Evaluation of Brassica Vegetables as Potential Feed for Ruminants

 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples of Cabbage Wastes
2.2. Animals and Feeding
2.3. Experimental Design
2.3.1. Experiment 1: In Vitro Ruminal Fermentation and Intestinal Digestibility of Discarded Vegetables
2.3.2. Experiment 2: In Vitro Fermentation and In Situ Degradability of Diets Containing Increasing Amounts of Brussels Sprouts
2.4. Chemical Analyses
2.5. Calculations and Statistical Analyses
3. Results and Discussion
3.1. Experiment 1: Chemical Composition, In Vitro Ruminal Fermentation, and In Vitro Intestinal Digestibility of Vegetables
3.2. Experiment 2: In Vitro Fermentation and In Situ Degradability of Diets Containing Increasing Amount of Brussels Sprouts
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- FAO. Global Food Losses and Food Waste-Extent, Causes and Prevention; FAO: Rome, Italy, 2011. [Google Scholar]
- Rivin, J.; Miller, Z.; Matel, O. Using Food Waste as Livestock Feed. University of Wisconsin System Board of Regents and University of Wisconsin-Extension, Cooperative Extension: United States, 2012. Available online: https://outagamie.extension.wisc.edu/files/2012/10/Using-Food-Waste-as-Livestock-Feed.pdf (accessed on 19 July 2019).
- Pignata, G.; Nicola, S. Profitability, marketing, and vegetable loss and waste. In Good Agricultural Practices for greenhouse vegetable production in the South. East. European countries—Principles for sustainable intensification of smallholder farms; Baudoin, W., Nersisyan, A., Shamilov, A., Hodder, A., Gutierrez, D., de Pascale, S., Nicola, S., Gruda, N., Urban, L., Tanny, J., Eds.; FAO: Roma, Italy, 2017; pp. 245–267. [Google Scholar]
- Nkosi, B.D.; Meeske, R.; Ratsaka, M.M.; Langa, T.; Motiang, M.D.; Groenewald, I.B. Effects of dietary inclusion of discarded cabbage (Brassica oleracea var. capitata) on the growth performance of South African Dorper lambs. S. Afr. J. Anim. Sci. 2016, 46, 35–41. [Google Scholar] [CrossRef][Green Version]
- Bakshi, M.P.S.; Wadhwa, M.; Makkar, H. Waste to worth: Vegetable wastes as animal feed. Cab. Rev. 2016, 11, 1–26. [Google Scholar] [CrossRef]
- Duncan, A.J.; Milne, J.A. Rumen microbial degradation of allyl cyanide as a possible explanation for the tolerance of sheep to brassica-derived glucosinolates. J. Sci. Food Agric. 1992, 58, 15–19. [Google Scholar] [CrossRef]
