, S.H. Substitution effect of fish meal with meat meal in diet on growth performance, feed consumption, feed utilization, chemical composition, hematology, and innate immune responses of rockfish (Sebastes schlegeli). Aquaculture 2023, 571, 739467. [Google Scholar] [CrossRef]
- Dawson, M.R.; Alam, M.S.; Watanabe, W.O.; Carroll, P.M.; Seaton, P.J. Evaluation of poultry by-product meal as an alternative to fish meal in the diet of juvenile Black sea bass reared in a recirculating aquaculture system. N. Am. J. Aquacult. 2018, 80, 74–87. [Google Scholar] [CrossRef]
- Woodgate, S.; Van Der Veen, J. The role of fat processing and rendering in the European Union animal production industry. Biotechnol. Agron. Soc. Environ. 2004, 8, 283–294. [Google Scholar]
- Toldrá, T.; Mora, L.; Reig, M. Current developments in meat by-products. In New Aspects of Meat Quality; Purslow, P.P., Ed.; Genes to Ethics: Cambridgeshire, UK, 2022; pp. 649–665. [Google Scholar] [CrossRef]
- Hicks, T.M.; Verbeek, C.J.R. Meat industry protein by-products: Sources and characteristics. In Protein Byproducts: Transformation from Environmental Burden into Value-Added Products; Dhillon, G.S., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 37–61. [Google Scholar] [CrossRef]
- Lee, S. Apparent digestibility coefficients of various feed ingredients for juvenile and grower rockfish (Sebastes schlegeli). Aquaculture 2002, 207, 79–95. [Google Scholar] [CrossRef]
- Williams, K.C.; Barlow, C.G.; Rodgers, L.J.; Ruscoe, I. Potential of meat meal to replace fish meal in extruded dry diets for barramundi, Lates calcarifer (Bloch). I. Growth performance. Aquac. Res. 2003, 34, 23–32. [Google Scholar] [CrossRef]
- Ha, M.S.; Cho, S.H.; Kim, T. Dietary substitution of fish meal by meat meal: Effects on juvenile olive flounder (Paralichthys olivaceus) growth performance, feed utilization, haematology, biochemical profile and disease resistance against Streptococcus iniae. Aquac. Nutr. 2021, 27, 1888–1902. [Google Scholar] [CrossRef]
- Sato, T.; Kikuchi, K. Meat meal as a protein source in the diet of juvenile Japanese flounder. Fish. Sci. 1997, 63, 877–880. [Google Scholar] [CrossRef]
- Karapanagiotidis, I.T.; Psofakis, P.; Mente, E.; Malandrakis, E.; Golomazou, E. Effect of fishmeal replacement by poultry by-product meal on growth performance, proximate composition, digestive enzyme activity, haematological parameters and gene expression of gilthead seabream (Sparus aurata). Aquac. Nutr. 2019, 25, 3–14. [Google Scholar] [CrossRef]
- Cruz-Suárez, L.E.; Nieto-López, M.; Guajardo-Barbosa, C.; Tapia-Salazar, M.; Scholz, U.; Ricque-Marie, D. Replacement of fish meal with poultry by-product meal in practical diets for Litopenaeus vannamei, and digestibility of the tested ingredients and diets. Aquaculture 2007, 272, 466–476. [Google Scholar] [CrossRef]
- LPSMS. Livestock Product Safety Management System (LPSMS). Annual Slaughter Report in 2022. 2023. Available online: https://www.lpsms.go.kr/home/stats/stats.do?statsFlag=statsDomesticFowls (accessed on 27 June 2023).
- Ha, M.S.; Lee, K.W.; Kim, J.; Yun, A.; Jeong, H.S.; Lee, M.J.; Baek, S.I.; Cho, S.H.; Kim, K.W.; Lim, S.G.; et al. Dietary substitution effect of fish meal with chicken by-product meal on growth, feed utilization, body composition, haematology and non-specific immune responses of olive flounder (Paralichthys olivaceus). Aquac. Nutr. 2021, 27, 315–326. [Google Scholar] [CrossRef]
- Nandakumar, S.; Ambasankar, K.; Dayal, J.S.; Raman, C.; Ali, S.R. Fish meal replacement with chicken waste meal in Asian seabass (Lates calcarifer) feeds. Indian J. Fish. 2013, 60, 109–114. [Google Scholar]
- Guo, J.; Wang, Y.; Bureau, D.P. Inclusion of rendered animal ingredients as fishmeal substitutes in practical diets for cuneate drum, Nibea miichthioides (Chu, Lo et Wu). Aquac. Nutr. 2007, 13, 81–87. [Google Scholar] [CrossRef]
