

Molecular Determination of Sex from Down and Feather in Wild and Reared Monomorphic and Dimorphic Birds at Juvenile Age

 ,
,  , , and
, , and 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction
2.3. PCR and RFLP Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seddon, P.J.; Armstrong, D.P.; Maloney, R.F. Developing the science of reintroduction biology. Conserv. Biol. 2007, 21, 303–312. [Google Scholar] [CrossRef] [PubMed]
- IUCN (World Conservation Union). Guidelines for Re-introductions; IUCN/SSC Re-introduction Specialist Group, IUCN: Gland, Switzerland; Cambridge, UK, 1998. [Google Scholar]
- Wu, C.P.; Horng, Y.M.; Wang, R.T.; Yang, K.T.; Huang, M.C. A novel sex-specific DNA marker in Columbidae birds. Theriogenology 2007, 67, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.Y.; Hong, Y.J.; Park, S.K.; Kim, Y.J.; Choi, T.Y.; Lee, H.; Min, M.S. Application of two complementary molecular sexing methods for East Asian bird species. Genes. Genom. 2008, 30, 365–372. Available online: https://hdl.handle.net/10371/8289 (accessed on 6 February 2025).
- Forsman, D. The Raptors of Europe and the Middle East. In A Handbook of Field Identification; T. & A.D. Poyser: London, UK, 1999. [Google Scholar]
- Volodin, I.; Kaiser, M.; Matrosova, V.; Volodina, E.; Klenova, A.; Filatova, O.; Kholodova, M. The technique of noninvasive distant sexing for four monomorphic Dendrocygna whistling duck species by their loud whistles. Bioacoustics 2009, 18, 277–290. [Google Scholar] [CrossRef]
- Richner, H. Avian laparoscopy as a field technique for sexing birds and an assessment of its effects on wild birds. J. Field. Ornithol. 1989, 60, 137–142. [Google Scholar]
- Bercovitz, A.B.; Czekala, N.M.; Lasley, B.L. A new method of sex determination in monomorphic birds. J. Zoo Anim. Med. 1978, 9, 114–124. [Google Scholar] [CrossRef]
- Harris, T.; Walters, C. Chromosomal sexing of the black shouldered kite (Elanus caeruleus) (Aves: Accipitridae). Genetica 1982, 60, 19–20. [Google Scholar]
- Reynolds, S.J.; Martin, G.R.; Wallace, L.L.; Wearn, C.P.; Hughes, B.J. Sexing sooty terns on Ascension island from morphometric measurements. J. Zool. 2008, 274, 2–8. [Google Scholar] [CrossRef]
- Cerit, H.; Avanus, K. Sex identification in avian species using DNA typing methods. Worlds Poult. Sci. J. 2007, 63, 91–100. [Google Scholar] [CrossRef]
- Ellegren, H. First gene on the avian W chromosome (CHD) provides a tag for universal sexing of non-ratite birds. Proc. R. Soc. Lond. B Biol. Sci. 1996, 263, 1635–1641. [Google Scholar] [CrossRef]
- Griffiths, R.; Daan, S.; Dijkstra, C. Sex identification in birds using two CHD genes. Proc. R. Soc. Lond. B Biol. Sci. 1996, 263, 1251–1256. [Google Scholar] [CrossRef]