- Mekasha, Y.; Tegegne, A.; Yami, A.; Umunna, N.N. Evaluation of non-conventional agro-industrial by-products as supplementary feeds for ruminants: In vitro and metabolism study with sheep. Small Rumin. Res. 2002, 44, 25–35. [Google Scholar] [CrossRef]
- Ngu, N.T.; Ledin, I. Effects of feeding wastes from Brassica species on growth of goats and pesticide/insecticide residues in goat meat. Asian-Aust. J. Anim. Sci 2005, 18, 197–202. [Google Scholar] [CrossRef]
- Wadhwa, M.; Kaushal, S.; Bakshi, M.P.S. Nutritive evaluation of vegetable wastes as complete feed for goat bucks. Small Rum. Res. 2006, 64, 279–284. [Google Scholar] [CrossRef]
- Wadhwa, M.; Bakshi, M.P.S. Utilization of Fruit and Vegetable Wastes as Livestock Feed and as Substrates for Generation of Other Value-Added Products; Makkar, H.P.S., Ed.; FAO: Rome, Italy, 2013. [Google Scholar]
- Mahgoub, O.; Kadim, I.T.; Eltahir, Y.; Al-Lawatia, S.; Al-Ismaili, A.M. Nutritional Value of Vegetable Wastes as Livestock Feed. Sultan Qaboos Univ. J. Sci. 2018, 23, 78–84. [Google Scholar] [CrossRef]
- Marino, C.T.; Hector, B.; Rodrigues, P.M.; Borgatti, L.O.; Meyer, P.M.; Alves da Silva, E.J.; Ørskov, E.R. Characterization of vegetables and fruits potential as ruminant feed by in vitro gas production technique. Livest. Res. Rural Dev. 2010, 22, 168. [Google Scholar]
- Kazemi, M.; Tornaghan, A.E.; Abadi, E.I.K.; Tazik, S.A.; Tohidi, R. Red (Brassica oleracea var. capitata) and White (Brassica oleracea var. botrytis) Cabbage Leaves Nutritional Value as Forage Feed: Comparison Study of In Vitro Gas Production and Determination of Chemical Composition. Direct. Res. J. Agric. Food Sci. 2016, 4, 176–181. [Google Scholar]
- Neugart, S.; Baldermann, S.; Hanschen, F.S.; Klopsch, R.; Wiesner-Reinhold, M.; Schreiner, M. The intrinsic quality of brassicaceous vegetables: How secondary plant metabolites are affected by genetic, environmental, and agronomic factors. Sci. Hortic. 2018, 233, 460–478. [Google Scholar] [CrossRef]
- Taljaard, T.L. Cabbage poisoning in ruminants. J. S. Afr. Vet. Assoc. 1993, 64, 96–100. [Google Scholar] [PubMed]
- Goering, M.K.; Van Soest, P.J. Forage Fiber Analysis (Apparatus, Reagents, Procedures and Some Applications). In Agricultural Handbook; Agriculture Handbook No 379; Agricultural Research Services: Washington, DC, USA, 1970. [Google Scholar]