- Sun, C.X.; Xu, W.N.; Li, X.F.; Zhang, D.D.; Qian, Y.; Jiang, G.Z.; Liu, W.B. Effects of fish meal replacement with animal protein blend on growth performance, nutrient digestibility and body composition of juvenile Chinese soft-shelled turtle Pelodiscus sinensis. Aquac. Nutr. 2016, 22, 315–325. [Google Scholar] [CrossRef]
- Gunathilaka, B.E.; Jeong, S.; Cho, M.; Kim, K.; Hur, S.; Lee, S.; You, S.; Lee, S. Effects of dietary fish meal replacement with alternative protein ingredients and their combinations on growth performance, feed utilization, fillet composition, and biochemical parameters of red seabream (Pagrus major). Aquac. Nutr. 2023, 2023, 8883739. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, K.; Han, H.; Zheng, Z.; Bureau, D.P. Potential of using a blend of rendered animal protein ingredients to replace fish meal in practical diets for malabar grouper (Epinephelus malabricus). Aquaculture 2008, 281, 113–117. [Google Scholar] [CrossRef]
- Bu, X.; Chen, A.; Lian, X.; Chen, F.; Zhang, Y.; Muhammad, I.; Ge, X.; Yang, Y. An evaluation of replacing fish meal with cottonseed meal in the diet of juvenile Ussuri catfish Pseudobagrus ussuriensis: Growth, antioxidant capacity, nonspecific immunity and resistance to Aeromonas hydrophila. Aquaculture 2017, 479, 829–837. [Google Scholar] [CrossRef]
- Yang, H.; Bian, Y.; Huang, L.; Lan, Q.; Ma, L.; Li, X.; Leng, X. Effects of replacing fish meal with fermented soybean meal on the growth performance, intestinal microbiota, morphology and disease resistance of largemouth bass (Micropterus salmoides). Aquac. Rep. 2022, 22, 100954. [Google Scholar] [CrossRef]
- Zhang, X.; Sun, Z.; Cai, J.; Wang, J.; Wang, G.; Zhu, Z.; Cao, F. Effects of dietary fish meal replacement by fermented moringa (Moringa oleifera Lam.) leaves on growth performance, nonspecific immunity and disease resistance against Aeromonas hydrophila in juvenile gibel carp (Carassius auratus gibelio var. CAS III). Fish Shellfish Immunol. 2020, 102, 430–439. [Google Scholar] [CrossRef]
- Kim, M.; Jin, J.W.; Han, H.J.; Choi, H.S.; Hong, S.; Cho, J.Y. Genotype and virulence of Streptococcus iniae isolated from diseased olive flounder Paralichthys olivaceus in Korea. Fish. Sci. 2014, 80, 1277–1284. [Google Scholar] [CrossRef]
- Shin, G.W.; Palaksha, K.J.; Yang, H.H.; Shin, Y.S.; Kim, Y.R.; Lee, E.Y.; Kim, H.Y.; Kim, Y.J.; Oh, M.J.; Yoshida, T.; et al. Discrimination of streptococcosis agents in olive flounder (Paralichthys olivaceus). Bull. Eur. Ass. Fish Pathol. 2006, 26, 68. [Google Scholar]
- Kim, J.; Sohn, S.; Kim, S.K.; Kim, S.; Kim, S.; Kim, S.M.; Kim, N.Y.; Hur, Y.B. Effects on the survival rates, hematological parameters, and neurotransmitters in olive flounders, Paralichthys olivaceus, reared in bio-floc and seawater by Streptococcus iniae challenge. Fish Shellfish Immunol. 2021, 113, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Lange, S.; Gudmundsdottir, B.K.; Magnadottir, B. Humoral immune parameters of cultured Atlantic halibut (Hippoglossus hippoglossus L.). Fish Shellfish Immunol. 2001, 11, 523–535. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Cho, S.H.; Kim, T. Effect of replacing dietary fish meal protein with combined animal meals on the growth performance of olive flounder (Paralichthys olivaceus). Aquac. Rep. 2023, 32, 101712. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemists Inc.: Rockville, MD, USA, 1990. [Google Scholar]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.S.; Kim, J.; Olowe, O.S.; Cho, S.H. Dietary optimum inclusion level of jack mackerel meal for olive flounder (Paralichthys olivaceus, Temminck & Schlegel, 1846). Aquaculture 2022, 559, 738432. [Google Scholar] [CrossRef]
- Alam, M.S.; Teshima, S.; Koshio, S.; Ishikawa, M. Arginine requirement of juvenile Japanese flounder Paralichthys olivaceus estimated by growth and biochemical parameters. Aquaculture 2002, 205, 127–140. [Google Scholar] [CrossRef]
- Forster, I.; Ogata, H.Y. Lysine requirement of juvenile Japanese flounder Paralichthys olivaceus and juvenile red sea bream Pagrus major. Aquaculture 1998, 161, 131–142. [Google Scholar] [CrossRef]
