
Amelioration of Cadmium-Induced Oxidative Damage in Wistar Rats by Vitamin C, Zinc and N-Acetylcysteine

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Chemicals
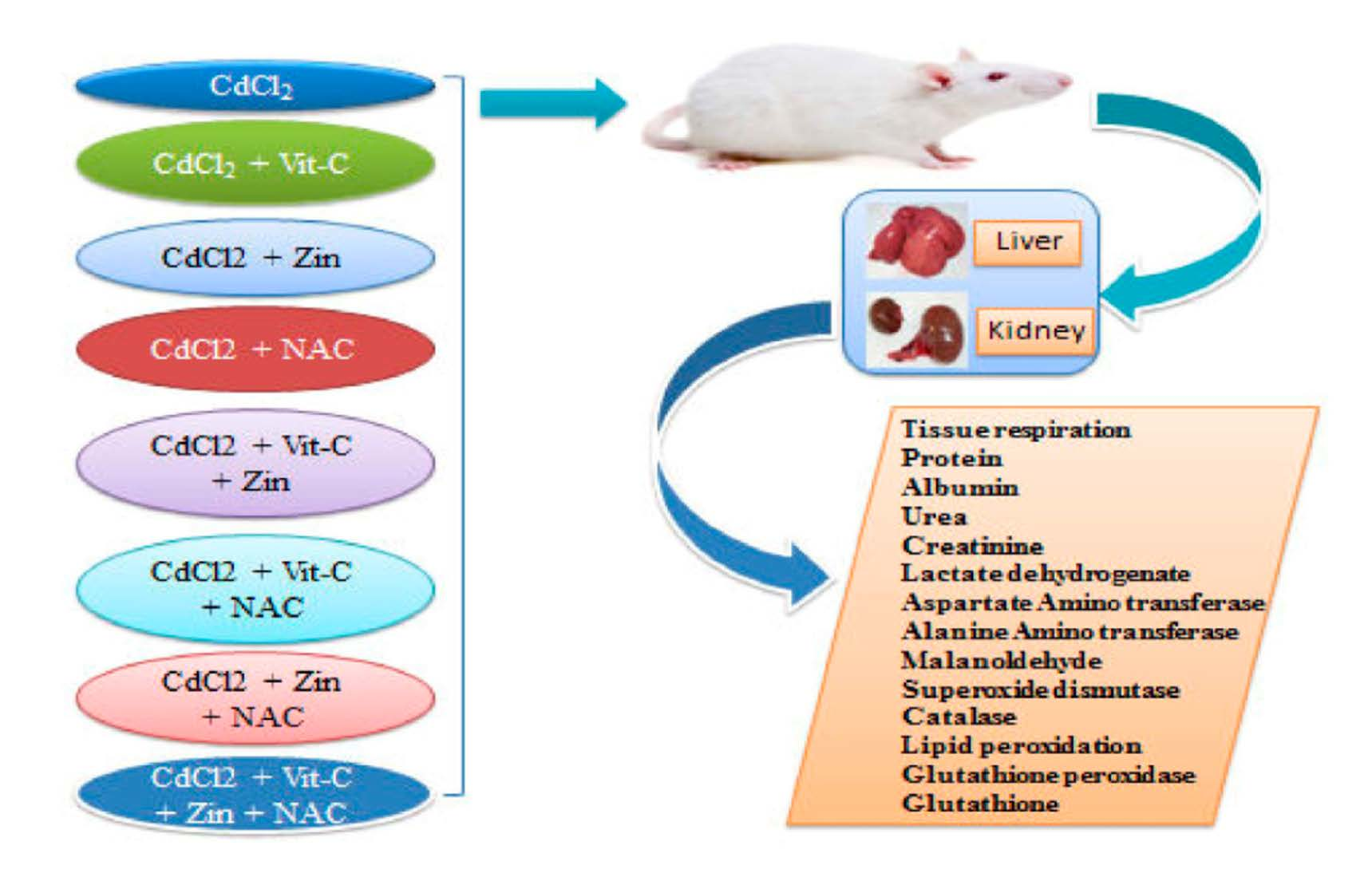
2.3. Experimental Design
2.4. Biochemical Analyses Were Performed by the following Methods
2.4.1. Tissue Respiration
2.4.2. Total Protein
2.4.3. Urea
2.4.4. Creatinine
2.4.5. Lactate Dehydrogenase (LDH)
2.4.6. Aspartate Amino Transferase (AST)
2.4.7. Alanine Amino Transferase (ALT)
2.4.8. Malondialdehyde (MDA)
2.4.9. Superoxide Dismutase (SOD)
2.4.10. Catalase (CAT)
2.4.11. Lipid Peroxidation (LPO)
2.4.12. Glutathione Peroxidase (GPx)
2.4.13. Glutathione (GSH)
2.5. Statistical Analysis and Data Presentation
3. Results
3.1. Body Weights and Relative Tissue Weights
3.2. Respiratory and Antioxidant Status
4. Discussion
4.1. Body Weights and Relative Organ Weights
4.2. Respiratory and Antioxidant Status
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jessica, B.; Emmanuel, S.; Renald, B. Heavy metal pollution in the environment and their toxicological effects on humans. Heliyon 2020, 6, e04691. [Google Scholar]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy metals toxicity in the environment. Mol. Clin. Environ. Toxicol. 2012, 101, 133–164. [Google Scholar]
- Das, S.C.; Al-Naemi, H.A. Cadmium toxicity: Oxidative stress, inflammation and tissue injury. Occup. Dis. Environ. Med. 2019, 7, 144–163. [Google Scholar] [CrossRef] [Green Version]
- Mohammad, N.S.; Tangpong, J.; Rahman, M.M. Toxicodynamics of Lead, Cadmium, Mercury and Arsenic induced kidney toxicity and treatment strategy: A mini review. Toxicol. Rep. 2018, 5, 704–713. [Google Scholar]
- Bull, S. Cadmium, Toxicological overview. Health Prot. Agency 2010, 3, 3–15. [Google Scholar]
- IPCS. International Programme on Chemical Safety. Cadmium. In Poisons Information Monograph, Dimercaprol (PIM 193). 2010. Available online: https://www.who.int/ (accessed on 14 December 2021).
- Genchi, G.; Maria, S.S.; Lauria, G.; Carocci, A.; Catalano, A. The Effects of Cadmium Toxicity. Int. J. Environ. Res. Public Health 2020, 17, 3782. [Google Scholar] [CrossRef] [PubMed]
- Jacopo, J.V.B.; Fiorillo, C.; Carrino, D.; Paternostro, F.; Taddei, N.; Gulisano, M.; Pacini, A.; Becatti, M. Cadmium-induced stress: Focus on the central nervous system. Antioxidants 2020, 9, 492–513. [Google Scholar]