- Morinha, F.; Cabral, J.A.; Bastos, E. Molecular sexing of birds: A comparative review of polymerase chain reaction (PCR)-based methods. Theriogenology 2012, 78, 703–714. [Google Scholar] [CrossRef]
- Garcia-Raventós, A.; Muñoz-Mérida, A.; Lapiedra, O.; Unzeta, M.; Ferrandiz-Rovira, M.; Sol, D. Identification of sex-linked SNPmarkers in wild populations of monomorphic birds. Mol. Ecol. Resour. 2023, 23, 1905–1913. [Google Scholar] [CrossRef]
- Sacchi, P.; Soglia, D.; Maione, S.; Meneguz, G.; Campora, M.; Rasero, R. A non-invasive test for sex identification in short-toed Eagle (Circaetus gallicus). Mol. Cell Probes 2004, 18, 193–196. [Google Scholar] [CrossRef]
- Costantini, V.; Guaricci, A.C.; Laricchiuta, P.; Rausa, F.; Lacalandra, G.M. DNA sexing in Humboldt Penguins (Spheniscus humboldti) from feather samples. Anim. Reprod. Sci. 2008, 106, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Vucicevic, M.; Stevanovic, J.; Simeunovic, P.; Vucicevic, I.; Ðelic, N.; Stanimirovic, Z.; Stojic, V. Analysis of the CHD gene for sex determination of protected bird species. In Proceedings of the International Symposium on Hunting, “Modern Aspects of Sustainable Management of Game Population”, Zemun-Belgrade, Serbia, 22–24 June 2012; Volume 24, pp. 83–86. [Google Scholar]
- Turcu, M.C.; Pastiu, A.I.; Bel, L.V.; Pusta, D.L. A comparison of feathers and oral swab samples as DNA sources for molecular sexing in companion birds. Animals 2023, 13, 525. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, R.; Double, M.C.; Orr, K.; Dawson, R.J. A DNA test to sex most birds. Mol. Ecol. 1998, 7, 1071–1075. [Google Scholar] [CrossRef]
- Bachtrog, D.; Mank, J.E.; Peichel, C.L.; Kirkpatrick, M.; Otto, S.P.; Ashman, T.L.; Hahn, M.W.; Kitano, J.; Mayrose, I.; Ming, R.; et al. Sex determination: Why so many ways of doing it? PLoS Biol. 2014, 12, e1001899. [Google Scholar] [CrossRef]
- Bayard De Volo, S.; Reynolds, R.T.; Douglas, M.R.; Antolin, M.F. An improved extraction method to increase DNA yield from molted feathers. Condor 2008, 110, 762–766. [Google Scholar]
- Peniche, G.; Shaw, D.J.; Dures, S.G.; Clavaglia, S.; Thompson, D.B.A.; Anderson, N.E.; Meredith, A.L. Determining sex in golden eagle (Aquila chrysaetos) nestlings. Eur. J. Wildl. Res. 2023, 69, 1. [Google Scholar] [CrossRef]
- Turcu, M.C.; Pastiu, A.I.; Bel, L.V.; Pusta, D.L. Minimally invasive sampling methods for molecular sexing of wild and companion birds. Animals 2023, 13, 3417. [Google Scholar] [CrossRef] [PubMed]
- Newton, I. The Sparrowhawk, 1st ed.; T and A.D. Poyser: Waterhouses, UK, 1986. [Google Scholar]
- Alonso, R.; Palomare, L.E.; Martinez, J.A. Sex determination in Eurasian sparrowhawks (Accipiter nisus). J. Raptor Res. 2011, 45, 48–55. [Google Scholar] [CrossRef]