- Hasanthi, M.; Kim, M.; Lim, H.; Lim, J.; Hur, S.; Lee, S.; Lee, B.; Kim, K.; Lee, K. The dietary requirement for threonine in juvenile olive flounder (Paralichthys olivaceus). Fish. Aquat. Sci. 2023, 26, 58–68. [Google Scholar] [CrossRef]
- Alam, M.S.; Teshima, S.; Ishikawa, M.; Koshio, S. Methionine requirement of juvenile Japanese flounder Paralichthys olivaceus. J. World Aquacult. Soc. 2000, 31, 618–626. [Google Scholar] [CrossRef]
- Kim, K.; Lee, S. Requirement of dietary n-3 highly unsaturated fatty acids for juvenile flounder (Paralichthys olivaceus). Aquaculture 2004, 229, 315–323. [Google Scholar] [CrossRef]
- Farhat Khan, M.A. Total sulfur amino acid requirement and cystine replacement value for fingerling stinging catfish, Heteropneustes fossilis (Bloch). Aquaculture 2014, 426, 270–281. [Google Scholar] [CrossRef]
- Zehra, S.; Khan, M.A. Total sulphur amino acid requirement and maximum cysteine replacement value for methionine for fingerling Catla catla (Hamilton). Aquac. Res. 2016, 47, 304–317. [Google Scholar] [CrossRef]
- Socas-Rodríguez, B.; Álvarez-Rivera, G.; Valdés, A.; Ibáñez, E.; Cifuentes, A. Food by-products and food wastes: Are they safe enough for their valorization? Trends Food Sci. Technol. 2021, 114, 133–147. [Google Scholar] [CrossRef]
- Volpato, J.A.; Ribeiro, L.B.; Torezan, G.B.; da Silva, I.C.; de Oliveira Martins, I.; Genova, J.L.; de Oliveira, N.T.E.; Carvalho, S.T.; de Oliveira Carvalho, P.L.; Vasconcellos, R.S. Characterization of the variations in the industrial processing and nutritional variables of poultry by-product meal. Poult. Sci. 2022, 101, 101926. [Google Scholar] [CrossRef] [PubMed]
- Tocher, D.R. Fatty acid requirements in ontogeny of marine and freshwater fish. Aquac. Res. 2010, 41, 717–732. [Google Scholar] [CrossRef]
- Pike, I.H.; Jackson, A. Fish oil: Production and use now and in the future. Lipid Technol. 2010, 22, 59–61. [Google Scholar] [CrossRef]
- Tocher, D.R.; Betancor, M.B.; Sprague, M.; Olsen, R.E.; Napier, J.A. Omega-3 long-chain polyunsaturated fatty acids, EPA and DHA: Bridging the gap between supply and demand. Nutrients 2019, 11, 89. [Google Scholar] [CrossRef]
- Kim, J.; Cho, S.H.; Kim, T.; Hur, S.W. Substitution effect of fish meal with various sources of animal by-product meals in feed on growth, feed utilization, body composition, haematology and non-specific immune response of olive flounder (Paralichthys olivaceus, Temminck & Schlegel, 1846). Aquac. Res. 2021, 52, 2802–2817. [Google Scholar] [CrossRef]
- Hara, T.J. Feeding behaviour in some teleosts is triggered by single amino acids primarily through olfaction. J. Fish Biol. 2006, 68, 810–825. [Google Scholar] [CrossRef]
- Polat, A.; Beklevik, G. The importance of betaine and some attractive substances as fish feed additives. In Feed Manufacturing in the Mediterranean Region: Recent Advances in Research and Technology; Brufau, J., Tacon, A., Eds.; CIHEAM: Zaragoza, Spain, 1999; pp. 217–220. [Google Scholar]
- Chen, Q.; Zhao, H.; Huang, Y.; Cao, J.; Wang, G.; Sun, Y.; Li, Y. Effects of dietary arginine levels on growth performance, body composition, serum biochemical indices and resistance ability against ammonia-nitrogen stress in juvenile yellow catfish (Pelteobagrus fulvidraco). Anim. Nutr. 2016, 2, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Tan, X.; Zhou, C.; Niu, J.; Xia, D.; Huang, Z.; Wang, J.; Wang, Y. Effect of dietary arginine levels on the growth performance, feed utilization, non-specific immune response and disease resistance of juvenile golden pompano Trachinotus ovatus. Aquaculture 2015, 437, 382–389. [Google Scholar] [CrossRef]
- Kader, M.A.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Bulbul, M. Supplemental effects of some crude ingredients in improving nutritive values of low fishmeal diets for red sea bream, Pagrus major. Aquaculture 2010, 308, 136–144. [Google Scholar] [CrossRef]
- Tharaka, K.; Benitez-Santana, T.; Gunathilaka, B.E.; Kim, M.; Lee, C.; Shin, J.; Lee, K. Evaluation of Antarctic krill (Euphausia superba) meal supplementation in diets for olive flounder (Paralichthys olivaceus). Aquac. Res. 2020, 51, 2291–2302. [Google Scholar] [CrossRef]