- Milena, A.; Aleksandra, B.D.; Evica, A.; Biljana, A.; Stanic, M.; Jelena, K.S.; Vesna, S.K.; Milos, J.; Novika, M.; Jelina, K.S.; et al. Toxic effect of Acute Cadmium and Lead exposure in Rat blood, liver and kidney. Int. J. Environ. Res. Public Health 2019, 16, 274–295. [Google Scholar]
- Brzoska, M.M.; Borowska, S.; Tomezyk, M. Antioxidants as a potential preventive and therapeutic strategy for Cadmium. Curr. Drug Targets 2016, 17, 1350–1384. [Google Scholar] [CrossRef]
- Amel, S.M.; Loukidi, B.; Reda, B. Effects of vitamin C and E against oxidative stress: Is Antioxidant supplementation Efficient? Curr. Neutraceuticals 2020, 1, 33–41. [Google Scholar]
- Elias, A.; Deo, O. Hepatoprotective effect of Vitamin C (Ascorbic acid). Pharmacol. Pharm. 2013, 4, 84–92. [Google Scholar]
- Umbreit, W.W.; Burris, R.H.; Stauffer, J.F. Manometric Techniques, 3rd ed.; Burgess Publishing Co.: Minneapolis, MN, USA, 1964. [Google Scholar]
- Lowry, O.H.; Rosenbrough, Ν.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Patton, C.J.; Crouch, S.R. Spectrophotometric and kinetics investigation of the Berthelot reaction for the determination of ammonia. Anal. Chem. 1977, 49, 464–469. [Google Scholar] [CrossRef]
- Faulkner, W.R.; King, J.W. Renal function. In Fundamentals of Clinical Chemistry; Tietz, N., Ed.; WB. Samders Co.: Philadelphia, PA, USA, 1976; pp. 975–1014. [Google Scholar]
- Kornberg, A. Methods of Enzymatic Analysis; Bergmeyer, H.O., Ed.; Academic Press: New York, NY, USA, 1974. [Google Scholar]
- Reitman, S.; Frankel, S. A colorimetric method for determination of serum ALT and AST. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autooxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Wendel, A. Glutathione Peroxidase. Methods Enzymol. 1981, 77, 325–333. [Google Scholar] [PubMed]
- Moron, M.S.; Depierre, J.W.; Mannervik, B. Levels of glutathione; glutathione reductase and glutathione S-transferase activities in rat lung and liver. Biochem. Biophys. Acta 1979, 582, 67–78. [Google Scholar] [CrossRef]
- Rahman, Z.; Singh, V.P. The relative impact of toxic heavy metals (THMs) [arsenic (As), cadmium (Cd), chromium (Cr) (VI), mercury (Hg), and lead (Pb)] on the total environment: An overview. Environ. Monit. Assess. 2019, 191, 419. [Google Scholar] [CrossRef]
- Renuka, M.; Aparna, Y.; Venkataramanaiah, P.; Srinivasulu Reddy, M. Vitamin, C, E and Zinc ameliorates Cadmium toxicity induced biochemical changes in male albino rats. Toxicol. Forensic Med. 2021, 6, 13–19. [Google Scholar]