- Canon, N.R.; Tell, L.A.; Needham, M.L.; Gardner, I.A. Flow cytometric analysis of nuclear DNA for sex identification in three psittacine species. Am. J. Vet. Res. 2000, 61, 847–850. [Google Scholar] [CrossRef] [PubMed]
- Elnomrosy, S.M.; Hagag, N.M.; AbdAllah, M.I.; Kolenda, R.; Zacharski, M. Application of loop-mediated isothermal amplification (LAMP) in sex identification of parrots bred in Egypt. Biology 2022, 11, 565. [Google Scholar] [CrossRef]
- Horng, Y.M.; Wu, C.P.; Wang, Y.C.; Huang, M.C. A novel molecular genetic marker for gender determination of pigeons. Theriogenology 2006, 65, 1759–1768. [Google Scholar] [CrossRef]
- Griffiths, R.; Tiwari, B. The isolation of molecular genetic markers for the identification of sex. Proc. Natl. Acad. Sci. USA 1993, 90, 8324–8326. [Google Scholar] [CrossRef] [PubMed]
- Mills, L.S.; Citta, J.J.; Lair, K.P.; Schwartz, M.K.; Tallmon, D.A. Estimating animal abundance using noninvasive DNA sampling: Promise and pitfalls. Ecol. Appl. 2000, 10, 283–294. [Google Scholar] [CrossRef]
- Griffiths, R.; Orr, K. The use of amplified fragment length polymorphism (AFLP) in the isolation of sex-specific markers. Mol. Ecol. 1999, 8, 671–674. [Google Scholar] [CrossRef]

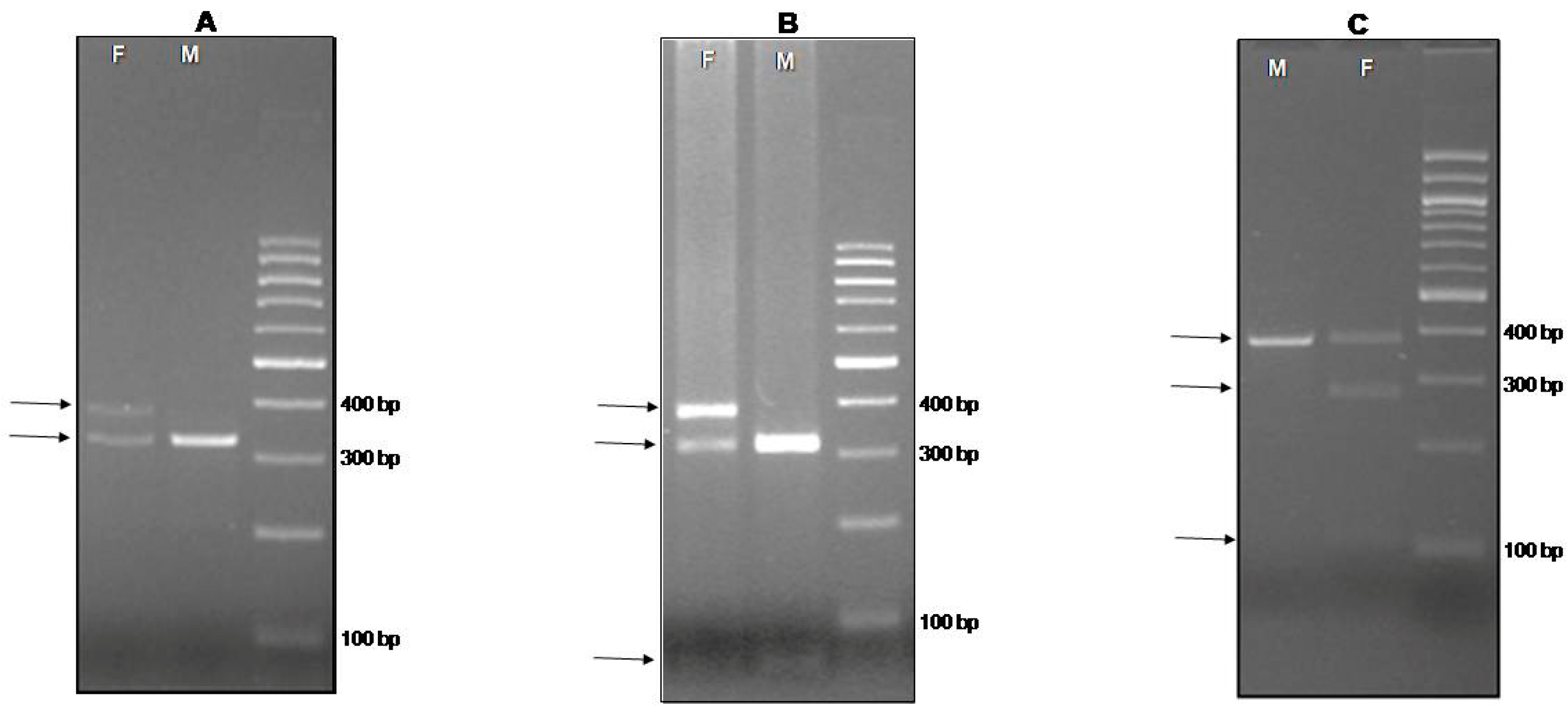


{kind=link}
{kind=link}
{kind=link}
| ORDER | SPECIES | ♂ P2/P8 | ♀ P2/P8 | ♂ HaeIII | ♀ HaeIII | ♂ Asp 700I | ♀ Asp 700I |
|---|---|---|---|---|---|---|---|
| Accipitriformes | Accipiter gentilis | 390 | 390 | 290 + 100 | 390 + 290 + 100 | ||
| Accipiter nisus | 390 | 390 | 330 + 60 | 390 + 330 + 60 | |||
| Aquila chrysaetos | 390 | 390 | 320 + 70 | 390 + 320 + 70 | |||
| Aquila rapax | 380 | 380 | 330 + 50 | 380 + 330 + 50 | |||
| Aquila spilogaster | 380 | 380 | 330 + 50 | 380 + 330 + 50 | |||
| Buteo buteo | 390 | 390 | 320 + 70 | 390 + 320 + 70 | |||
| Cathartes aura | 380 | 400 + 380 | |||||
| Circaetus gallicus | 390 | 390 | 320 + 70 | 390 + 320 + 70 | |||
| Circus aeruginosus | 390 | 390 | 330 + 60 | 390 + 330 + 60 | |||
| Circus pygargus | 390 | 390 | 320 + 70 | 390 + 320 + 70 | |||
| Falco eleonorae | 390 | 390 | 330 + 60 | 390 + 330 + 60 | |||
| Gypohierax angolensis | 380 | 390 + 380 | |||||
| Gyps africanus | 390 | 390 | 330 + 60 | 390 + 330 + 60 | |||
| Gyps fulvus | 390 | 390 | 330 + 60 | 390 + 330 + 60 | |||
| Gyps rueppelli | 390 | 390 | 330 + 60 | 390 + 330 + 60 | |||
| Haliaeetus albicilla | 380 | 380 | 340 + 40 | 380 + 340 + 40 | |||
| Hieraaetus pennatus | 380 | 380 | 330 + 50 | 380 + 330 + 50 | |||
| Milvus milvus | 390 | 390 | 320 + 70 | 390 + 320 + 70 | |||
| Neophron percnopterus | 370 | 380 + 370 | |||||
| Parabuteo unicinctus | 390 | 390 | 330 + 60 | 390 + 330 + 60 | |||
| Pernis apivorus | 380 | 390 + 380 | |||||
| Sarcogyps calvus | 400 | 400 | 320 + 80 | 400 + 320 + 80 | |||