- Jeong, H.S.; Choi, D.G.; Lee, K.W.; Cho, S.H.; Lim, S.G.; Lee, B.J.; Hur, S.W.; Son, M.H.; Lee, S.H.; Kim, K.W. Attractiveness of various crude feed ingredients to juvenile olive flounder (Paralichthys olivaceus, Temminck & Schlegel) and its application to aquaculture. Aquac. Res. 2020, 51, 4517–4532. [Google Scholar] [CrossRef]
- Baek, S.I.; Jeong, H.S.; Cho, S.H. Replacement effect of fish meal by plant protein sources in olive flounder (Paralichthys olivaceus) feeds with an addition of jack mackerel meal on growth, feed availability, and biochemical composition. Aquac. Nutr. 2023, 2023, 7965258. [Google Scholar] [CrossRef]
- Jeong, H.S.; Cho, S.H. Inclusion effect of jack mackerel meal as feed stimulants in diets replacing different levels of fish meal with various animal protein sources on growth performance of olive flounder (Paralichthys olivaceus). Aquac. Rep. 2023, 28, 101450. [Google Scholar] [CrossRef]
- Hancz, C. Feed efficiency, nutrient sensing and feeding stimulation in aquaculture: A review. Acta Agr. Kapos. 2020, 24, 35–54. [Google Scholar] [CrossRef]
- Ariweriokuma, S.V.; Akinrotimi, O.A.; Gabriel, U.U. Effects of cypermethrin on condition factor and organosomatic indices of Clarias gariepinus. J. Agric. Soc. Res. 2011, 11, 67–72. [Google Scholar]
- Wu, Y.B.; Ren, X.; Chai, X.J.; Li, P.; Wang, Y. Replacing fish meal with a blend of poultry by-product meal and feather meal in diets for giant croaker (Nibea japonica). Aquac. Nutr. 2017, 24, 1085–1091. [Google Scholar] [CrossRef]
- Peres, H.; Santos, S.; Oliva-Teles, A. Selected plasma biochemistry parameters in gilthead seabream (Sparus aurata) juveniles. J. Appl. Ichthyol. 2013, 29, 630–636. [Google Scholar] [CrossRef]
- Kiron, V. Fish immune system and its nutritional modulation for preventive health care. Anim. Feed Sci. Technol. 2012, 173, 111–133. [Google Scholar] [CrossRef]
- Yin, C.; Li, N.; Ren, L.; Wang, Z.; Chai, L.; Lan, J. Identification and characterization of two highly homologous lysozymes from red swamp crayfish, Procambarus clarkii. Fish. Shellfish Immunol. Rep. 2021, 2, 100017. [Google Scholar] [CrossRef]
- Ribeiro, F.; Garcia, A.R.; Pereira, B.P.; Fonseca, M.; Mestre, N.C.; Fonseca, T.G.; Mestre, N.C.; Fonseca, T.G.; Ilharco, L.M.; Bebianno, M.J. Microplastics effects in Scrobicularia plana. Mar. Pollut. Bull. 2017, 122, 379–391. [Google Scholar] [CrossRef]
- Yamamoto, T.; Unuma, T.; Akiyama, T. The influence of dietary protein and fat levels on tissue free amino acid levels of fingerling rainbow trout (Oncorhynchus mykiss). Aquaculture 2000, 182, 353–372. [Google Scholar] [CrossRef]
- Nho, S.; Shin, G.; Park, S.; Jang, H.; Cha, I.; Ha, M.; Kim, Y.; Park, Y.; Dalvi, R.; Kang, B.; et al. Phenotypic characteristics of Streptococcus iniae and Streptococcus parauberis isolated from olive flounder (Paralichthys olivaceus). FEMS Microbiol. Lett. 2009, 293, 20–27. [Google Scholar] [CrossRef]
- Pham, H.D.; Siddik, M.A.B.; Le, H.M.; Ngo, M.V.; Nguyen, M.V.; Francis, D. Effects of dietary tuna viscera hydrolysate supplementation on growth, intestinal mucosal response, and resistance to Streptococcus iniae infection in pompano (Trachinotus blochii). Aquac. Nutr. 2022, 2022, 3645868. [Google Scholar] [CrossRef]
- Sealey, W.M.; Hardy, R.W.; Barrows, F.T.; Pan, Q.; Stone, D.A. Evaluation of 100% fish meal substitution with chicken concentrate, protein poultry by-product blend, and chicken and egg concentrate on growth and disease resistance of juvenile rainbow trout, Oncorhynchus mykiss. J. World Aquacult. Soc. 2011, 42, 46–55. [Google Scholar] [CrossRef]


{kind=link}
| Experimental Diets | |||||||
|---|---|---|---|---|---|---|---|
| CMC0 | CMC10 | CMC20 | CMC40 | CMC60 | CMC80 | CMC100 | |
| Ingredient (%, DM) | |||||||
| Fish meal 1 | 65.0 | 58.5 | 52.0 | 39.0 | 26.0 | 13.0 | 0.0 |
| Combined meat meal and chicken by-product meal (CMC) 2 | 0.0 | 6.1 | 12.2 | 25.0 | 36.7 | 49.0 | 61.2 |
| Dehulled soybean meal | 12.0 | 12.0 | 12.0 | 12.0 | 12.0 | 12.0 | 12.0 |