- Casalino, E.; Sblano, C.; Landriscina, C. Enzyme activity alteration by Cadmium administration to rat: The possibility of iron involvement in lipid peroxidation. Arch. Biochem. Biophys. 1997, 346, 171–179. [Google Scholar] [CrossRef]
- Bauer, R.; Demeter, I.; Hasemann, V.; Jahansen, J.T. Structural properties of the Zinc site in the cup.; n-superoxide dismutase: Perturbed angular correlation of gamma ray spectroscopy on the Cu-111 Cd-Superoxide Dismutage derivative. Biochem. Biophys. Res. Commum. 1980, 94, 1296–1302. [Google Scholar] [CrossRef]
- Hari, K.; Aydm, C.; Vahit, K.; Alpaslan, D.; Kadir, S. Effects of selenium with Vitamin E and melatonin on cadmium-induced oxidative damage in rat liver and kidneys. Biol. Trace. Elem. Res. 2008, 125, 236–244. [Google Scholar]
- Patra, R.C.; Swarup, D.; Senapati, S.K. Effects of cadmium on lipid peroxides and superoxide dismutase in hepatic, renal and testicular tissue of rats. Vet. Hum. Toxicol. 1999, 41, 65–67. [Google Scholar]
- Shukla, G.S.; Chandra, S.V. Cadmium toxicity and bioantioxidants: Status of Vitamin E and ascorbic acid of selected organs in rat. J. Appl. Toxicol. 1989, 9, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, Z.A.; Vu, T.T.; Zaman, K. Oxidative stress as a mechanism of chronic cadmium induced hepatotoxicity and renal toxicity and protection by antioxidants. Toxicol. Appl. Pharmacol. 1999, 154, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Kostic, M.M.; Ognjanovic, B.; Dimitrijevic, S.; Zikic, R.V.; Stajn, A.; Rosic, G.L.; Ivkovic, R.V. Cadmium-induced changes of antioxidant and metabolic status in red blood cells of rats: In vivo effects. Eur. J. Haematol. 1993, 51, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Zorica, P.B.; Danijela, D.C.; Zivorad, M. Zinc or Magnesium supplementation modulates cadmium intoxication in blood, kidney, spleen and bone of rabbits. Biol. Trace. Elem. Res. 2008, 124, 110–117. [Google Scholar]
- Mates, J.M. Effects of antioxidant enzymes in the molecular control of reactive oxygen species toxicology. Toxicology 2000, 153, 83–104. [Google Scholar] [CrossRef]
- Giuseppe, C.; Elisa, F.; Claudio, L.; Anna, M.R. Cadmium and mitochondria. Mitochondrion 2009, 9, 377–384. [Google Scholar]