| Sarcoramphus papa | 380 | 400 + 380 | |||||
| Anseriformes | Alopochen aegyptiaca | 360 | 370 + 360 | ||||
| Anas platyrhynchos | 380 | 380 | 320 + 60 | 380 + 320 + 60 | |||
| Aythya fuligula | 360 | 370 + 360 | |||||
| Chauna chavaria | 370 | 370 | 370 | 370 | 260 + 110 | 370 + 260 + 110 | |
| Cygnus atratus | 370 | 380 + 370 | |||||
| Netta rufina | 360 | 380 + 360 | |||||
| Oxyura leucocephala | 370 | 380 + 370 | |||||
| Spatula querquedula | 370 | 380 + 370 | |||||
| Apodiformes | Apus apus | 380 | 380 | 330 + 50 | 380 + 330 + 50 | ||
| Bucerotiformes | Aceros nipalensis | 380 | 380 | 320 + 60 | 380 + 320 + 60 | ||
| Bucorvus leadbeateri | 390 | 390 | 330 + 60 | 390 + 330 + 60 | |||
| Upupa epops | 380 | 390 + 380 | |||||
| Charadriiformes | Actitis hypoleucos | 370 | 380 + 370 | ||||
| Chroicocephalus ridibundus | 360 | 380 + 360 | |||||
| Burhinus oedicnemus | 360 | 380 + 360 | |||||
| Ichthyaetus melanocephalus | 380 | 380 | 310 + 70 | 380 + 310 + 70 | |||
| Himantopus himantopus | 380 | 380 | 340 + 40 | 380 + 340 + 40 | |||
| Numenius arquata | 380 | 390 + 380 | |||||
| Scolopax rusticola | 370 | 380 + 370 | |||||
| Vanellus vanellus | 390 | 390 | 320 + 70 | 390 + 320 + 70 | |||
| Ciconiiformes | Ciconia ciconia | 390 | 390 | 320 + 70 | 390 + 320 + 70 | ||
| Columbiformes | Columba livia | 350 | 370 + 350 | ||||
| Columba livia domestica | 350 | 370 + 350 | |||||
| Streptopelia decaocto | 390 | 390 | 300 + 90 | 390 + 300 + 90 | |||
| Coraciiformes | Alcedo atthis | 370 | 390 + 370 | ||||
| Coracias garrulus | 380 | 390 + 380 | |||||
| Dacelo leachii | 360 | 380 + 360 | |||||
| Merops apiaster | 380 | 380 | 340 + 40 | 380 + 340 + 40 | |||
| Cuculiformes | Cuculus canorus | 380 | 380 | 320 + 60 | 380 + 320 + 60 | ||
| Falconiformes | Falco vespertinus | 390 | 390 | 330 + 60 | 390 + 330 + 60 | ||
| Caracara plancus | 380 | 400 + 380 | |||||
| Falco ardosiaceus | 400 | 400 | 350 + 50 | 400 + 350 + 50 | |||
| Falco biarmicus | 380 | 380 | 330 + 50 | 380 + 330 + 50 | |||
| Falco cherrug | 380 | 390 + 380 | |||||
| Falco jugger | 390 | 390 | 330 + 60 | 390 + 330 + 60 | |||
| Falco naumanni | 400 | 400 | 350 + 50 | 400 + 350 + 50 | |||
| Falco peregrinus | 380 | 390 + 380 | |||||
| Falco subbuteo | 400 | 400 | 350 + 50 | 400 + 350 + 50 | |||
| Galliformes | Alectoris barbara | 370 | 380 + 370 | ||||
| Alectoris graeca | 370 | 380 + 370 | |||||
| Coturnix coturnix | 390 | 390 | 330 + 60 | 390 + 330 + 60 | |||
| Pavo cristatus | 360 | 380 + 360 | |||||
| Pavo muticus | 360 | 380 + 360 | |||||
| Phasianus colchicus | 370 | 380 + 370 | |||||
| Polypectron napoleonis | 380 | 380 | 310 + 70 | 380 + 310 + 70 | |||
| Gruiformes | Balearica regulorum | 390 | 390 | 320 + 70 | 390 + 320 + 70 | ||
| Fulica atra | 380 | 380 | 340 + 40 | 380 + 340 + 40 | |||