| Wheat flour | 14.9 | 15.7 | 16.4 | 18.3 | 19.3 | 20.7 | 22.3 |
| Krill meal | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 |
| Fish oil | 2.3 | 2.3 | 2.3 | 2.3 | 2.3 | 1.8 | 1.0 |
| Soybean oil | 2.3 | 1.9 | 1.6 | 0.8 | 0.2 | 0.0 | 0.0 |
| Vitamin premix 3 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 |
| Mineral premix 4 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 | 1.0 |
| Choline | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| Nutrients (%, DM) | |||||||
| Dry matter | 97.0 ± 0.27 | 96.8 ± 0.03 | 96.4 ± 0.21 | 95.8 ± 0.13 | 96.1 ± 0.23 | 94.1 ± 0.90 | 94.2 ± 0.04 |
| Crude protein | 56.0 ± 0.37 | 55.9 ± 0.78 | 56.6 ± 0.09 | 56.4 ± 0.35 | 56.6 ± 0.06 | 57.0 ± 0.40 | 57.0 ± 0.72 |
| Crude lipid | 10.8 ± 0.01 | 10.6 ± 0.34 | 10.8 ± 0.28 | 10.5 ± 0.08 | 10.6 ± 0.03 | 10.8 ± 0.31 | 10.8 ± 0.39 |
| Ash | 14.4 ± 0.00 | 13.9 ± 0.10 | 12.9 ± 0.19 | 11.0 ± 0.15 | 10.8 ± 0.11 | 9.0 ± 0.19 | 7.8 ± 0.39 |
| Ingredients | Requirement | Experimental Diets | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FM | MM | CBM | CMC | CMC0 | CMC10 | CMC20 | CMC40 | CMC60 | CMC80 | CMC100 | ||
| Essential amino acid (EAA) (%) | ||||||||||||
| Arginine | 3.78 | 5.32 | 4.68 | 5.00 | 2.04–2.10 1 | 3.03 | 3.14 | 3.43 | 3.74 | 3.82 | 3.92 | 3.99 |
| Histidine | 2.05 | 1.17 | 1.47 | 1.32 | 1.28 | 1.27 | 1.25 | 1.23 | 1.22 | 1.21 | 1.20 | |
| Isoleucine | 2.83 | 1.66 | 2.81 | 2.24 | 1.95 | 1.93 | 1.92 | 1.89 | 1.88 | 1.86 | 1.85 | |
| Leucine | 4.93 | 3.48 | 4.82 | 4.15 | 4.40 | 4.23 | 4.17 | 4.10 | 4.08 | 3.91 | 3.69 | |
| Lysine | 5.14 | 3.56 | 4.43 | 4.00 | 1.50–2.10 2 | 3.91 | 3.74 | 3.47 | 3.44 | 3.40 | 3.31 | 3.22 |
| Methionine | 1.84 | 0.86 | 1.45 | 1.16 | 1.44–1.49 3 | 1.14 | 0.99 | 0.94 | 0.84 | 0.79 | 0.64 | 0.53 |
| Phenylalanine | 2.71 | 2.08 | 2.68 | 2.38 | 1.90 | 1.88 | 1.86 | 1.84 | 1.80 | 1.77 | 1.74 | |
| Threonine | 2.87 | 2.00 | 2.76 | 2.38 | 1.03 4 | 2.26 | 2.24 | 2.17 | 2.13 | 2.09 | 2.03 | 1.95 |
| Tryptophan | 0.71 | 0.53 | 0.50 | 0.52 | 0.54 | 0.48 | 0.47 | 0.46 | 0.43 | 0.40 | 0.33 | |
| Valine | 3.44 | 2.61 | 3.33 | 2.97 | 2.14 | 2.10 | 2.07 | 2.03 | 2.00 | 1.98 | 1.93 | |
| ∑EAA 5 | 30.30 | 23.27 | 28.93 | 26.10 | 22.55 | 22.00 | 21.75 | 21.70 | 21.51 | 21.03 | 20.43 | |
| Non-essential amino acid (NEAA) (%) | ||||||||||||
| Alanine | 4.18 | 5.82 | 4.47 | 5.15 | 3.09 | 3.17 | 3.22 | 3.28 | 3.33 | 3.37 | 3.53 | |
| Aspartic acid | 6.03 | 5.04 | 5.86 | 5.45 | 4.88 | 4.83 | 4.67 | 4.66 | 4.60 | 4.54 | 4.49 | |
| Cysteine | 0.63 | 0.37 | 0.45 | 0.41 | 0.63 | 0.57 | 0.55 | 0.45 | 0.42 | 0.40 | 0.37 | |
| Glutamic acid | 8.49 | 8.16 | 9.07 | 8.62 | 6.85 | 6.90 | 6.91 | 6.95 | 6.99 | 7.03 | 7.04 | |
| Glycine | 4.19 | 12.26 | 5.82 | 9.04 | 3.08 | 3.44 | 3.91 | 4.49 | 5.17 | 5.71 | 6.45 | |
| Proline | 2.59 | 7.34 | 3.82 | 5.58 | 3.83 | 3.95 | 4.02 | 4.18 | 4.46 | 5.25 | 6.04 | |
| Serine | 2.57 | 2.59 | 2.52 | 2.56 | 1.96 | 1.95 | 1.93 | 1.92 | 1.90 | 1.87 | 1.86 | |
| Tyrosine | 1.61 | 1.23 | 1.97 | 1.60 | 1.86 | 1.85 | 1.83 | 1.80 | 1.78 | 1.77 | 1.74 | |
| ∑NEAA 6 | 30.29 | 42.81 | 33.98 | 38.40 | 26.18 | 26.66 | 27.04 | 27.73 | 28.65 | 29.94 | 31.52 | |
| Ingredients | Requirement | Experimental Diets | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FM | MM | CBM | CMC | CMC0 | CMC10 | CMC20 | CMC40 | CMC60 | CMC80 | CMC100 | ||
| C14:0 | 4.16 | 2.60 | 1.41 | 2.01 | 3.00 | 2.98 | 2.82 | 2.56 | 2.19 | 2.13 | 2.05 | |
| C16:0 | 27.11 | 29.89 | 35.89 | 32.89 | 15.87 | 17.39 | 17.84 | 18.97 | 19.28 | 20.23 | 23.15 | |
| C18:0 | 9.44 | 12.27 | 13.57 | 12.92 | 5.70 | 6.43 | 6.46 | 7.14 | 8.66 | 10.13 | 13.29 | |
| ∑SFA 1 | 40.71 | 44.76 | 50.87 | 47.82 | 24.57 | 26.80 | 27.12 | 28.67 | 30.13 | 32.49 | 38.49 | |
| C16:1n−7 | 9.26 | 7.88 | 5.22 | 6.55 | 3.41 | 3.38 | 3.30 | 3.11 | 2.96 | 2.56 | 2.42 | |
| C17:1n−7 | 0.59 | 0.47 | 0.29 | 0.38 | 0.52 | 0.50 | 0.49 | 0.45 | 0.42 | 0.38 | 0.34 | |
| C18:1n−9 | 17.33 | 34.37 | 37.59 | 35.98 | 19.86 | 22.51 | 24.73 | 29.60 | 31.23 | 35.27 | 38.49 | |