- Koyu, A.; Gokcimen, A.; Ozguner, F.; Bayram, D.S.; Kocak, A. Evaluation of the effects of cadmium on rat liver. Mol. Cell. Biochem. 2006, 284, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Şlencu, B.G.; Ciobanu, C.; Solcan, C.; Anton, A.; Ciobanu, S.; Solcan, G.; Cuciureanu, R. Effect of Selenium Supplementation on Serum Amylase, Lactate Dehydrogenase and Alkaline Phosphatase Activities in Rats Exposed to Cadmium or Lead. Cercet. Agron. Mold. 2014, 4, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Babaknejad, N.; Moshtaghie, A.A.; Nayeri, H.; Hani, M.; Bahrami, S. Protective Role of Zinc and Magnesium against Cadmium Nephrotoxicity in Male Wistar Rats. Biol. Trace Element Res. 2016, 174, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.C.; Frei, B. Does vitamin C act as a pro-oxidant under physiological conditions? The Federation of American Societies for Experimental Biology. FASEB J. 1999, 13, 1007–1024. [Google Scholar] [CrossRef] [Green Version]
- Tarasub, N.; Junseecha, T.; Tarasub, C.; Ayutthaya, W.D.N. Protective effects of Curcumin, Vitamin, C, or their Combination on Cadmium-induced hepatotoxicity. J. Basic Clin. Pharm. 2012, 3, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Yousef, M.I.; Salem, M.H.; Kamel, K.I.; Hassan, G.A.; El-Nouty, F.D. Influence of ascorbic acid supplementation on the haematological and clinical biochemistry parameters of male rabbits exposed to aflatoxin B1. J. Environ. Sci. Health 2003, 38, 193–209. [Google Scholar] [CrossRef]
- Yousef, M.I. Aluminium-induced changes in hemato-biochemical parameters, lipid peroxidation and enzyme activities of male rabbits: Protective role of ascorbic acid. Toxicology 2004, 199, 47–57. [Google Scholar] [CrossRef]
- Gunther, M.R. Probing the free radicals formed in the metmyoglobin–hydrogen peroxide reaction. Free Radic. Biol. Med. 2004, 36, 1345–1354. [Google Scholar] [CrossRef]
- Pradeepkiran, J.A.; Siva Prasad, B.; Rajesh, A.V.; Bhaskar, M. Calcium, Zinc and Vitamin E ameliorate Cadmium-induced renal oxidative damage in albino rats. Toxicol. Rep. 2016, 3, 591–597. [Google Scholar]
- Jan, A.T.; Azam, M.; Siddiqui, K.; Ali, A.; Choi, I.; Haq, Q.M.R. Heavy metals and human health: Mechanistic insight into toxicity and counter defence system of Antioxidants. Int. J. Mol. Sci. 2015, 16, 29592–29630. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
| Parameters | Control | Cd Treated | Vit-C | Zinc | NAC | C + Zinc | C + NAC | Zinc + NAC | C + Zinc +NAC |
|---|---|---|---|---|---|---|---|---|---|
| IBW | 225.19 ±10.12 | 226.12 ±10.14 | 225.18 ±9.79 | 224.19 ±10.17 | 225.49 ±10.15 | 225.42 ±10.19 | 226.74 ±10.45 | 225.48 ±10.12 | 224.14 ±10.15 |
| FBW | 268.15 a ±10.17 | 122.35 b ±10.22 | 238.14 b,c ±10.12 | 237.19 b,c ±10.03 | 238.32 b,c ±10.05 | 238.05 b,c ±10.14 | 237.78 b,c ±9.89 | 238.04 b,c ±9.88 | 238.05 b,c ±10.13 |