| Grus grus | 390 | 390 | 320 + 70 | 390 + 320 + 70 | |||
| Rallus aquaticus | 380 | 390 + 380 | |||||
| Otidiformes | Otis tarda | 380 | 390 + 380 | ||||
| Passeriformes | Acridotheres tristis | 370 | 390 + 370 | ||||
| Cardinalis cardinalis | 320 | 360 + 320 | |||||
| Carduelis carduelis | 320 | 360 + 320 | |||||
| Chloris chloris | 340 | 360 + 340 | |||||
| Coloeus monedula | 370 | 390 + 370 | |||||
| Corvus corone | 360 | 380 + 360 | |||||
| Corvus corax | 360 | 380 + 360 | |||||
| Erythrura psittacea | 370 | 390 + 370 | |||||
| Gracula religiosa | 380 | 400 + 380 | |||||
| Lanius collurio | 370 | 390 + 370 | |||||
| Leiothrix lutea | 370 | 380 + 370 | |||||
| Monticola solitarius | 380 | 380 | 330 + 50 | 380 + 330 + 50 | |||
| Oriolus oriolus | 370 | 370 | 290 + 80 | 370 + 290 + 80 | |||
| Parus major | 400 | 400 | 350 + 50 | 400 + 350 + 50 | |||
| Pica pica | 370 | 390 + 370 | |||||
| Pinicola enucleator | 360 | 380 + 360 | |||||
| Serinus serinus | 350 | 350 | 270 + 80 | 350 + 270 + 80 | |||
| Spinus atratus | 360 | 380 + 360 | |||||
| Spinus magellanicus | 330 | 360 + 330 | |||||
| Turdus iliacus | 360 | 380 + 360 | |||||
| Turdus philomelos | 360 | 380 + 360 | |||||
| Turdus pilaris | 370 | 390 + 370 | |||||
| Turdus viscivorus | 360 | 380 + 360 | |||||
| Pelecaniformes | Ardea cinerea | 380 | 380 | 320 + 60 | 380 + 320 + 60 | ||
| Ardea purpurea | 380 | 380 | 320 + 60 | 380 + 320 + 60 | |||
| Egretta garzetta | 370 | 380 + 370 | |||||
| Endocimus ruber | 390 | 390 | 340 + 50 | 390 + 340 + 50 | |||
| Geronticus eremita | 390 | 390 | 340 + 50 | 390 + 340 + 50 | |||
| Ixobrychus minutus | 390 | 390 | 330 + 60 | 390 + 330 + 60 | |||
| Nycticorax nycticorax | 390 | 390 | 330 + 60 | 390 + 330 + 60 | |||
| Pelecanus crispus | 400 | 400 | 330 + 70 | 400 + 330 + 70 | |||
| Phoenicopteriformes | Phoenicopterus chilensis | 390 | 390 | 390 | 390 | 280 + 110 | 390 + 280 + 110 |
| Phoenicopterus roseus | 390 | 390 | 390 | 390 | 280 + 110 | 390 + 280 + 110 | |
| Podicipediformes | Podiceps cristatus | 370 | 380 + 370 | ||||
| Psittaciformes | Agapornis taranta | 360 | 380 + 360 | ||||
| Agapornis fischeri | 360 | 380 + 360 | |||||
| Agapornis personatus | 360 | 380 + 360 | |||||
| Agapornis roseicollis | 360 | 380 + 360 | |||||
| Alipiopsitta xanthops | 380 | 400 + 380 | |||||
| Amazona amazonica | 390 | 400 + 390 | |||||
| Amazona farinosa | 390 | 400 + 390 | |||||
| Ara ararauna | 380 | 390 + 380 | |||||
| Ara chloropterus | 380 | 390 + 380 | |||||
| Aratinga solstitialis | 390 | 400 + 390 | |||||
| Barnardius zonarius | 380 | 400 + 380 | |||||
| Cacatua galerita | 380 | 380 | 320 + 60 | 380 + 320 + 60 | |||
| Cacatua sulphurea | 380 | 380 | 320 + 60 | 380 + 320 + 60 | |||
| Cyanoliseus patagonus | 370 | 390 + 370 | |||||