| C20:1n−9 | 1.12 | 0.62 | 0.22 | 0.42 | 1.32 | 0.96 | 0.92 | 0.90 | 0.85 | 0.81 | 0.57 | |
| ∑MUFA 2 | 28.30 | 43.34 | 43.32 | 43.33 | 25.11 | 27.35 | 29.44 | 34.06 | 35.46 | 39.02 | 41.82 | |
| C18:2n−6 | 2.07 | 1.66 | 1.66 | 27.21 | 25.69 | 24.54 | 22.50 | 21.87 | 18.39 | 13.05 | ||
| C18:3n−3 | 0.73 | 0.59 | 0.19 | 0.39 | 3.97 | 3.59 | 3.35 | 2.89 | 2.72 | 1.97 | 1.31 | |
| C20:4n−6 | 1.53 | 0.73 | 0.59 | 0.66 | 1.07 | 0.95 | 0.86 | 0.79 | 0.59 | 0.35 | 0.21 | |
| C20:5n−3 | 4.89 | 5.02 | 4.41 | 4.12 | 3.46 | 2.89 | 2.03 | 1.87 | ||||
| C22:5n−3 | 0.90 | 0.33 | 0.29 | 0.31 | 0.98 | 0.84 | 0.73 | 0.64 | 0.49 | 0.37 | 0.30 | |
| C22:6n−3 | 13.11 | 8.44 | 6.94 | 6.71 | 5.41 | 4.61 | 2.50 | 1.09 | ||||
| ∑n−3 HUFA 3 | 18.90 | 0.33 | 0.29 | 0.31 | 7.41–9.26 4 | 14.44 | 12.19 | 11.56 | 9.51 | 7.99 | 4.90 | 3.26 |
| Unknown | 7.76 | 8.59 | 4.74 | 5.84 | 3.63 | 3.43 | 3.13 | 1.58 | 1.24 | 2.88 | 1.86 | |
| Experimental Diets | Initial Weight (g/fish) | Final Weight (g/fish) | Survival (%) | Weight Gain (g/fish) | SGR (%/day) 1 |
|---|---|---|---|---|---|
| CMC0 | 9.2 ± 0.00 | 44.3 ± 0.96 c | 100 ± 0.00 | 35.1 ± 0.96 c | 2.81 ± 0.039 c |
| CMC10 | 9.2 ± 0.00 | 49.2 ± 0.60 b | 97.3 ± 2.31 | 40.0 ± 0.60 b | 2.99 ± 0.021 b |
| CMC20 | 9.2 ± 0.00 | 56.5 ± 2.58 a | 100 ± 0.00 | 47.3 ± 2.58 a | 3.24 ± 0.082 a |
| CMC40 | 9.2 ± 0.01 | 49.4 ± 1.18 b | 100 ± 0.00 | 40.2 ± 1.17 b | 3.00 ± 0.040 b |
| CMC60 | 9.2 ± 0.01 | 40.9 ± 1.63 c | 98.7 ± 2.31 | 31.7 ± 1.62 c | 2.66 ± 0.068 c |
| CMC80 | 9.2 ± 0.01 | 33.9 ± 0.63 d | 97.3 ± 2.31 | 24.7 ± 0.63 d | 2.33 ± 0.033 d |
| CMC100 | 9.2 ± 0.01 | 27.1 ± 1.31 e | 97.3 ± 2.31 | 17.8 ± 0.32 e | 1.92 ± 0.021 e |
| p–value | p < 0.0001 | p > 0.1 | p < 0.0001 | p < 0.0001 |
| Dependent Variables | Regression Analysis | |||
|---|---|---|---|---|
| Equation | p-Value | R2 | Ymax (%) | |
| Weight gain | Y = 0.000117X3 − 0.021899X2 + 0.852379X + 35.0989 | 0.0001 | 0.9560 | 24.1 |
| SGR | Y = 0.000003662X3 − 0.000756X2 + 0.030304X + 2.8084 | 0.0001 | 0.9711 | 24.4 |
| Feed consumption | Y = 0.000102X3 − 0.019279X2 + 0.748248X + 35.0246 | 0.0001 | 0.9608 | 24.0 |
| FE | Y = −0.0000002692X3 − 0.00004974X2 + 0.002006X + 1.0121 | 0.0001 | 0.7412 | 17.7 |
| PER | Y = 0.0000004873X3 − 0.000100X2 + 0.003680X + 1.7915 | 0.0001 | 0.8589 | 21.9 |
| Experimental Diets | Feed Consumption (g/fish) | FE 1 | PER 2 | PR 3 (%) | K 4 (g/cm3) | VSI 5 (%) | HIS 6 (%) |
|---|---|---|---|---|---|---|---|
| CMC0 | 33.75 ± 0.544 c | 1.04 ± 0.012 ab | 1.85 ± 0.021 a | 34.78 ± 0.632 | 1.08 ± 0.018 | 3.16 ± 0.004 | 1.23 ± 0.000 |
| CMC10 | 39.57 ± 0.542 b | 1.04 ± 0.022 ab | 1.81 ± 0.011 a | 33.73 ± 0.127 | 1.10 ± 0.003 | 3.16 ± 0.015 | 1.22 ± 0.000 |
| CMC20 | 45.16 ± 0.760 a | 1.05 ± 0.040 a | 1.85 ± 0.071 a | 34.31 ± 1.327 | 1.11 ± 0.007 | 3.16 ± 0.011 | 1.22 ± 0.003 |
| CMC40 | 39.43 ± 0.930 b | 1.02 ± 0.012 ab | 1.81 ± 0.022 a | 33.90 ± 0.400 | 1.11 ± 0.019 | 3.16 ± 0.001 | 1.23 ± 0.000 |
| CMC60 | 31.60 ± 1.080 cd | 1.02 ± 0.007 ab | 1.79 ± 0.034 ab | 34.03 ± 0.389 | 1.06 ± 0.055 | 3.16 ± 0.002 | 1.22 ± 0.001 |
| CMC80 | 25.27 ± 0.689 d | 1.00 ± 0.006 ab | 1.74 ± 0.029 ab | 33.71 ± 0.468 | 1.04 ± 0.057 | 3.16 ± 0.005 | 1.22 ± 0.005 |
| CMC100 | 18.99 ± 0.592 e | 0.99 ± 0.020 b | 1.69 ± 0.074 b | 33.79 ± 1.303 | 1.04 ± 0.063 | 3.16 ± 0.008 | 1.22 ± 0.006 |
| p–value | p < 0.0001 | p < 0.03 | p < 0.002 | p > 0.3 | p > 0.07 | p > 0.8 | p > 0.8 |
| Experimental Diets | AST (U/L) | ALT (U/L) | ALP (U/L) | TBL (mg/dL) | TCO (mg/dL) | TRG (mg/dL) | TPT (g/dL) | ALB (g/dL) |
|---|---|---|---|---|---|---|---|---|
| CMC0 | 13.0 ± 1.22 | 5.7 ± 0.71 | 92.3 ± 4.66 | 0.9 ± 0.15 | 179.4 ± 16.71 | 443.2 ± 14.86 | 3.7 ± 0.35 | 1.1 ± 0.15 |
| CMC10 | 13.0 ± 2.00 | 5.9 ± 1.05 | 92.4 ± 10.63 | 0.9 ± 0.28 | 180.4 ± 10.56 | 448.2 ± 26.48 | 3.5 ± 0.43 | 1.0 ± 0.11 |
| CMC20 | 13.2 ± 2.49 | 6.1 ± 1.27 | 91.2 ± 9.04 | 0.9 ± 0.22 | 180.0 ± 20.67 | 449.4 ± 29.48 | 3.6 ± 0.28 | 1.0 ± 0.14 |
| CMC40 | 13.3 ± 2.12 | 6.0 ± 0.87 | 92.1 ± 12.40 | 0.9 ± 0.29 | 180.4 ± 11.42 | 449.2 ± 28.27 | 3.6 ± 0.22 | 1.1 ± 0.12 |