| BWG/L | +19.07 b | −45.89 a | +5.76 b | +5.79 b | +5.69 b | +5.60 b | +4.87 b | +5.57 b | +5.60 b |
| LW | 10.84 a ±0.63 PDC | 4.82 b,c ±0.25 −55.54 | 8.12 b,c ±0.45 −25.09 | 8.24 b,c ±0.38 −23.99 | 8.25 b,c ±0.39 −23.89 | 8.18 b,c ±0.42 −32.52 | 8.25 b,c ±0.41 −23.89 | 8.05 b,c ±0.38 −27.74 | 8.19 b,c ±0.35 −24.45 |
| HIS-L | 4.04 a PDC | 3.94 a −2.48 | 3.41 b −15.59 | 3.48 b −13.86 | 3.46 b −14.36 | 3.44 b −14.85 | 3.47 b −14.11 | 3.38 b −16.34 | 3.44 b −14.85 |
| KW | 2.93 a ±0.28 PDC | 1.23 b ±0.14 −44.37 | 2.08 b,c ±0.15 −29.01 | 2.12 b,c ±0.17 −27.65 | 2.11 b,c ±0.14 −27.99 | 2.14 b,c ±0.17 −29.96 | 2.14 b,c ±0.18 −26.96 | 2.12 b,c ±0.15 −27.65 | 2.11 b,c ±0.15 −27.99 |
| HIS-K | 1.09 a PDC | 1.01 a −7.34 | 0.87 b −20.18 | 0.89 b −18.35 | 0.89 b −18.35 | 0.90 b −17.43 | 0.90 b −17.43 | 0.89 b −18.35 | 0.89 b −18.35 |
| Parameters | Control | Cd Treated | Vit-C | Zinc | NAC | C + Zinc | C + NAC | Zinc + NAC | C + Zinc +NAC |
|---|---|---|---|---|---|---|---|---|---|
| Basal Metabolic Rate (BMR) | 4.23 a ±0.24 PDC | 2.72 b ±0.13 −35.70 | 3.41 b,c ±0.13 −19.39 | 3.54 b,c ±0.15 −16.31 | 3.53 b,c ±0.16 −16.55 | 3.54 b,c ±0.13 −16.31 | 3.48 b,c ±0.15 −17.73 | 3.53 b,c ±0.14 −16.55 | 3.54 b,c ±0.15 −16.31 |
| Tissue Respiration (TR), Liver | 1033.46 a ±24.77 PDC | 642.14 b ±18.15 −37.87 | 842.18 b,c ±16.19 −18.51 | 852.14 b,c ±17.15 −17.55 | 860.15 b,c ±17.19 −16.77 | 862.19 b,c ±17.14 −16.57 | 863.14 b,c ±16.75 −16.48 | 861.15 b,c ±17.13 −16.67 | 867.18 b,c ±14.19 −16.09 |
| Tissue Respiration (TR), Kidney | 858.77 a ±12.85 PDC | 508.42 b ±12.12 −40.80 | 660.18 b,c ±12.19 −23.12 | 672.13 b,c ±15.14 −21.73 | 673.13 b,c ±11.14 −21.61 | 671.72 b,c ±13.49 −21.78 | 675.70 b,c ±14.19 −21.32 | 673.49 b,c ±13.45 −21.58 | 677.43 b,c ±14.46 −21.12 |
| Parameters | Control | Cd Treated | Vit-C | Zinc | NAC | C + Zinc | C + NAC | Zinc + NAC | C + Zinc + NAC |
|---|---|---|---|---|---|---|---|---|---|
| Total Protein (mg/mL) | 15.38 a ±1.29 PDC | 33.49 b ±2.28 +117.75 | 20.31 b,c ±1.72 +32.05 | 25.34 b,c ±1.83 +64.48 | 26.77 b,c ±1.49 +74.06 | 26.82 b,c ±1.34 +74.97 | 22.14 b,c ±1.13 +43.95 | 20.74 b,c ±1.14 +34.85 | 20.18 b,c ±1.12 +31.21 |
| Albumin (mg/mL) | 2.38 a ±0.22 PDC | 2.89 b ±0.23 +21.43 | 3.04 b,c ±0.23 +27.73 | 3.08 b,c ±0.24 +29.41 | 3.12 b,c ±0.25 +31.09 | 2.94 b,c ±0.22 +23.53 | 2.95 b,c ±0.22 +23.95 | 2.99 b,c ±0.24 +25.63 | 3.02 b,c ±0.22 +26.89 |