- Marcos, C.N.; de Evan, T.; Molina-Alcaide, E.; Carro, M.D. Nutritive value of tomato pomace for ruminants and its influence on in vitro methane production. Animals 2019, 9, 343. [Google Scholar] [CrossRef] [PubMed]
- Gargallo, S.; Calsamiglia, S.; Ferret, A. Technical note: A modified three-step in vitro procedure to determine intestinal digestion of proteins. J. Anim. Sci. 2006, 84, 2163–2167. [Google Scholar] [CrossRef] [PubMed]
- Belverdy, M.S.; Alamouti, A.A.; Khadem, A.A.; González, J.; Carro, M.D.; Kianmehr, M.H.; Azizi, M.H. Evaluation of a novel method for ruminal protection of soybean meal protein using different fat sources. Arch. Anim. Nutr. 2019, 73, 158–169. [Google Scholar]
- Weatherburn, M.W. Phenol-hypochlorite reaction for determination of ammonia. Anal. Chem. 1967, 39, 971–974. [Google Scholar] [CrossRef]
- García-Martínez, R.; Ranilla, M.J.; Tejido, M.L.; Carro, M.D. Effects of disodium fumarate on in vitro rumen microbial growth, methane production and fermentation of diets differing in their forage concentrate ratio. Br. J. Nutr. 2005, 94, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M.E.; Ranilla, M.J.; Tejido, M.L.; Ramos, S.; Carro, M.D. The effect of the diet fed to donor sheep on in vitro methane production and ruminal fermentation of diets of variable composition. Anim. Feed Sci. Technol. 2010, 158, 126–135. [Google Scholar] [CrossRef]
- SAS Institute. SAS/STAT® Users Guide, Version 9.3; SAS Inst. Inc.: Cary, NC, USA, 2017. [Google Scholar]
- Ranilla, M.J.; López, S.; Giráldez, F.J.; Valdés, C.; Carro, M.D. Comparative digestibility and digesta flow kinetics in two breeds of sheep. Anim. Sci. 1998, 66, 389–396. [Google Scholar] [CrossRef]
- Demeyer, D. Quantitative aspects of microbial metabolism in the rumen and hindgut. In Rumen Microbial Metabolism and Ruminant Digestion; Jouany, J.P., Ed.; INRA Editions: Paris, France, 1991; pp. 217–237. [Google Scholar]
- NRC (National Research Council). Nutrient Requirements of Dairy Cattle, 7th ed.; National Academy of Sciences: Washington, DC, USA, 2001. [Google Scholar]
- Cone, J.W.; van Gelder, A.H. Influence of protein fermentation on gas production profiles. Anim. Feed Sci.Technol. 1999, 76, 251–264. [Google Scholar] [CrossRef]