| CMC60 | 13.0 ± 2.29 | 5.9 ± 1.54 | 91.6 ± 12.85 | 1.0 ± 0.17 | 182.2 ± 9.65 | 449.7 ± 35.31 | 3.5 ± 0.20 | 1.0 ± 0.09 |
| CMC80 | 13.2 ± 1.92 | 6.0 ± 1.32 | 91.7 ± 16.29 | 0.9 ± 0.22 | 180.4 ± 15.44 | 445.2 ± 24.09 | 3.5 ± 0.35 | 1.0 ± 0.12 |
| CMC100 | 12.8 ± 1.92 | 5.9 ± 1.54 | 91.3 ± 14.61 | 0.9 ± 0.21 | 183.4 ± 11.64 | 447.7 ± 27.40 | 3.6 ± 0.34 | 1.0 ± 0.15 |
| p–value | p > 0.9 | p > 0.9 | p > 0.9 | p > 0.9 | p > 0.9 | p > 0.9 | p > 0.9 | p > 0.2 |
| Experimental Diets | Lysozyme Activity (U/mL) | SOD (%) |
|---|---|---|
| CMC0 | 741.5 ± 79.91 | 69.3 ± 5.01 |
| CMC10 | 878.9 ± 93.41 | 70.8 ± 5.39 |
| CMC20 | 857.0 ± 206.31 | 75.2 ± 6.52 |
| CMC40 | 754.2 ± 36.66 | 70.4 ± 4.67 |
| CMC60 | 919.8 ± 40.51 | 73.9 ± 7.62 |
| CMC80 | 801.0 ± 164.46 | 73.6 ± 10.27 |
| CMC100 | 884.9 ± 53.8 | 68.3 ± 5.25 |
| p–value | p > 0.4 | p > 0.5 |
| Experimental Diets | Moisture | Crude Protein | Crude Lipid | Ash |
|---|---|---|---|---|
| CMC0 | 75.2 ± 0.26 | 17.4 ± 0.14 | 3.3 ± 0.01 | 3.7 ± 0.04 |
| CMC10 | 74.9 ± 0.56 | 17.5 ± 0.04 | 3.2 ± 0.09 | 3.6 ± 0.13 |
| CMC20 | 75.3 ± 0.18 | 17.5 ± 0.06 | 3.4 ± 0.09 | 3.7 ± 0.18 |
| CMC40 | 75.2 ± 0.09 | 17.6 ± 0.04 | 3.1 ± 0.14 | 3.5 ± 0.09 |
| CMC60 | 75.4 ± 0.24 | 17.5 ± 0.07 | 3.2 ± 0.07 | 3.6 ± 0.11 |
| CMC80 | 75.1 ± 0.14 | 17.4 ± 0.03 | 3.3 ± 0.11 | 3.7 ± 0.07 |
| CMC100 | 75.2 ± 0.19 | 17.5 ± 0.07 | 3.4 ± 0.14 | 3.5 ± 0.12 |
| p–value | p > 0.4 | p > 0.1 | p > 0.1 | p > 0.2 |
| Experimental Diets | ||||||||
|---|---|---|---|---|---|---|---|---|
| CMC0 | CMC10 | CMC20 | CMC40 | CMC60 | CMC80 | CMC100 | p–Value | |
| Essential amino acid (EAA) (%) | ||||||||
| Arginine | 0.35 ± 0.044 | 0.36 ± 0.040 | 0.37 ± 0.045 | 0.37 ± 0.025 | 0.39 ± 0.040 | 0.40 ± 0.030 | 0.41 ± 0.040 | p > 0.4 |
| Histidine | 0.36 ± 0.038 | 0.35 ± 0.031 | 0.33 ± 0.032 | 0.33 ± 0.029 | 0.31 ± 0.040 | 0.30 ± 0.030 | 0.29 ± 0.045 | p > 0.2 |
| Isoleucine | 0.69 ± 0.038 | 0.69 ± 0.040 | 0.68 ± 0.045 | 0.67 ± 0.040 | 0.66 ± 0.035 | 0.66 ± 0.055 | 0.65 ± 0.045 | p > 0.8 |
| Leucine | 1.16 ± 0.006 | 1.15 ± 0.020 | 1.15 ± 0.025 | 1.14 ± 0.035 | 1.13 ± 0.030 | 1.13 ± 0.025 | 1.12 ± 0.045 | p > 0.5 |
| Lysine | 0.51 ± 0.068 | 0.49 ± 0.025 | 0.49 ± 0.015 | 0.48 ± 0.045 | 0.47 ± 0.045 | 0.47 ± 0.035 | 0.46 ± 0.035 | p > 0.9 |
| Methionine | 0.46 ± 0.021 | 0.44 ± 0.020 | 0.42 ± 0.030 | 0.41 ± 0.025 | 0.41 ± 0.040 | 0.40 ± 0.020 | 0.39 ± 0.025 | p > 0.1 |
| Phenylalanine | 0.63 ± 0.012 | 0.61 ± 0.025 | 0.60 ± 0.050 | 0.59 ± 0.025 | 0.58 ± 0.040 | 0.56 ± 0.045 | 0.55 ± 0.035 | p > 0.2 |
| Threonine | 0.73 ± 0.030 | 0.73 ± 0.030 | 0.71 ± 0.025 | 0.70 ± 0.030 | 0.70 ± 0.035 | 0.69 ± 0.035 | 0.69 ± 0.030 | p > 0.2 |
| Tryptophan | 0.15 ± 0.029 | 0.15 ± 0.025 | 0.14 ± 0.040 | 0.14 ± 0.020 | 0.13 ± 0.025 | 0.13 ± 0.030 | 0.12 ± 0.025 | p > 0.7 |
| Valine | 0.78 ± 0.038 | 0.76 ± 0.040 | 0.75 ± 0.025 | 0.74 ± 0.040 | 0.74 ± 0.025 | 0.73 ± 0.025 | 0.71 ± 0.040 | p > 0.2 |
| Non-essential amino acid (NEAA) (%) | ||||||||
| Alanine | 1.17 ± 0.044 | 1.19 ± 0.035 | 1.20 ± 0.035 | 1.21 ± 0.025 | 1.23 ± 0.040 | 1.23 ± 0.035 | 1.25 ± 0.035 | p > 0.1 |
| Aspartic acid | 1.11 ± 0.047 | 1.09 ± 0.035 | 1.07 ± 0.040 | 1.07 ± 0.050 | 1.06 ± 0.040 | 1.04 ± 0.035 | 1.03 ± 0.035 | p > 0.2 |
| Cysteine | 0.19 ± 0.010 | 0.17 ± 0.030 | 0.16 ± 0.030 | 0.14 ± 0.045 | 0.14 ± 0.030 | 0.13 ± 0.030 | 0.12 ± 0.030 | p > 0.1 |
| Glutamic acid | 2.19 ± 0.061 | 2.21 ± 0.035 | 2.23 ± 0.025 | 2.24 ± 0.030 | 2.24 ± 0.035 | 2.26 ± 0.035 | 2.27 ± 0.025 | p > 0.2 |
| Glycine | 1.37 ± 0.136 | 1.39 ± 0.040 | 1.41 ± 0.055 | 1.41 ± 0.006 | 1.41 ± 0.062 | 1.42 ± 0.035 | 1.44 ± 0.055 | p > 0.9 |
| Proline | 0.82 ± 0.026 | 0.82 ± 0.031 | 0.83 ± 0.025 | 0.85 ± 0.015 | 0.85 ± 0.020 | 0.86 ± 0.030 | 0.87 ± 0.030 | p > 0.1 |
| Serine | 0.67 ± 0.032 | 0.67 ± 0.035 | 0.65 ± 0.025 | 0.64 ± 0.031 | 0.64 ± 0.035 | 0.63 ± 0.035 | 0.63 ± 0.035 | p > 0.5 |
| Tyrosine | 0.39 ± 0.058 | 0.38 ± 0.045 | 0.38 ± 0.050 | 0.36 ± 0.035 | 0.35 ± 0.025 | 0.35 ± 0.020 | 0.34 ± 0.045 | p > 0.6 |
| Experimental Diets | ||||||||
|---|---|---|---|---|---|---|---|---|