| Urea (mg/dL) | 34.14 a ±2.12 | 41.19 b ±2.74 | 38.77 b,c ±2.49 | 39.42 b,c ±2.49 | 38.45 b,c ±2.44 | 39.25 b,c ±2.38 | 37.77 b,c ±2.74 | 38.41 ±2.49 | 36.77 b,c ±1.42 |
| Creatinine (mg/dL) | 0.65 a ±0.04 PDC | 0.83 b ±0.08 + 20.62 | 0.82 b,c ±0.07 +13.56 | 0.79 b,c ±0.08 +21.54 | 0.78 b,c ±0.07 +20.00 | 0.77 b,c ±0.07 +18.46 | 0.74 b,c ±0.08 +13.85 | 0.73 b,c ±0.08 +12.31 | 0.74 b,c ±0.07 +13.85 |
| LDH (µ moles of formazon formed/mg protein/h) | 18.14 a ±0.25 PDC | 39.79 b ±0.34 +119.35 | 35.14 b,c ±0.29 +93.72 | 34.72 b,c ±0.27 +91.40 | 27.45 b,c ±0.25 +51.32 | 25.79 b,c ±0.34 +42.18 | 24.74 b,c ±0.31 +36.38 | 21.74 b,c ±0.31 +19.85 | 25.78 b,c ±0.29 +42.12 |
| AST (U/L) | 53.14 a ±2.77 PDC | 459.78 b ±25.85 +765 | 254.72 b,c ±15.75 +379 | 277.45 b,c ±18.74 +422 | 245.42 b,c ±19.45 +361 | 205.74 b,c ±18.42 +287 | 214.12 b,c ±20.14 +303 | 205.74 b,c ±18.85 +287 | 201.71 b,c ±18.45 +279 |
| ALT (U/L) | 39.77 a ±2.04 PDC | 365.72 b ±20.14 +196 | 204.14 b,c ±15.05 +413 | 201.45 b,c ±19.41 +407 | 194.15 b,c ±19.42 +388 | 190.45 b,c ±20.42 +379 | 185.45 b,c ±20.14 +366 | 168.74 b,c ±14.55 +324 | 158.74 b,c ±12.85 +299 |
| ALP (U/L) | 108.12 a ±8.14 PDC | 38.19 b ±2.15 −64.68 | 62.13 b,c ±3.75 −42.54 | 68.34 b,c ±3.49 −36.79 | 70.13 b,c ±3.74 −35.14 | 73.14 b,c ±4.18 −32.35 | 75.18 b,c ±4.42 −30.47 | 78.11 b,c ±4.49 −27.76 | 80.13 b,c ±4.13 −25.87 |
| Parameters | Control | Cd Treated | Vit-C | Zinc | NAC | C + Zinc | C + NAC | Zinc + NAC | C + Zinc + NAC |
|---|---|---|---|---|---|---|---|---|---|
| MDA | 475 a ±25 PDC | 1015 b ±39 +114 | 885 b,c ±29 +86 | 901 b,c ±40 +90 | 950 b,c ±32 +100 | 809 b,c ±29 +70 | 804 b,c ±32 +69 | 775 b,c ±29 +64 | 670 b,c ±32 +41 |
| SOD | 66.79 a ±2.15 PDC | 31.73 b ±1.14 −35 | 34.44 b,c ±1.18 −48 | 36.18 b,c ±1.15 −46 | 38.14 b,c ±1.19 −43 | 40.12 b,c ±1.28 −40 | 42.14 b,c ±1.29 −37 | 44.75 b,c ±1.32 −33 | 51.14 b,c ±1.12 −23 |
| CAT | 88.14 a ±2.12 PDC | 42.44 b ±1.13 −52 | 44.75 b,c ±1.14 −49 | 47.75 b,c ±1.25 −46 | 49.75 b,c ±1.32 −43 | 53.72 b,c ±1.34 −40 | 59.14 b,c ±1.42 −37 | 60.19 b,c ±1.38 −33 | 68.13 b,c ±1.18 −23 |
| LPO | 5.14 a ±0.28 PDC | 9.85 b ±0.38 +92 | 8.35 b,c ±0.32 +63 | 8.19 b,c ±0.31 +59 | 7.74 b,c ±0.26 +51 | 7.15 b,c ±0.25 +39 | 6.74 b,c ±0.27 +31 | 6.34 b,c ±0.26 +23 | 6.15 b,c ±0.24 +20 |
| GPX | 8.15 a ±0.46 PDC | 4.05 b ±0.28 −50 | 4.18 b,c ±0.24 −49 | 4.34 b,c ±0.25 −47 | 4.75 b,c ±0.26 −42 | 5.25 b,c ±0.29 −36 | 5.79 b,c ±0.32 −29 | 6.42 b,c ±0.35 −22 | 7.04 b,c ±0.28 −14 |