| Eos bornea | 370 | 390 + 370 | |||||
| Lorius garrulus | 370 | 390 + 370 | |||||
| Melopsittacus undulatus | 380 | 400 + 380 | |||||
| Myiopsitta monachus | 370 | 390 + 370 | |||||
| Nandaius nenday | 370 | 390 + 370 | |||||
| Nymphicus hollandicus | 360 | 380 + 360 | |||||
| Pionites leucogaster | 370 | 390 + 370 | |||||
| Pionus senilis | 370 | 390 + 370 | |||||
| Platycercus elegans | 380 | 390 + 380 | |||||
| Platycercus eximius | 380 | 390 + 380 | |||||
| Poicephalus senegalus | 370 | 390 + 370 | |||||
| Psittacula eupatria | 370 | 390 + 370 | |||||
| Psittacula krameri | 370 | 390 + 370 | |||||
| Psittacus erithacus | 370 | 390 + 370 | |||||
| Pyrrhura frontalis | 370 | 390 + 370 | |||||
| Pyrrhura picta | 370 | 390 + 370 | |||||
| Trichoglossus haematodus | 370 | 390 + 370 | |||||
| Trichoglossus moluccanus | 370 | 390 + 370 | |||||
| Sphenisciformes | Spheniscus humboldti | 370 | 380 + 370 | ||||
| Strigiformes | Asio flammeus | 390 | 390 | 320 + 70 | 390 + 320 + 70 | ||
| Asio otus | 390 | 390 | 320 + 70 | 390 + 320 + 70 | |||
| Athene noctua | 380 | 380 | 320 + 60 | 380 + 320 + 60 | |||
| Bubo africanus | 380 | 380 | 320 + 60 | 390 + 320 + 60 | |||
| Bubo bubo | 390 | 390 | 320 + 70 | 390 + 320 + 70 | |||
| Bubo virginianus | 380 | 380 | 320 + 60 | 390 + 320 + 60 | |||
| Otus scops | 380 | 380 | 320 + 60 | 380 + 320 + 60 | |||
| Strix aluco | 360 | 370 + 360 | |||||
| Strix rufipes | 360 | 370 + 360 | |||||
| Strix uralensis | 360 | 370 + 360 | |||||
| Tyto alba | 390 | 390 | 320 + 70 | 390 + 320 + 70 | |||
| Suliformes | Phalacrocorax carbo | 390 | 390 | 340 + 50 | 390 + 340 + 50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guaricci, A.C.; Cinone, M.; Desantis, S.; Lacalandra, G.M.; Albrizio, M. Molecular Determination of Sex from Down and Feather in Wild and Reared Monomorphic and Dimorphic Birds at Juvenile Age. Animals 2025, 15, 892. https://doi.org/10.3390/ani15060892
Guaricci AC, Cinone M, Desantis S, Lacalandra GM, Albrizio M. Molecular Determination of Sex from Down and Feather in Wild and Reared Monomorphic and Dimorphic Birds at Juvenile Age. Animals. 2025; 15(6):892. https://doi.org/10.3390/ani15060892
Chicago/Turabian StyleGuaricci, Antonio Ciro, Mario Cinone, Salvatore Desantis, Giovanni Michele Lacalandra, and Maria Albrizio. 2025. "Molecular Determination of Sex from Down and Feather in Wild and Reared Monomorphic and Dimorphic Birds at Juvenile Age" Animals 15, no. 6: 892. https://doi.org/10.3390/ani15060892
APA StyleGuaricci, A. C., Cinone, M., Desantis, S., Lacalandra, G. M., & Albrizio, M. (2025). Molecular Determination of Sex from Down and Feather in Wild and Reared Monomorphic and Dimorphic Birds at Juvenile Age. Animals, 15(6), 892. https://doi.org/10.3390/ani15060892