| CMC0 | CMC10 | CMC20 | CMC40 | CMC60 | CMC80 | CMC100 | p–Value | |
| C14:0 | 3.25 ± 0.050 a | 3.22 ± 0.047 ab | 3.17 ± 0.079 abc | 3.16 ± 0.040 abc | 3.14 ± 0.030 abc | 3.08 ± 0.075 bc | 3.04 ± 0.036 c | p < 0.004 |
| C16:0 | 16.56 ± 0.400 c | 16.73 ± 0.097 bc | 16.80 ± 0.056 abc | 16.96 ± 0.061 abc | 17.11 ± 0.176 ab | 17.23± 0.095 a | 17.28 ± 0.032 a | p < 0.001 |
| C18:0 | 4.58 ± 0.065 b | 4.64 ± 0.060 ab | 4.69 ± 0.051 ab | 4.71 ± 0.114 ab | 4.74 ± 0.062 ab | 4.78 ± 0.049 a | 4.80 ± 0.072 a | p < 0.03 |
| ∑SFA 1 | 24.39 ± 0.515 b | 24.59 ± 0.180 ab | 24.66 ± 0.148 ab | 24.83 ± 0.083 ab | 24.99 ± 0.150 ab | 25.09 ± 0.081 a | 25.12 ± 0.075 a | p < 0.01 |
| C14:1n−5 | 0.19 ± 0.010 | 0.17 ± 0.030 | 0.16 ± 0.031 | 0.16 ± 0.015 | 0.14 ± 0.006 | 0.15 ± 0.025 | 0.14 ± 0.029 | p > 0.1 |
| C16:1n−7 | 4.00 ± 0.015 a | 3.99 ± 0.070 ab | 3.97 ± 0.031 ab | 3.94 ± 0.042 ab | 3.94 ± 0.056 ab | 3.92 ± 0.050 ab | 3.88 ± 0.021 b | p < 0.03 |
| C17:1n−7 | 0.60 ± 0.045 | 0.58 ± 0.050 | 0.57 ± 0.040 | 0.57 ± 0.021 | 0.55 ± 0.040 | 0.54 ± 0.031 | 0.53 ± 0.020 | p > 0.3 |
| C18:1n−9 | 20.95 ± 0.095 g | 22.54 ± 0.154 f | 23.07 ± 0.176 e | 24.62 ±0.076 d | 26.46 ± 0.090 c | 28.70 ± 0.122 b | 30.05 ± 0.108 a | p < 0.0001 |
| C20:1n−9 | 0.35 ± 0.010 | 0.35 ± 0.015 | 0.34 ± 0.015 | 0.33 ± 0.030 | 0.33 ± 0.006 | 0.32 ± 0.026 | 0.31 ± 0.020 | p > 0.1 |
| ∑MUFA 2 | 26.09 ± 0.065 g | 27.64 ± 0.249 f | 28.12 ± 0.115 e | 29.61 ± 0.091 d | 31.42 ± 0.136 c | 33.63 ± 0.140 b | 34.91 ± 0.122 a | p < 0.0001 |
| C18:2n−6 | 21.24 ± 0.170 a | 21.08 ± 0.072 ab | 21.01 ± 0.100 ab | 20.87 ± 0.080 bc | 20.71 ± 0.074 cd | 20.63 ± 0.125 cd | 20.52 ± 0.080 d | p < 0.0001 |
| C18:3n−3 | 4.48 ± 0.100 a | 4.46 ± 0.107 a | 4.25 ± 0.064 ab | 4.16 ± 0.121 bc | 3.91 ± 0.114 cd | 3.74 ± 0.036 de | 3.62 ± 0.066 e | p < 0.0001 |
| C20:2n−6 | 1.56 ± 0.035 | 1.55 ± 0.050 | 1.54 ± 0.045 | 1.52 ± 0.060 | 1.51 ± 0.076 | 1.50 ± 0.035 | 1.48± 0.026 | p > 0.4 |
| C20:3n−3 | 0.26 ± 0.006 | 0.24 ± 0.006 | 0.24 ± 0.038 | 0.23 ± 0.035 | 0.23 ± 0.030 | 0.22 ± 0.025 | 0.22 ± 0.035 | p > 0.9 |
| C20:3n−6 | 0.74 ± 0.015 | 0.74 ± 0.031 | 0.73 ± 0.046 | 0.73 ± 0.035 | 0.72 ± 0.032 | 0.72 ± 0.046 | 0.71 ± 0.047 | p > 0.6 |
| C20:4n−6 | 1.11 ± 0.015 a | 1.08 ± 0.085 ab | 1.05 ± 0.050 ab | 1.00 ± 0.055 ab | 0.97 ± 0.035 abc | 0.94 ± 0.060 bc | 0.84 ± 0.050 c | p < 0.0001 |
| C20:5n−3 | 4.96 ± 0.035 a | 4.71 ± 0.144 ab | 4.65 ± 0.035 b | 4.58 ± 0.125 bc | 4.33 ± 0.119 c | 3.53 ± 0.035 d | 3.39 ± 0.131 d | p < 0.0001 |
| C22:5n−3 | 1.54 ± 0.015 a | 1.48 ± 0.079 ab | 1.43 ± 0.031 abc | 1.39 ± 0.029 abcd | 1.33 ± 0.076 bcd | 1.29 ± 0.057 cd | 1.26 ± 0.089 d | p < 0.0001 |
| C22:6n−3 | 8.56 ± 0.175 a | 8.39 ± 0.171 ab | 7.99 ± 0.125 b | 7.53 ± 0.100 c | 6.58 ± 0.159 d | 5.28 ± 0.091 e | 4.41 ± 0.163 f | p < 0.0001 |
| ∑n−3 HUFA 3 | 15.33 ± 0.230 a | 14.82 ± 0.229 b | 14.31 ± 0.110 c | 13.73 ± 0.190 d | 12.47 ± 0.104 e | 10.32 ± 0.046 f | 9.29 ± 0.100 g | p < 0.0001 |
| Unknown | 5.06 ± 0.115 | 4.05 ± 0.192 | 4.01 ± 0.168 | 3.54 ± 0.297 | 3.30 ± 0.267 | 3.43 ± 0.053 | 3.52 ± 0.045 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sim, Y.J.; Cho, S.H.; Kim, T. Fish Meal Substitution Effects with the Combined Animal Proteins in the Feeds of Olive Flounder (Paralichthys olivaceus) on Growth Performance, Feed Availability, and Disease Resistance against Streptococcus iniae. Animals 2024, 14, 1162. https://doi.org/10.3390/ani14081162
Sim YJ, Cho SH, Kim T. Fish Meal Substitution Effects with the Combined Animal Proteins in the Feeds of Olive Flounder (Paralichthys olivaceus) on Growth Performance, Feed Availability, and Disease Resistance against Streptococcus iniae. Animals. 2024; 14(8):1162. https://doi.org/10.3390/ani14081162
Chicago/Turabian StyleSim, Yu Jin, Sung Hwoan Cho, and Taeho Kim. 2024. "Fish Meal Substitution Effects with the Combined Animal Proteins in the Feeds of Olive Flounder (Paralichthys olivaceus) on Growth Performance, Feed Availability, and Disease Resistance against Streptococcus iniae" Animals 14, no. 8: 1162. https://doi.org/10.3390/ani14081162