- Wallace, R.J.; Cotta, M.A. The Rumen Microbial Ecosystem; Hobson, P.N., Stewart, C.S., Eds.; Elsevier Applied Science: London, UK, 1988. [Google Scholar]
- AFRC (Agricultural Food Research Council). Energy and Protein Requirements of Ruminants; CAB International: Wallingford, UK, 1993. [Google Scholar]
- Tamminga, S.; van Straalen, W.M.; Subnel, A.P.J.; Meijer, R.G.M.; Steg, A.; Wever, C.J.G.; Blok, M.C. The Dutch protein evaluation system: The DVE/OEB-system. Livest. Prod. Sci. 1994, 40, 139–155. [Google Scholar] [CrossRef]
- Madsen, J.; Hvelplund, T.; Weisbjerg, M.R.; Bertilsson, J.; Olsson, I.; Sporndly, R.; Harstad, O.M.; Volden, H.; Tuori, M.; Var-vikko, T.; et al. The AAT/PBV protein evaluation system for ruminants—a revision. Nor. J. Agric. Sci. Suppl. 1995, 19, 1–37. [Google Scholar]
- Van Soest, P.J. Nutritional Ecology of the Ruminant, 2nd ed.; Cornell University Press: Ithaca, NY, USA, 1994. [Google Scholar]
- Carro, M.D.; Lebzien, P.; Rohr, K. Effects of pore size of nylon bags and dilution rate on fermentation parameters in a semi-continuous artificial rumen. Small Rumin. Res. 1995, 15, 113–119. [Google Scholar] [CrossRef]


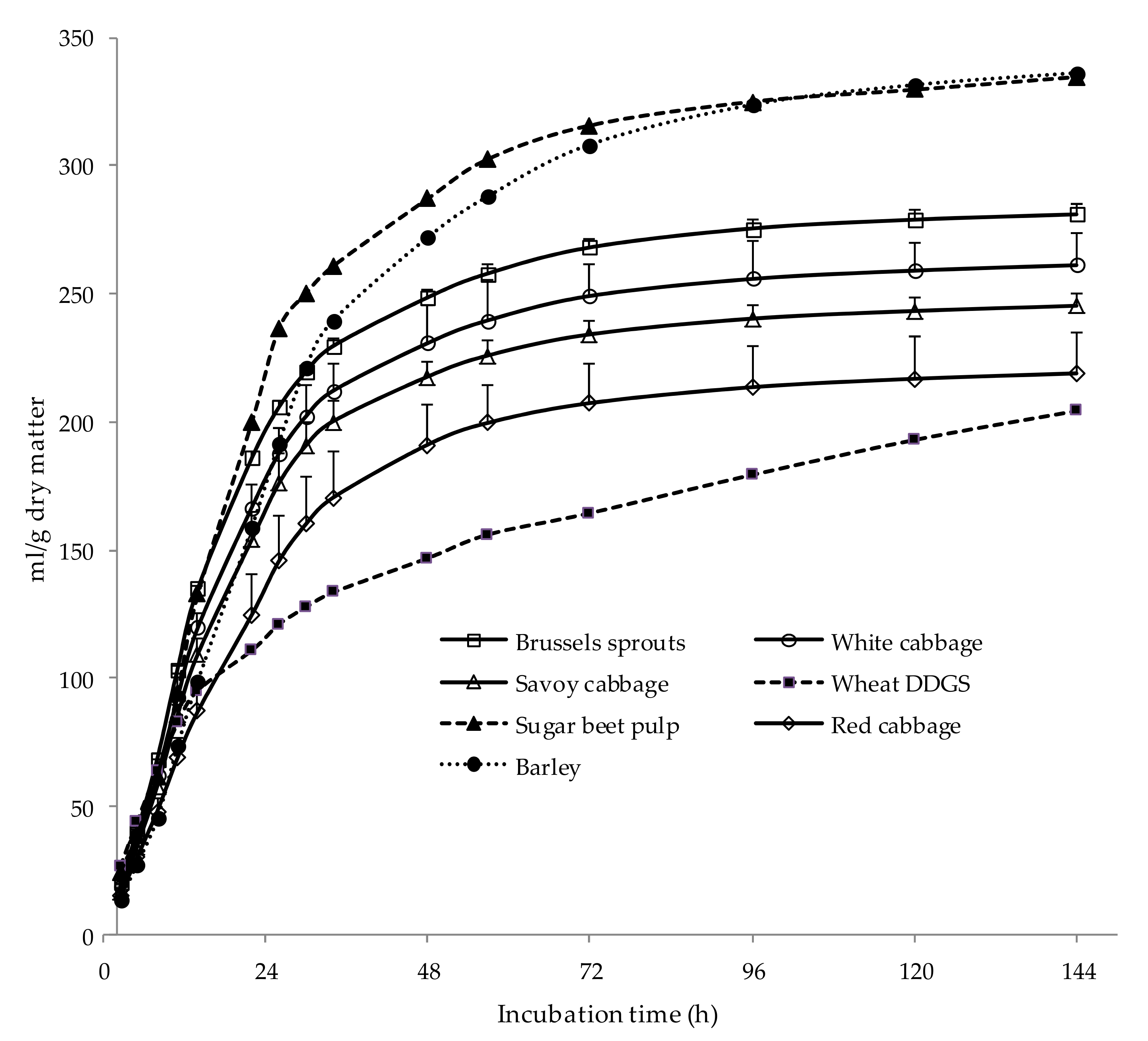{kind=link}
| Item | Cabbages | SEM 2 | p | Reference Feeds | |||||
|---|---|---|---|---|---|---|---|---|---|
| Brussels Sprouts | White Cabbage | Savoy Cabbage | Red Cabbage | Barley | Sugar Beet Pulp | Wheat DDGS | |||
| Dry matter (%) | 16.3 b | 5.64 a | 7.14 a | 6.60 a | 0.388 | <0.001 | 89.9 | 89.1 | 92.2 |
| Organic matter | 92.2 b | 84.7 a | 86.4 a | 89.1 a | 0.44 | <0.001 | 97.3 | 94.9 | 95.5 |
| Crude protein (CP) | 24.8 b | 19.5 a | 20.7 a | 19.8 a | 0.59 | <0.001 | 12.4 | 9.44 | 32.9 |
| Ether extract | 2.90 a | 3.57 ab | 4.36 a | 4.54 a | 0.220 | 0.002 | 3.16 | 0.80 | 4.61 |
| Sugars | 41.4 | 35.3 | 34.2 | 27.2 | 3.35 | 0.095 | 3.69 | 13.5 | 6.97 |
| Neutral detergent fiber (NDF) | 17.5 a | 23.5 ab | 28 b | 25.4 b | 1.54 | 0.007 | 22.7 | 48 | 29.5 |
| Acid detergent fiber | 10.3 a | 15.4 ab | 17.3 b | 17.6 b | 0.751 | <0.001 | 5.23 | 24.2 | 11.2 |
| Lignin | 0.25 | 1.53 | 1.97 | 2.43 | 0.574 | 0.119 | 1.22 | 2.16 | 3.33 |
| Hemicellulose | 7.24 | 8.15 | 10.7 | 8.87 | 0.845 | 0.087 | 19.6 | 23.9 | 18.3 |
| Lignin (% of NDF) | 1.43 | 6.20 | 7.01 | 9.40 | 1.914 | 0.092 | 5.37 | 4.49 | 11.2 |