| GSH | 4.93 a ±0.41 PDC | 2.85 b ±0.22 −42 | 2.92 b,c ±0.23 −41 | 2.94 b,c ±0.24 −40 | 3.02 b,c ±0.27 −39 | 3.14 b,c ±0.29 −36 | 3.25 b,c ±0.32 −34 | 3.32 b,c ±0.33 −33 | 3.49 b,c ±0.32 −29 |
| Parameters | Control | Cd Treated | Vit-C | Zinc | NAC | C + Zinc | C + NAC | Zinc + NAC | C + Zinc + NAC |
|---|---|---|---|---|---|---|---|---|---|
| MDA | 528 a ±29 PDC | 1114 b ±35 +111 | 1074 b,c ±38 +103 | 995 b,c ±24 +88 | 880 b,c ±22 +67 | 740 b,c ±25 +40 | 730 b,c ±26 +38 | 722 b,c ±27 +37 | 685 b,c ±24 +30 |
| SOD | 45.72 a ±2.14 PDC | 24.12 b ±1.10 −47 | 26.19 b ±1.12 −43 | 28.12 b ±1.14 −38 | 30.12 b,c ±1.15 −34 | 31.18 b,c ±1.14 −32 | 32.18 b,c ±1.14 −30 | 33.11 b,c ±1.14 −28 | 33.75 b,c ±1.71 −26 |
| CAT | 82.18 a ±2.12 PDC | 43.42 b ±1.14 −47 | 45.45 b ±1.13 −45 | 48.74 b ±1.14 −41 | 51.23 b ±1.24 −38 | 53.42 b,c ±1.42 −35 | 58.41 b,c ±1.56 −29 | 60.79 b,c ±1.57 −26 | 65.77 b,c ±2.12 −20 |
| LPO | 4.73 a ±0.24 PDC | 8.73 b ±0.42 +85 | 8.14 b,c ±0.33 +72 | 8.04 b,c ±0.32 +70 | 7.34 b,c ±0.29 +55 | 7.49 b,c ±0.28 +58 | 6.43 b,c ±0.31 +36 | 6.05 b,c ±0.25 +28 | 5.79 b,c ±0.23 +22 |
| GPX | 7.32 a ±0.38 PDC | 3.92 b ±0.24 −46 | 4.14 b ±0.24 −43 | 4.54 b ±0.25 −38 | 4.89 b,c ±0.26 −33 | 5.12 b,c ±0.26 −30 | 5.34 b,c ±0.25 −27 | 5.59 b,c ±0.24 −24 | 6.13 b,c ±0.25 −16 |
| GSH | 4.12 a ±0.34 PDC | 2.42 b ±0.22 −41 | 2.63 b,c ±0.22 −36 | 2.79 b,c ±0.21 −32 | 2.84 b,c ±0.23 −31 | 3.02 b,c ±0.23 −27 | 3.12 b,c ±0.24 −24 | 3.23 b,c ±0.24 −22 | 3.38 b,c ±0.24 −18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poli, V.; Madduru, R.; Aparna, Y.; Kandukuri, V.; Motireddy, S.R. Amelioration of Cadmium-Induced Oxidative Damage in Wistar Rats by Vitamin C, Zinc and N-Acetylcysteine. Med. Sci. 2022, 10, 7. https://doi.org/10.3390/medsci10010007
Poli V, Madduru R, Aparna Y, Kandukuri V, Motireddy SR. Amelioration of Cadmium-Induced Oxidative Damage in Wistar Rats by Vitamin C, Zinc and N-Acetylcysteine. Medical Sciences. 2022; 10(1):7. https://doi.org/10.3390/medsci10010007
Chicago/Turabian StylePoli, Venkataramanaiah, Renuka Madduru, Yenukolu Aparna, Vimala Kandukuri, and Srinivasulu Reddy Motireddy. 2022. "Amelioration of Cadmium-Induced Oxidative Damage in Wistar Rats by Vitamin C, Zinc and N-Acetylcysteine" Medical Sciences 10, no. 1: 7. https://doi.org/10.3390/medsci10010007
APA StylePoli, V., Madduru, R., Aparna, Y., Kandukuri, V., & Motireddy, S. R. (2022). Amelioration of Cadmium-Induced Oxidative Damage in Wistar Rats by Vitamin C, Zinc and N-Acetylcysteine. Medical Sciences, 10(1), 7. https://doi.org/10.3390/medsci10010007