| NDICP 1 (% CP) | 4.89 | 15.8 | 17.8 | 13.9 | 3.26 | 0.091 | 14.5 | 55 | 28.9 |
| Item | Cabbages | SEM 3 | p | Reference Feeds | |||||
|---|---|---|---|---|---|---|---|---|---|
| Brussels Sprouts | White Cabbage | Savoy Cabbage | Red Cabbage | Barley | Sugar Beet Pulp | Wheat DDGS | |||
| Gas production parameters 1 | |||||||||
| A (mL/g) | 275 c | 219 a | 229 ab | 233 b | 3.27 | <0.001 | 352 | 329 | 185 |
| c (%/h) | 5.55 c | 4.51 b | 4.75 b | 3.71 a | 0.17 | <0.001 | 5 | 5.21 | 4.15 |
| Lag (h) | 2.59 | 2.44 | 3.03 | 2.73 | 0.187 | 0.169 | 2.85 | 3.83 | 0 |
| AGPR (mL/h) | 9.11 c | 6.17 ab | 6.49 b | 5.47 a | 0.239 | <0.001 | 10.5 | 9.52 | 5.55 |
| DMED (%) | 58.4 b | 51.7 a | 49.1 a | 50.4 a | 0.69 | <0.001 | 45 | 45.2 | 38.4 |
| Fermentation parameters 2 | |||||||||
| Gas (mL) | 39.5 c | 34.2 a | 35 a | 36.8 b | 0.51 | <0.001 | 49.9 | 43.9 | 26.3 |
| pH | 6.52 a | 6.61 bc | 6.65 c | 6.60 b | 0.011 | <0.001 | 6.60 | 6.56 | 6.73 |
| Total volatile fatty acids (VFA; µmol) | 1632 | 1636 | 1612 | 1673 | 19.6 | 0.186 | 1452 | 1700 | 1311 |
| Individual VFA (mol/100 mol) | |||||||||
| Acetate (Ac) | 57.5 a | 62.6 b | 62.5 b | 62.9 b | 0.25 | <0.001 | 56.6 | 65.5 | 53.4 |
| Propionate (Pr) | 28.2 b | 24.5 a | 24.3 a | 24.1 a | 0.25 | <0.001 | 22.2 | 25.2 | 33.3 |
| Butyrate | 9.06 b | 8.18 a | 8.04 a | 8.31 a | 0.104 | <0.001 | 17.5 | 6.93 | 6.34 |
| Minor VFA | 5.23 b | 4.73 a | 5.15 b | 4.65 a | 0.106 | <0.001 | 3.70 | 2.31 | 6.96 |
| Ac/Pr (mol/mol) | 2.04 a | 2.55 b | 2.57 b | 2.60 b | 0.031 | <0.001 | 2.55 | 2.60 | 1.61 |
| NH3-N (mg/L) | 288 c | 223 a | 257 b | 218 a | 5.4 | <0.001 | 156 | 86.8 | 223 |
| Item | Brussels Sprouts | White Cabbage | Savoy Cabbage | Red Cabbage | SEM 1 | p |
|---|---|---|---|---|---|---|
| DM rumen (%) | 93.1 b | 91.9 b | 89.5 ab | 86.9 a | 1.03 | 0.001 |
| CP rumen (%) | 91.6 a | 94.4 b | 92.7 ab | 91.8 a | 0.77 | 0.042 |
| DM intestinal (%) | 73.5 c | 45.7 b | 44.2 b | 35.3 a | 2.57 | <0.001 |
| CP intestinal (%) | 90.2 c | 71.2 ab | 74.1 b | 61.4 a | 3.24 | 0.008 |
| Item | Diet | |||
|---|---|---|---|---|
| Control | BS8 | BS16 | BS24 | |
| Diet ingredients (g/100 g fresh matter) | ||||
| Alfalfa hay | 40.0 | 40.0 | 40.0 | 40.0 |
| Concentrate | 60.0 | 60.0 | 60.0 | 60.0 |
| Concentrate ingredients (g/100 g fresh matter) | ||||
| Brussel sprouts | - | 8.0 | 16.0 | 24.0 |
| Corn | 32.0 | 28.0 | 25.0 | 23.0 |
| Barley | 30.0 | 28.8 | 26.5 | 23.0 |
| Wheat | 15.0 | 15.0 | 15.0 | 15.0 |
| Soybean meal 46% | 14.0 | 11.2 | 8.5 | 6.0 |
| Wheat bran | 7.0 | 7.0 | 7.0 | 7.0 |
| Calcium soap | 1.0 | 1.0 | 1.0 | 1.0 |
| Calcium carbonate | 0.5 | 0.5 | 0.5 | 0.5 |
| Mineral/vitamin premix | 0.5 | 0.5 | 0.5 | 0.5 |
| Chemical composition of diets 1 | ||||
| Dry matter | 89.7 | 89.7 | 89.7 | 89.7 |
| Organic matter | 93.0 | 92.4 | 91.6 | 90.9 |
| Crude protein | 16.1 | 16.1 | 16.1 | 16.1 |
| Neutral detergent fiber | 31.5 | 31.6 | 31.7 | 31.6 |
| Acid detergent fiber | 15.9 | 16.1 | 16.4 | 16.7 |
| Ether extract | 4.1 | 4.1 | 4.0 | 4.0 |
| Item | Diet | SEM 2 | p | ||||
|---|---|---|---|---|---|---|---|
| Control | BS8 | BS16 | BS24 | Lineal | Quadratic | ||
| Gas production parameters 1 | |||||||
| A (mL/g DM) | 280 a | 289 ab | 290 ab | 293 b | 3.47 | 0.027 | 0.388 |
| c (%/h) | 3.90 | 4.00 | 4.00 | 3.99 | 0.061 | 0.449 | 0.330 |
| Lag (h) | 1.10 | 0.87 | 0.62 | 0.64 | 0.21 | 0.126 | 0.574 |
| AGPR (mL/h) | 7.40 | 7.98 | 8.06 | 8.09 | 0.21 | 0.053 | 0.239 |
| DMED (%) | 40.6 | 41.9 | 42.2 | 41.9 | 0.62 | 0.147 | 0.214 |
| Fermentation parameters (8 h) | |||||||
| Total volatile fatty acids (VFA; µmol) | 1284 a | 1391 ab | 1501 bc | 1561 c | 34.8 | <0.001 | 0.512 |
| Individual VFA (mol/ 100 mol) | |||||||
| Acetate (Ac) | 61.1 | 61.6 | 61.3 | 61.7 | 0.17 | 0.152 | 0.822 |
| Propionate (Pr) | 22.9 a | 22.9 a | 23.3 b | 23.6 b | 0.11 | 0.001 | 0.384 |
| Butyrate | 12.8 c | 12.5 bc | 12.3 b | 11.8 a | 0.12 | <0.001 | 0.582 |
| Minor VFA 3 | 3.11 | 3.01 | 3.04 | 2.95 | 0.061 | 0.141 | 0.929 |
| Ac/Pr (mol/mol) | 2.69 b | 2.71 b | 2.64 ab | 2.63 a | 0.022 | 0.019 | 0.454 |
| NH3-N (mg/L) | 143 a | 148 ab | 154 b | 155 b | 2.9 | 0.033 | 0.560 |
| CH4 (mL) | 6.90 | 7.24 | 7.02 | 7.36 | 0.330 | 0.453 | 0.991 |
| CH4/VFA (mL/mmol) | 5.40 | 5.22 | 4.68 | 4.71 | 0.311 | 0.092 | 0.739 |
| AFOM (%) 4 | 31.3 a | 33.4 ab | 36.3 bc | 37.9 c | 0.84 | <0.001 | 0.474 |
| Fermentation parameters (24 h) | |||||||
| pH | 6.79 b | 6.75 ab | 6.74 a | 6.73 a | 0.01 | 0.004 | 0.394 |
| Total VFA (µmol) | 2446 a | 2650 b | 2673 b | 2684 b | 0.1 | <0.001 | 0.008 |
| Individual VFA (mol/ 100 mol) | |||||||
| Acetate (Ac) | 61.5 | 61.6 | 61.5 | 61.7 | 28.36 | 0.342 | 0.583 |
| Propionate (Pr) | 18.7 a | 19.0 ab | 19.3 b | 19.6 b | 0.12 | <0.001 | 0.788 |
| Butyrate | 15.5 d | 14.9 b | 14.7b | 14.1 a | 0.14 | <0.001 | 0.931 |
| Minor VFA 3 | 4.27 a | 4.53 ab | 4.53 ab | 4.57 b | 0.092 | 0.048 | 0.217 |
| Ac/Pr (mol/mol) | 3.31 c | 3.28 bc | 3.20 ab | 3.16 a | 0.0143 | 0.003 | 0.962 |
| NH3-N (mg/L) | 189 | 203 | 207 | 209 | 6.1 | 0.090 | 0.506 |
| CH4 (mL) | 14.9 | 15.1 | 14.8 | 15.2 | 0.41 | 0.804 | 0.846 |
| CH4/VFA (mL/mmol) | 6.10 b | 5.69 ab | 5.53 a | 5.65 a | 0.140 | 0.039 | 0.095 |
| AFOM (%) 4 | 60.5 a | 64.0 b | 65.1 b | 65.4 b | 0.66 | 0.001 | 0.007 |
| Item 1 | Diet | SEM 2 | p | ||||
|---|---|---|---|---|---|---|---|
| Control | BS8 | BS16 | BS24 | Lineal | Quadratic | ||
| Dry matter | |||||||
| a (%) | 33.6 a | 34.7 a | 37.9 b | 40.5 c | 0.33 | <0.001 | 0.269 |
| b (%) | 45.6 b | 44.6 b | 42.5 a | 40.9 a | 0.34 | 0.001 | 0.628 |
| a + b (%) | 79.2 | 79.4 | 80.4 | 81.4 | 0.59 | 0.141 | 0.705 |
| c (h−1) | 0.262 | 0.240 | 0.206 | 0.238 | 0.0102 | 0.227 | 0.180 |
| ED (%) | 72.7 a | 72.7 a | 73.2 ab | 75.1 b | 0.67 | 0.042 | 0.192 |
| Crude protein | |||||||
| a (%) | 35.7 a | 47.2 c | 45.8 b | 49.1 d | 0.23 | <0.001 | <0.001 |
| b (%) | 55.9 c | 44.5 b | 45.7 b | 42.2 a | 0.34 | <0.001 | 0.001 |
| a + b (%) | 91.6 | 91.7 | 91.5 | 91.3 | 0.23 | 0.551 | 0.728 |
| c (h−1) | 0.167 a | 0.176 ab | 0.227 bc | 0.242 c | 0.0091 | 0.008 | 0.871 |
| ED (%) | 80.2 a | 83.1 b | 84.3 b | 85.0 b | 0.77 | 0.004 | 0.211 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Evan, T.; Vintimilla, A.; Marcos, C.N.; Ranilla, M.J.; Carro, M.D. Evaluation of Brassica Vegetables as Potential Feed for Ruminants. Animals 2019, 9, 588. https://doi.org/10.3390/ani9090588
de Evan T, Vintimilla A, Marcos CN, Ranilla MJ, Carro MD. Evaluation of Brassica Vegetables as Potential Feed for Ruminants. Animals. 2019; 9(9):588. https://doi.org/10.3390/ani9090588
Chicago/Turabian Stylede Evan, Trinidad, Andrea Vintimilla, Carlos N. Marcos, María José Ranilla, and María Dolores Carro. 2019. "Evaluation of Brassica Vegetables as Potential Feed for Ruminants" Animals 9, no. 9: 588. https://doi.org/10.3390/ani9090588
APA Stylede Evan, T., Vintimilla, A., Marcos, C. N., Ranilla, M. J., & Carro, M. D. (2019). Evaluation of Brassica Vegetables as Potential Feed for Ruminants. Animals, 9(9), 588. https://doi.org/10.3390/ani9090588